
Characterization of the Radiation Desiccation Response Regulon of the Radioresistant Bacterium Deinococcus radiodurans by Integrative Genomic Analyses

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains; Plasmids; Oligonucleotides; Media
2.2. D. Radiodurans R1 Sequencing; Assembly and Annotation
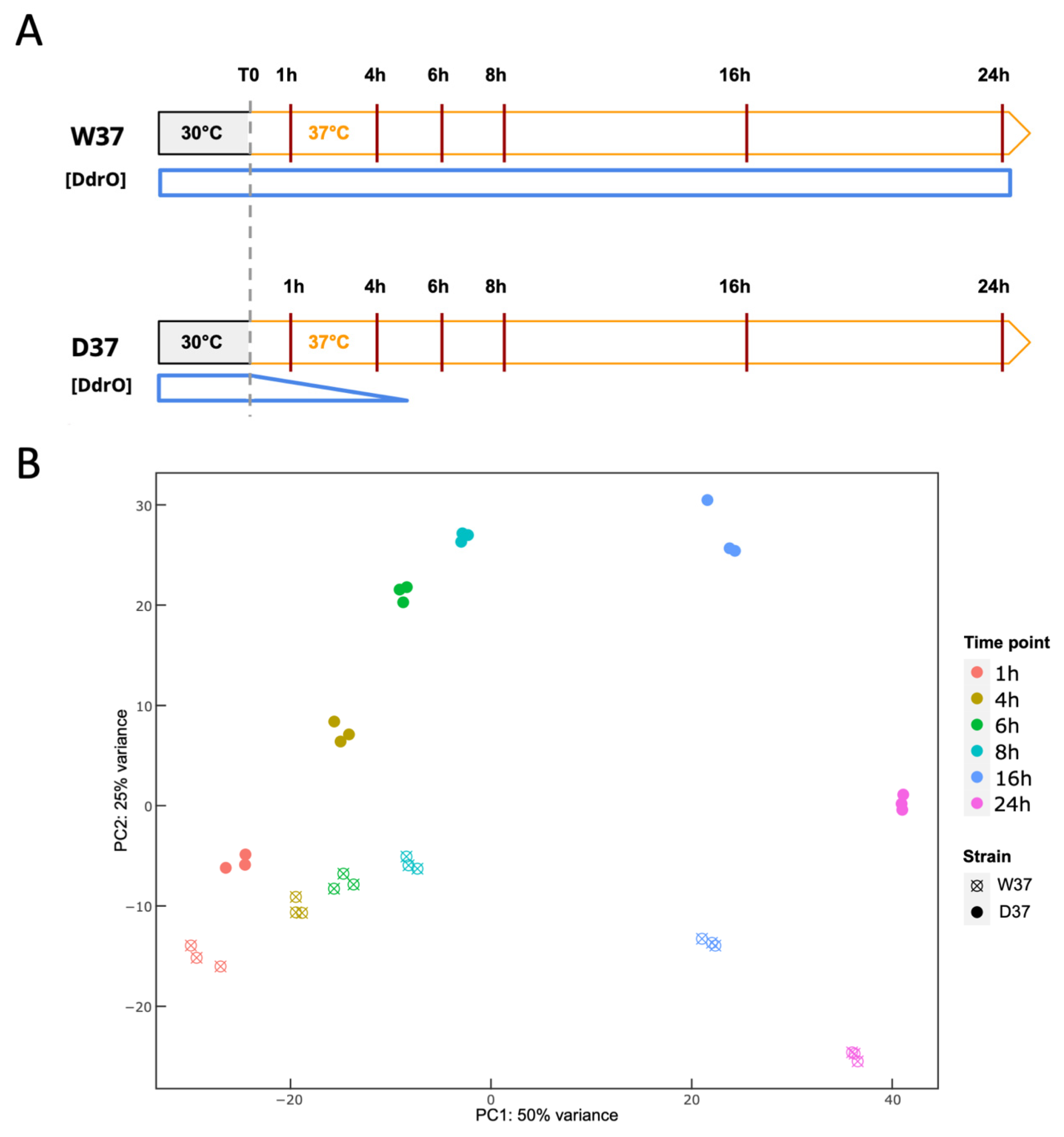
2.3. Time Course Experiment
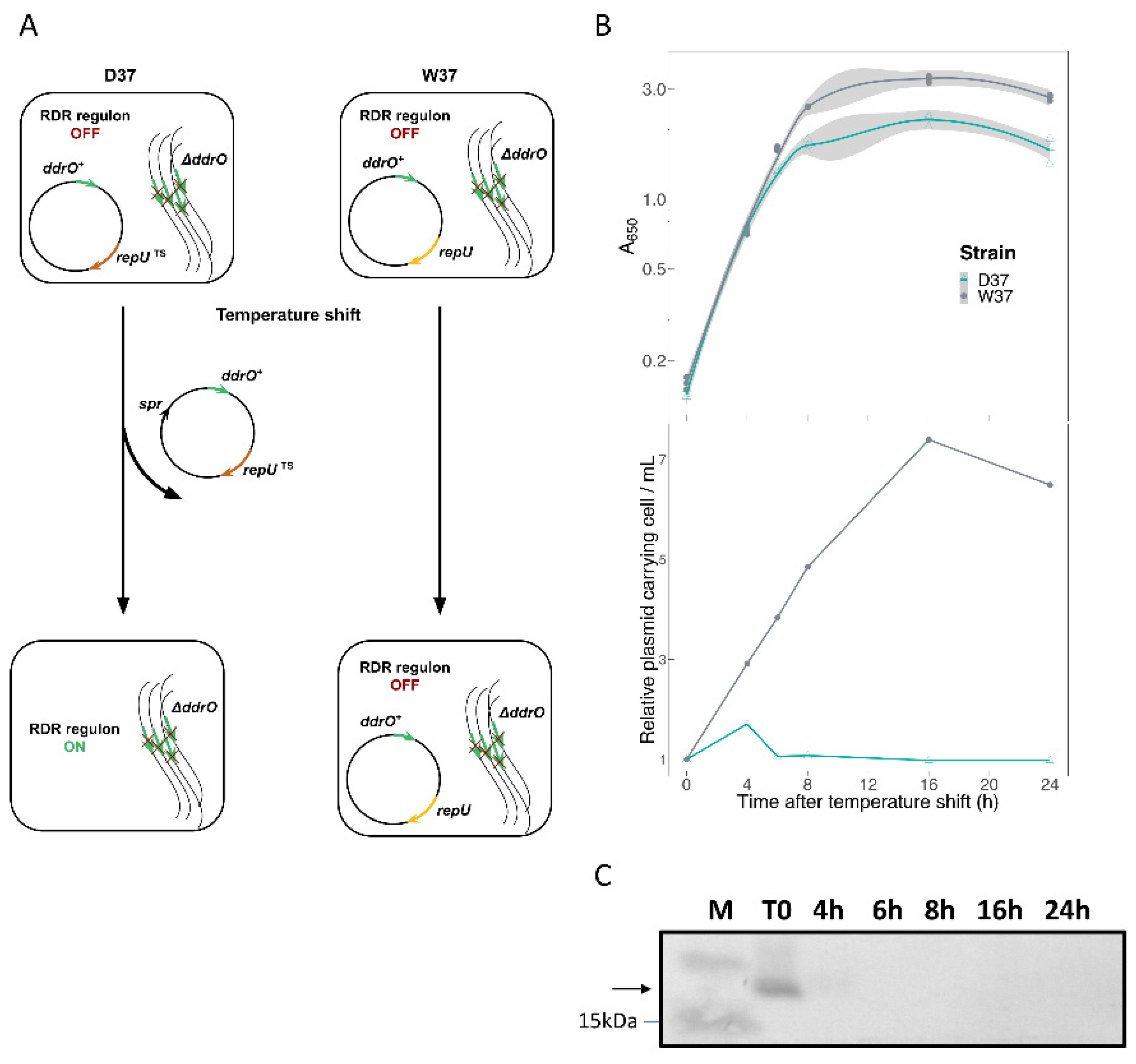
2.3.1. DdrO Depletion
2.3.2. RNA Extraction, cDNA Library Construction, and Sequencing
2.3.3. RNA-Seq Data Analysis
2.4. Western Blot Analysis
2.5. Chromatin Immunoprecipitation (ChIP)
2.6. ChIP-Seq
2.7. Western Blot Analysis of RDR Tagged-Proteins
2.8. Sensitivity Assay to DNA-Damaging Agents Mitomycin C and UVC
2.9. Deposition of Sequences and of Expression Data
3. Results
3.1. Genome Sequencing
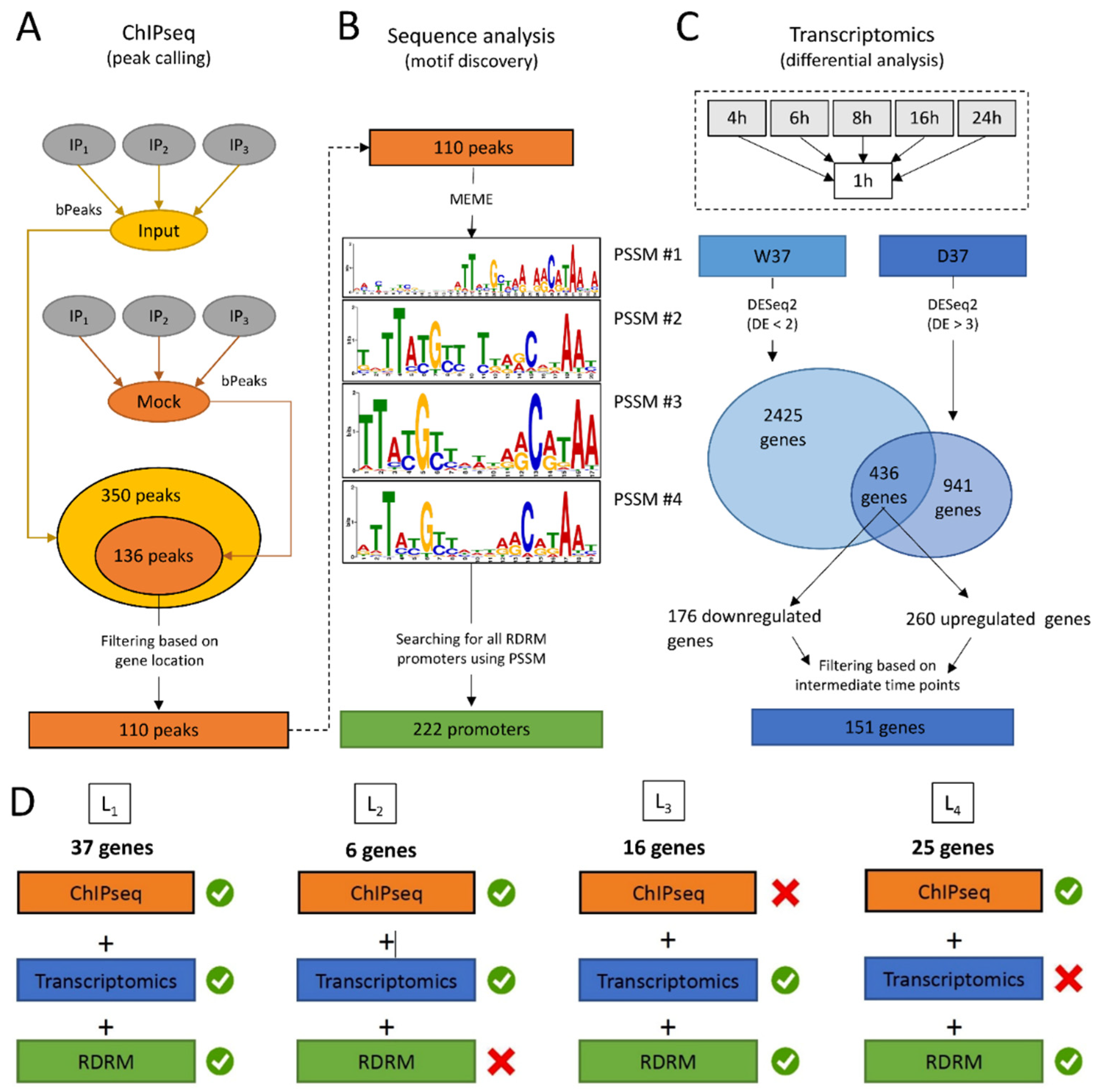
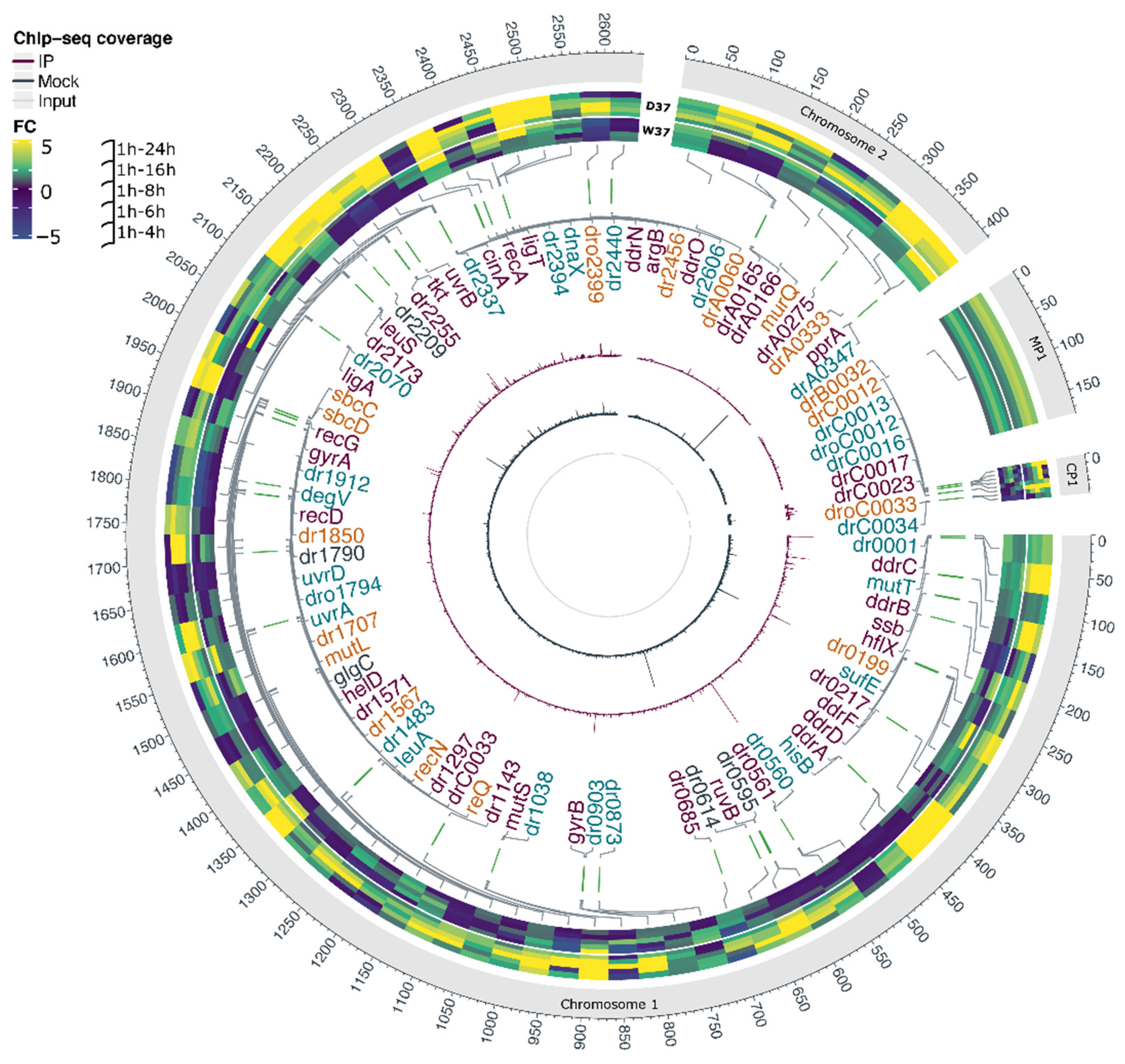
