
Single-Cell Proteomic Profiling Identifies Nanoparticle Enhanced Therapy for Triple Negative Breast Cancer Stem Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemical
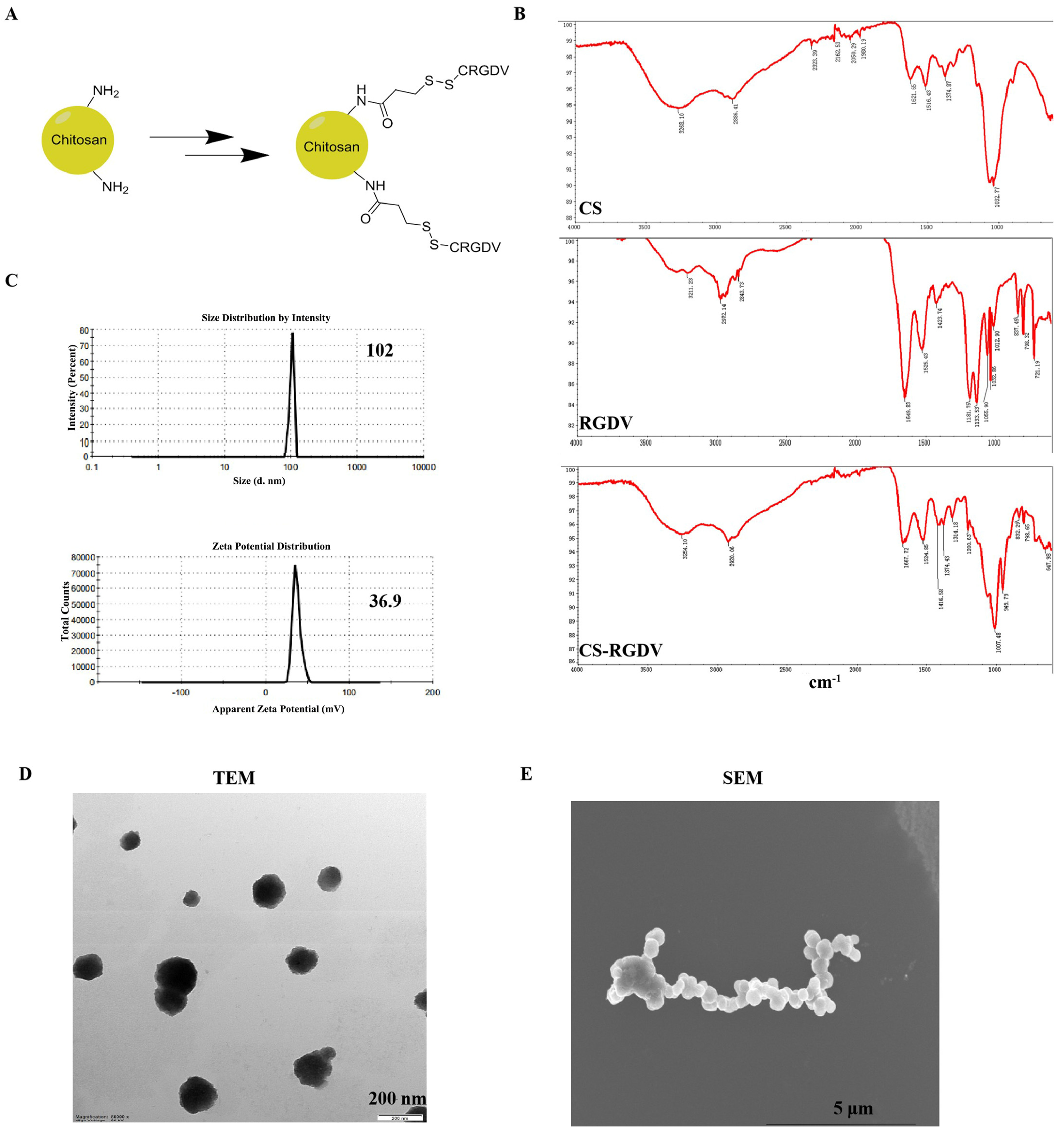
2.2. Preparation and Characterization of Chitosan-Modified Peptide Nanoparticles
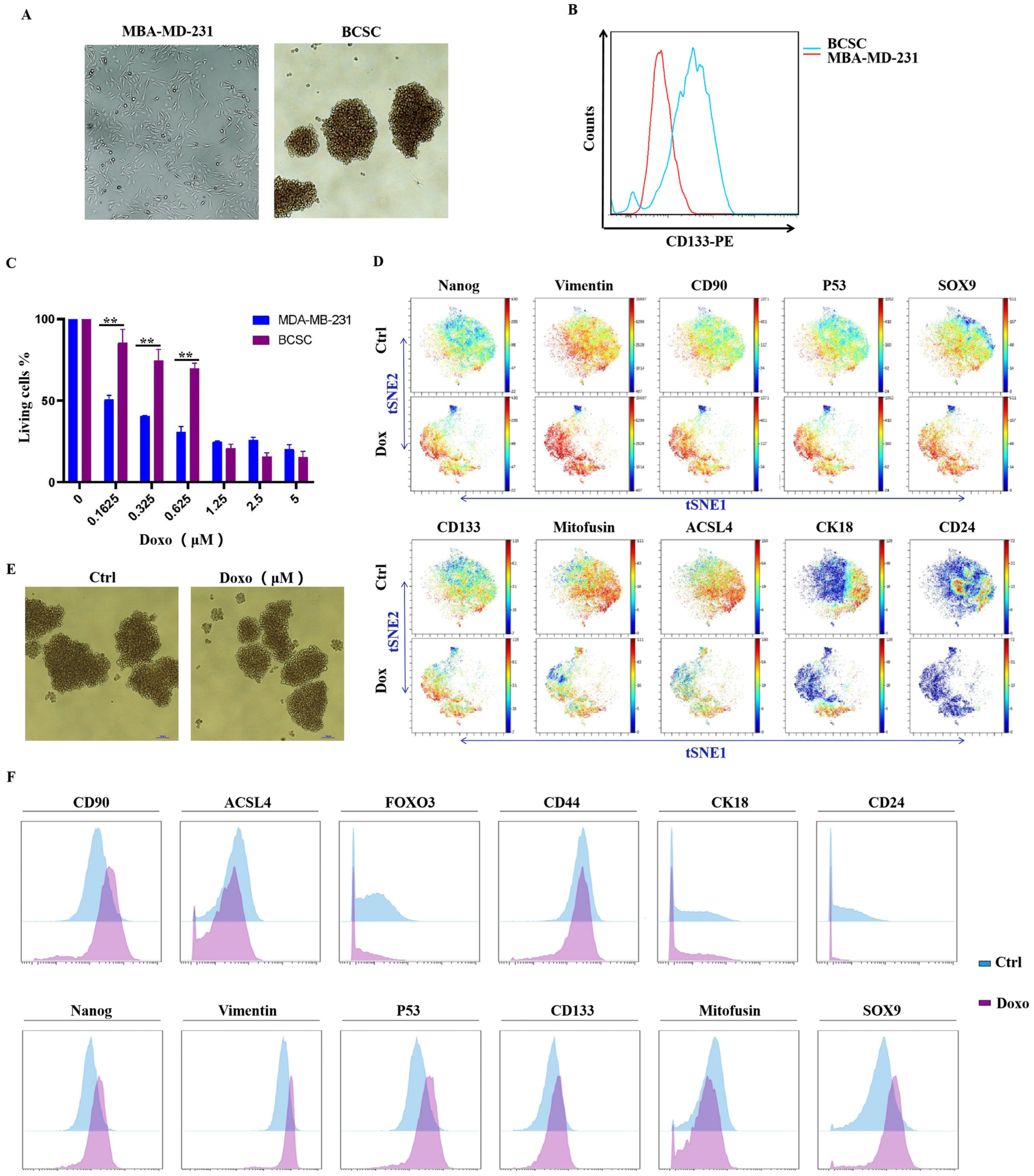
2.3. MDA-MB-231/4T1 Cells and Spheroid Cells Culture
2.4. CCK8 Assay Detected Drug Sensitivity
2.5. Immunofluorescence Staining
2.6. Time-of-Flight Mass Cytometry
2.7. Statistical Analysis
3. Results
3.1. Prepared and Characterization of Chitosan Nanosized Peptide
3.2. MDA-MB-231-Cultured Spheroid Cells Exhibit BCSC-like Properties, and Administration of Doxorubicin Could Enhance Stemness in Spheroid Cells
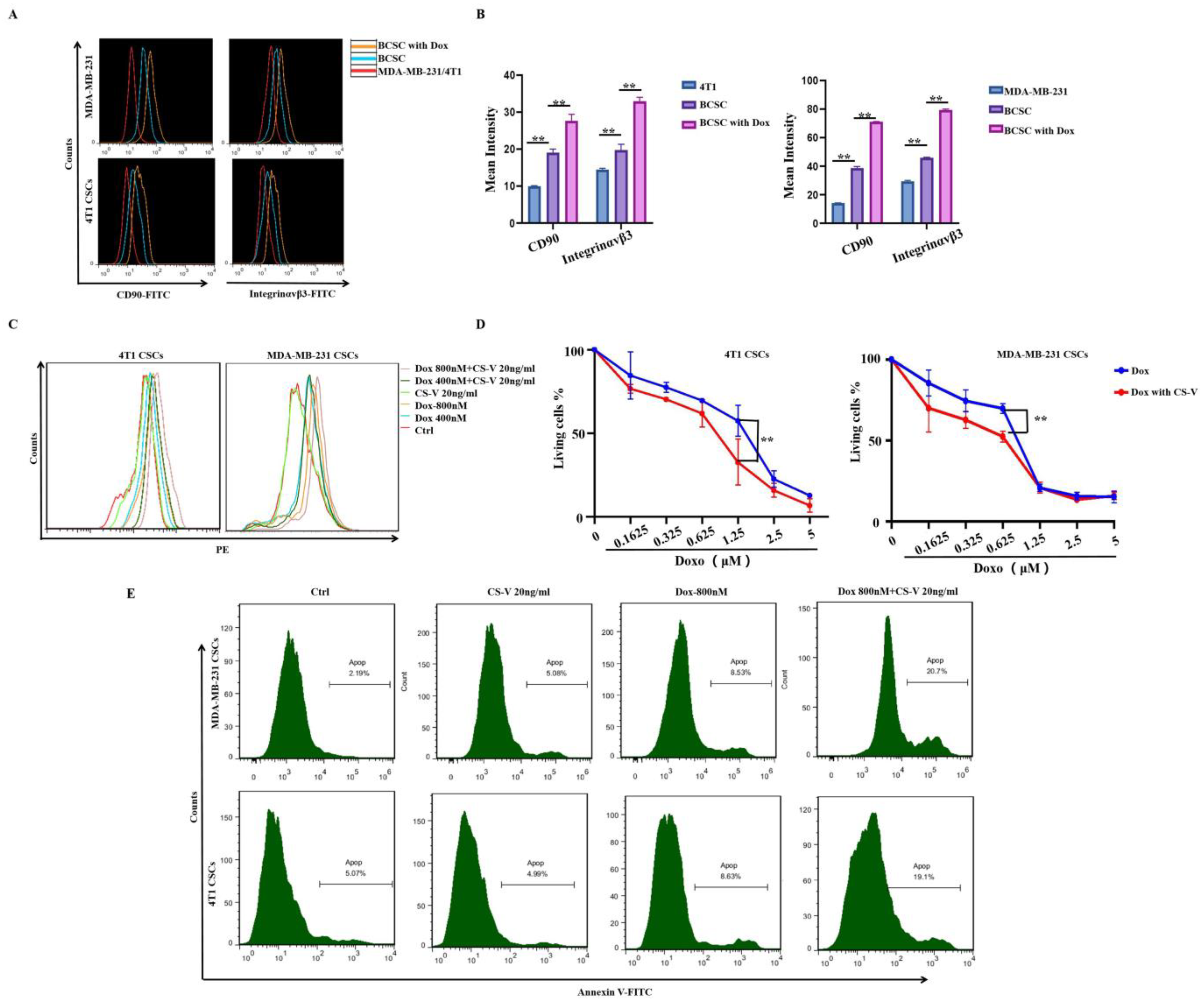
3.3. Integrin αvβ3 and CD90 Expression Was Significantly Induced by Doxorubicin, and Integrin αvβ3-Targeted CS-V Nanopeptide Could Promote Doxorubicin Efficacy against MDA-MB-231 Spheroid Cells
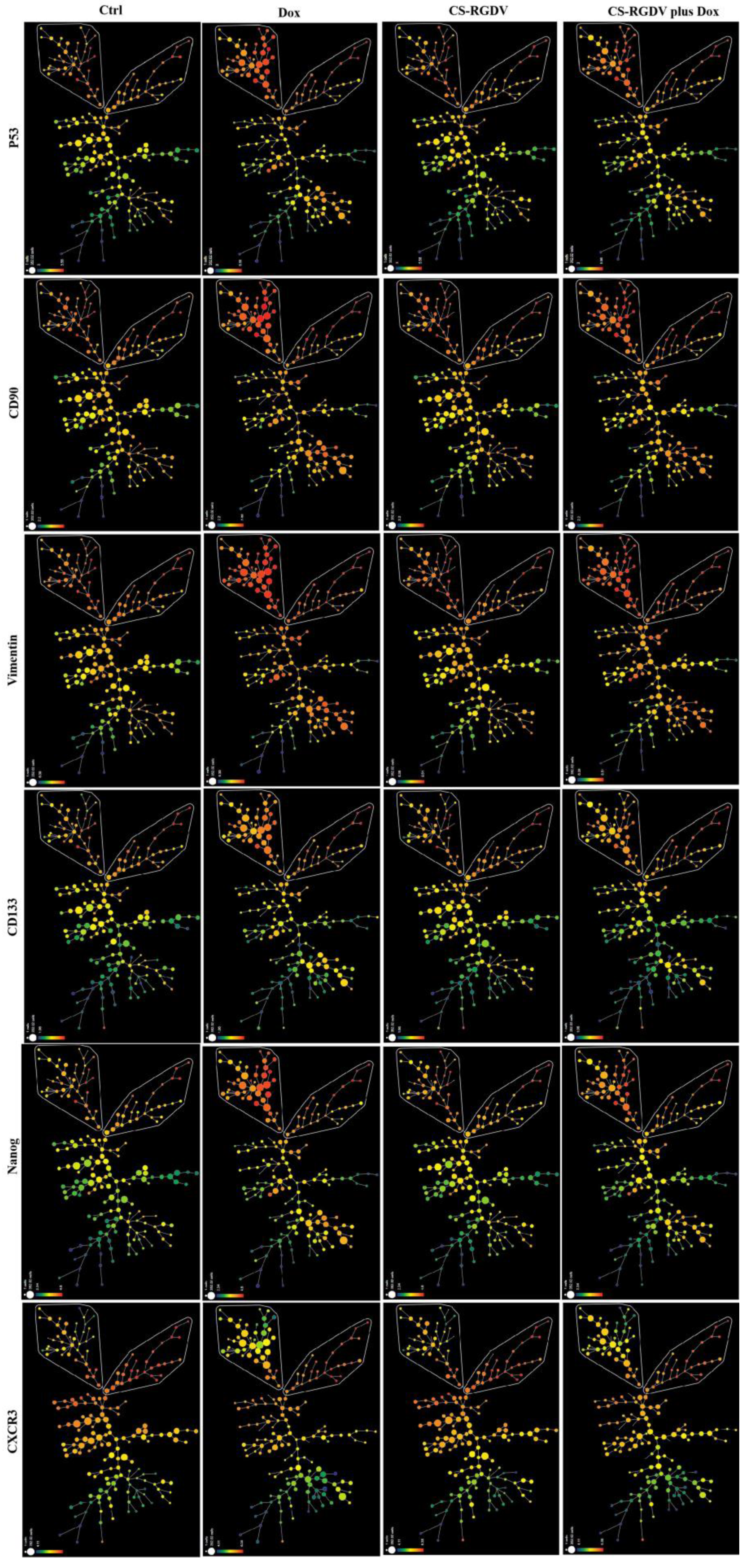
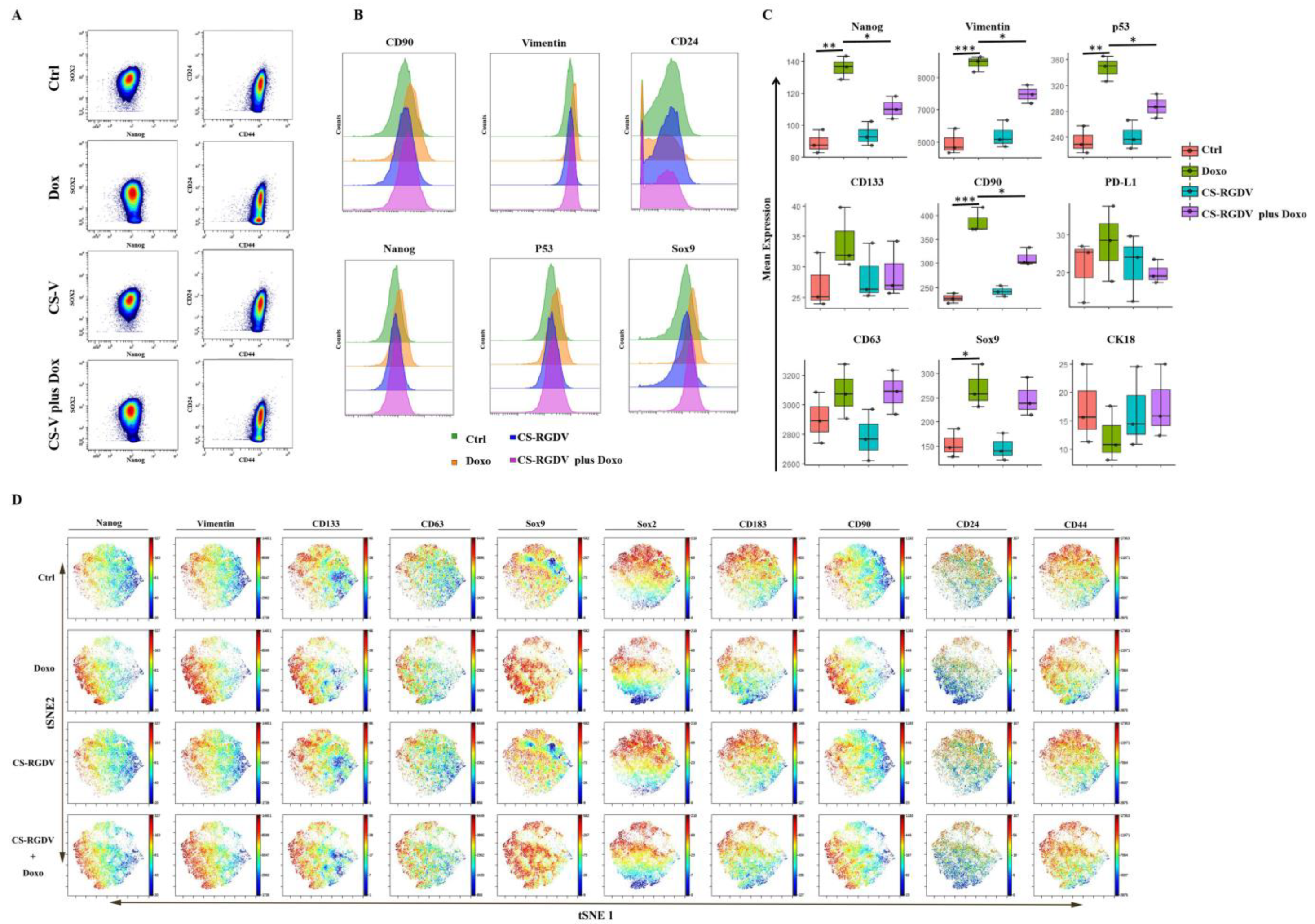
3.4. Visualizing CS-V-Promoted Doxorubicin against BCSCs at Single-Cell-Level
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Rossi, L.; Biagioni, C.; McCartney, A.; Migliaccio, I.; Curigliano, G.; Sanna, G.; Moretti, E.; Minisini, A.M.; Cinieri, S.; Tondini, C.; et al. Clinical outcomes after palbociclib with or without endocrine therapy in postmenopausal women with hormone receptor positive and HER2-negative metastatic breast cancer enrolled in the TREnd trial. Breast Cancer Res. 2019, 21, 71. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gupta, P.; Srivastava, S.K. Penfluridol overcomes paclitaxel resistance in metastatic breast cancer. Sci. Rep. 2019, 9, 5066. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.R. The role of chemotherapy and targeted agents in patients with metastatic breast cancer. Eur. J. Cancer 2011, 47, S38–S47. [Google Scholar] [CrossRef]
- Gupta, G.; Lee, C.D.; Guye, M.L.; van Sciver, R.E.; Lee, M.P.; Lafever, A.C.; Pang, A.; Tang-Tan, A.M.; Winston, J.S.; Samli, B.; et al. Unmet Clinical Need: Developing Prognostic Biomarkers and Precision Medicine to Forecast Early Tumor Relapse, Detect Chemo-Resistance and Improve Overall Survival in High-Risk Breast Cancer. Ann. Breast Cancer Ther. 2020, 4, 48–57. [Google Scholar] [PubMed]
- Wang, Q.; Jiang, J.; Ying, G.; Xie, X.Q.; Zhang, X.; Xu, W.; Zhang, X.; Song, E.; Bu, H.; Ping, Y.F.; et al. Tamoxifen enhances stemness and promotes metastasis of ERalpha36(+) breast cancer by upregulating ALDH1A1 in cancer cells. Cell Res. 2018, 28, 336–358. [Google Scholar] [CrossRef]
- Liu, L.; Yang, L.; Yan, W.; Zhai, J.; Pizzo, D.P.; Chu, P.; Chin, A.R.; Shen, M.; Dong, C.; Ruan, X.; et al. Chemotherapy Induces Breast Cancer Stemness in Association with Dysregulated Monocytosis. Clin. Cancer Res. 2018, 24, 2370–2382. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Dong, C.; Ruan, X.; Yan, W.; Cao, M.; Pizzo, D.; Wu, X.; Yang, L.; Liu, L.; Ren, X.; et al. Chemotherapy-Induced Extracellular Vesicle miRNAs Promote Breast Cancer Stemness by Targeting ONECUT2. Cancer Res. 2019, 79, 3608–3621. [Google Scholar] [CrossRef]