3.2. In Vivo Identification of DdrO Binding Sites by ChIP-Seq Assays
3.3. Transcriptome Analysis of D. radiodurans in Response to the Depletion of DdrO
3.4. Integration of the Data: The DdrO Map in D. radiodurans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID (White et al. 1999) [48] | Local ID | Genes | Definition | Position Relative to Start Codon | RDRM | DdrO Dependent Induction Fold Change 1h/Time Point  | Protein Induction with MMC/DdrO/IrrE Dependent | Predicted to RDR Regulon (Makarova et al. 2007; Blanchard et al. 2017) [19,33] | Previous Experiments |
|---|---|---|---|---|---|---|---|---|---|
| Replication, recombination, and repair | |||||||||
| DR0003 | DRO_0003 | ddrC | DdrC | −38 | GTTATGTCAAAAACATAATC |  | ND | X | 3, 5 |
| DR0070 | DRO_0070 | ddrB | single-stranded DNA-binding protein | −31 | TGTTATGTTATTTACGTAAG |  | ND | X | 1, 2, 3, 4, 5 |
| DR0100 | DRO_0099 | ssb | single-stranded DNA-binding protein | −118 | TTTTATGTCATTGACATAAT |  | ND | X | 1, 2, 3, 4, 5 |
| DR0326 | DRO_0323 | ddrD | DNA repair protein | −29 | ATTCTGCTAAAAACAGAATA |  | ND | X | 1, 2, 5 |
| DR0423 | DRO_0421 | ddrA | single-stranded DNA-binding protein | −23/−44 | ATTCTGTTCTAAACTAAAT/ TTTATGTCTTGACCGTAAT |  | ND | X | 1, 2, 3, 4, 5 |
| DR0596 | DRO_0596 | ruvB | Holliday junction DNA helicase RuvB | −28 | ATTTCGCAAATAGCGTAAT |  | ND | X | 1, 2, 3, 5 |
| DR0906 | DRO_0899 | gyrB | DNA topoisomerase IV subunit B | −157 | ATTCTGTAAGAGACGTAAT |  | ND | X | 1, 2, 3, 4, 5 |
| DR1039 | DRO_1033 | mutS | DNA mismatch repair protein MutS | −43 | GTTTCGCTCAGAACGTAAA |  | ND | X | 1, 2 |
| DR1916 | DRO_1894 | recG | ATP-dependent DNA helicase RecG | −17 | GTTACGCTGTGAGCGAAAT |  | yes/yes | 3 | |
| DR1572 | DRO_1552 | helD | DNA helicase | +9 | TTTATGTCTCTGGCAGAAC |  | no/no | 5 | |
| DR1902 | DRO_1880 | recD | exodeoxyribonuclease V | −46 | ATTACGCTGATGACATAAT |  | ND | 5 | |