- Pastushenko, I.; Mauri, F.; Song, Y.; de Cock, F.; Meeusen, B.; Swedlund, B.; Impens, F.; Van Haver, D.; Opitz, M.; Thery, M.; et al. Fat1 deletion promotes hybrid EMT state, tumour stemness and metastasis. Nat. Cell Biol. 2021, 589, 448–455. [Google Scholar]
- Velazquez-Quesada, I.; Ruiz-Moreno, A.J.; Casique-Aguirre, D.; Aguirre-Alvarado, C.; Cortés-Mendoza, F.; de la Fuente-Granada, M.; García-Pérez, C.; Pérez-Tapia, S.M.; González-Arenas, A.; Segura-Cabrera, A.; et al. Pranlukast Antagonizes CD49f and Reduces Stemness in Triple-Negative Breast Cancer Cells. Drug Des. Dev. Ther. 2020, 14, 1799–1811. [Google Scholar] [CrossRef]
- Sridharan, S.; Howard, C.; Tilley, A.M.C.; Subramaniyan, B.; Tiwari, A.K.; Ruch, R.J.; Raman, D. Novel and Alternative Targets Against Breast Cancer Stemness to Combat Chemoresistance. Front. Oncol. 2019, 9, 1003. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Lesperance, J.; Seguin, L.; Gozo, M.; Kato, S.; Franovic, A.; Yebra, M.; Shattil, S.J.; Cheresh, D.A. Integrin alphavbeta3 drives slug activation and stemness in the pregnant and neoplastic mammary gland. Dev. Cell 2014, 30, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Seguin, L.; Weis, S.M.; Cheresh, D.A. Variety in the Tumor Microenvironment: Integrin Splicing Regulates Stemness. Cell Stem. Cell 2014, 14, 557–558. [Google Scholar] [CrossRef] [Green Version]
- Seguin, L.; Kato, S.; Franovic, A.; Camargo, M.F.; Lesperance, J.; Elliott, K.C.; Yebra, M.; Mielgo, A.; Lowy, A.M.; Husain, H.; et al. An integrin beta(3)-KRAS-RalB complex drives tumour stemness and resistance to EGFR inhibition. Nat. Cell Biol. 2014, 16, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Sarkhosh-Inanlou, R.; Imani, M.; Sam, M.R. The response of PIK3CA/KRAS-mutant colorectal cancer stem-like cells to RGD-peptide FraC produced by the strawberry anemone: A promising water-soluble peptide-based inhibitor of metastasis-driver gene CXCR4, stem cell regulatory genes and self-renewal. Biomed Pharm. 2020, 132, 110807. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.; Rajabi, S.; Tavirani, M.R.; Shokri, B.; Zarghi, A. Anti-Breast Cancer Activities of Ketoprofen-RGD Conjugate by Targeting Breast Cancer Stem-Like Cells and Parental Cells. Anticancer. Agents Med. Chem. 2021, 21, 1027–1036. [Google Scholar] [CrossRef]
- Cumber, A.J.; Forrester, J.A.; Foxwell, B.M.; Ross, W.C.; Thorpe, P.E. Preparation of antibody-toxin conjugates. Methods Enzym. 1985, 112, 207–225. [Google Scholar]
- Saber, M.M.; Bahrainian, S.; Dinarvand, R.; Atyabi, F. Targeted drug delivery of Sunitinib Malate to tumor blood vessels by cRGD-chiotosan-gold nanoparticles. Int. J. Pharm. 2017, 517, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.-S.; Tseng, H.-Y.; Chen, Y.-A.; Shen, P.-C.; Al Haq, A.T.; Chen, L.-M.; Tung, Y.-C.; Hsu, H.