| DR1913 | DRO_1891 | gyrA | DNA gyrase subunit A | −118 | ATTACGTGATTAACATAAT |  | ND | X | 1, 2, 3, 4, 5 |
| DR2069 | DRO_2042 | ligA | DNA ligase (NAD(+)) LigA | −47 | ATTCTGCCCTGAACCGAAC |  | yes/yes | 5 | |
| DR2275 | DRO_2249 | uvrB, putatively in operon with dr2276 | excinuclease ABC subunit B | −105 | CTTACGCTGTGGGCGTAAA |  | ND | X | 1,2,3,5 |
| DR2338 | DRO_2308 | cinA, first gene in operon with ligT and recA | Competence inducible protein A | −35 | GTTATGCTGCTAGCAGAAA |  | ND | X | 1,2,3,4,5 |
| DRA0346 | DRO_A0342 | pprA | DNA repair protein | −29 | AATCTGTTCAGGGCATAAT |  | ND | X | 1,2,3,4,5 |
| Regulation of transcription | |||||||||
| DR2574 | DRO_2545 | ddrO | transcriptional regulator | −153 | ATTCTGTATTGACCGTAGC |  | ND | X | 1,2,3,5 |
| Translation and post-translational modification | |||||||||
| DR0139 | DRO_0139 | hflX | GTPase HflX | −390 | GTTCTGTCCGGGGCGAAAC |  | yes/no | 4 | |