-L. MCT-1/miR-34a/IL-6/IL-6R signaling axis promotes EMT progression, cancer stemness and M2 macrophage polarization in triple-negative breast cancer. Mol. Cancer 2019, 18, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- YCho, H.; Ro, E.J.; Yoon, J.S.; Mizutani, T.; Kang, D.W.; Park, J.C.; Il, K.T.; Clevers, H.; Choi, K.Y. 5-FU promotes stemness of colorectal cancer via p53-mediated WNT/beta-catenin pathway activation. Nat. Commun. 2020, 11, 5321. [Google Scholar]
- Zhang, K.; Che, S.; Su, Z.; Zheng, S.; Zhang, H.; Yang, S.; Shanglin, Y.; Liu, J. CD90 promotes cell migration, viability and sphere-forming ability of hepatocellular carcinoma cells. Int. J. Mol. Med. 2018, 41, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Hu, S.B.; Wang, L.Y.; Zhang, X.; Zhou, X.; Yang, B.; Xiong, J.; Liu, N.; Li, Y.; Wu, Y.Z.; et al. Autophagy-dependent generation of Axin2+ cancer stem-like cells promotes hepatocarcinogenesis in liver cirrhosis. Oncogene 2017, 36, 6725–6737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaikh, M.V.; Kala, M.; Nivsarkar, M. CD90 a potential cancer stem cell marker and a therapeutic target. Cancer Biomark. 2016, 16, 301–307. [Google Scholar] [CrossRef]
- Brenet, M.; Martinez, S.; Perez-Nunez, R.; Perez, L.A.; Contreras, P.; Diaz, J.; Avalos, A.M.; Schneider, P.; Quest, A.; Leyton, L. Thy-1 (CD90)-Induced Metastatic Cancer Cell Migration and Invasion Are beta3 Integrin-Dependent and Involve a Ca(2+)/P2X7 Receptor Signaling Axis. Front. Cell Dev. Biol. 2020, 8, 592442. [Google Scholar] [CrossRef]
- Chen, W.-C.; Chang, Y.-S.; Hsu, H.-P.; Yen, M.-C.; Huang, H.-L.; Cho, C.-Y.; Wang, C.-Y.; Weng, T.-Y.; Lai, P.-T.; Chen, C.-S.; et al. Therapeutics targeting CD90-integrin-AMPK-CD133 signal axis in liver cancer. Oncotarget 2015, 6, 42923–42937. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.G.; Desai, K.; Prabhu, J.S.; Hari, P.S.; Remacle, J.; Sridhar, T.S. beta3 integrin promotes chemoresistance to epirubicin in MDA-MB-231 through repression of the pro-apoptotic protein, BAD. Exp. Cell Res. 2019, 346, 137–145. [Google Scholar] [CrossRef]
- Li, Y.; Reader, J.C.; Ma, X.; Kundu, N.; Kochel, T.; Fulton, A. Divergent roles of CXCR3 isoforms in promoting cancer stem-like cell survival and metastasis. Breast Cancer Res. Treat. 2015, 149, 403–415. [Google Scholar] [CrossRef]
- Spiwok, V.; Kříž, P. Time-Lagged t-Distributed Stochastic Neighbor Embedding (t-SNE) of Molecular Simulation Trajectories. Front. Mol. Biosci. 2020, 7, 132. [Google Scholar] [CrossRef]
- Belkina, A.C.; Ciccolella, C.O.; Anno, R.; Halpert, R.; Spidlen, J.; Snyder-Cappione, J.E. Automated optimized parameters for T-distributed stochastic neighbor embedding improve visualization and analysis of large datasets. Nat. Commun. 2019, 10, 5415. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Wang, F.; Tao, P. t-Distributed Stochastic Neighbor Embedding Method with the Least Information Loss for Macromolecular Simulations. J. Chem. Theory Comput. 2018, 14, 5499–5510. [Google Scholar] [CrossRef]
- Wang, C.; Wang, T.; Lv, D.; Li, L.; Yue, J.; Chen, H.-Z.; Xu, L. Acquired Resistance to EGFR TKIs Mediated by TGFβ1/Integrin β3 Signaling in EGFR-Mutant Lung Cancer. Mol. Cancer Ther. 2019, 18, 2357–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; Lv, D.; Wang, C.; Li, L.; Zhao, Q.; Chen, H.; Xu, L. Epigenetic silencing of miR-483-3p promotes acquired gefitinib resistance and EMT in EGFR-mutant NSCLC by targeting integrin beta3. Oncogene 2018, 37, 4300–4312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanker, A.; Estrada, M.V.; Bianchini, G.; Moore, P.D.; Zhao, J.; Cheng, F.; Koch, J.P.; Gianni, L.; Tyson, D.; Sánchez, V.; et al. Extracellular Matrix/Integrin Signaling Promotes Resistance to Combined Inhibition of HER2 and PI3K in HER2+ Breast Cancer. Cancer Res. 2017, 77, 3280–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.S.; Wendt, M.K. Integrin-mediated resistance to epidermal growth factor receptor-targeted therapy: An inflammatory situation. Breast Cancer Res. 2014, 16, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Yao, J.F.; Deng, X.F.; Zheng, X.D.; Jia, M.; Wang, Y.Q.; Huang, Y.; Zhu, J.H. 14, 15-EET induces breast cancer cell EMT and cisplatin resistance by up-regulating integrin alphavbeta3 and activating FAK/PI3K/AKT signaling. J. Exp. Clin. Cancer Res. 2018, 37, 23. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Mass | Antibody Clone | Source |
|---|---|---|---|
| NANOG | 142 | 23D2-3C6 | Biolegend |
| VIMENTIN | 143 | AF2105 | R&D |
| P53 | 144 | AF1043 | R&D |
| CD133 | 145 | MAB11331 | R&D |
| CD51/CD61 | 168 | 23C6 | Biolegend |
| CD63 | 146 | H5C6 | Biolegend |
| SOX9 | 153 | sc-166505 | Santa Cruz |
| SOX2 | 155 | MAB2018 | R&D |
| CD183 | 156 | MAB160 | R&D |
| Mitofusin | 149 | sc-166644 | Santa Cruz |
| CD90 | 159 | 5E10 | Biolegend |
| PDL1 | 209 | MAB1561 | R&D |
| INTEGRIN b7 | 162 | FIB27 | Biolegend |
| ASMA | 163 | MAB1420 | R&D |
| E-CADHERION | 164 | sc-8426 | Santa Cruz |
| NFATC1 | 154 | 7A6 | Biolegend |
| MUSASHI-1 | 155 | 3F8-E6-B9 | Biolegend |
| CK18 | 167 | sc-32329 | Santa Cruz |
| CD24 | 169 | ML5 | Biolegend |
| FOXO3 | 170 | sc-48348 | Santa Cruz |
| CD44 | 171 | C44Mab-5 | Biolegend |
| 53BP2 | 172 | sc-398311 | Santa Cruz |
| OCT4 | 146 | 3A2A20 | Biolegend |
| β-catenin | 147 | AF1329 | R&D |
| E2F1 | 148 | AF4825 | R&D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Lei, B.; Li, L.; Liu, J.; Li, Z.; Pang, Y.; Liu, T.; Li, Z. Single-Cell Proteomic Profiling Identifies Nanoparticle Enhanced Therapy for Triple Negative Breast Cancer Stem Cells. Cells 2021, 10, 2842. https://doi.org/10.3390/cells10112842
Wang W, Lei B, Li L, Liu J, Li Z, Pang Y, Liu T, Li Z. Single-Cell Proteomic Profiling Identifies Nanoparticle Enhanced Therapy for Triple Negative Breast Cancer Stem Cells. Cells. 2021; 10(11):2842. https://doi.org/10.3390/cells10112842
Chicago/Turabian StyleWang, Wenzheng, Bo Lei, Lin Li, Jianyu Liu, Zhihui Li, Yuheng Pang, Tong Liu, and Zhigao Li. 2021. "Single-Cell Proteomic Profiling Identifies Nanoparticle Enhanced Therapy for Triple Negative Breast Cancer Stem Cells" Cells 10, no. 11: 2842. https://doi.org/10.3390/cells10112842