| DR2174 | DRO_2145 | leuS | leucine—tRNA ligase | −291 | CATATGTCATGAGCATAAC |  | no/no | ||
| DR2255 | DRO_2229 | Putatively n operon with dr2254 | GNAT family N-acetyltransferase | −209/−193 | AATACGCTAGGGGCGTAAA/ ATTCCGGTAAAGACAGAAT |  | no/no | 3 | |
| DR2441 | DRO_2415 | ddrN, putatively in operon with dr2442 | acetyltransferase | −109 | AATTTGTTATTTGCGAACT |  | yes/yes | 3,5 | |
| Metabolism and metabolic transport | |||||||||
| DR0217 | DRO_0217 | thiosulfate sulfurtransferase | +4 | ATTACGCCAAAGACGTGTT |  | no/no | 4 | ||
| DR0561 | DRO_0559 | sugar ABC transporter substrate-binding protein | −245 | GTTCAGGAAAAAACATAAC |  | no protein observed | |||
| DR1297 | DRO_1288 | ABC transporter | −131 | GTTACGCTCCTAAACAAAT |  | no protein observed | |||
| DR2256 | DRO_2230 | tkt | transketolase | −182/−199 | TTTACGCCCCTAGCGTATT/ ATTCTGTCTTTACCGGAAT |  | ND | X | 1,2,3 |
| DRA0275 | DRO_A0273 | cytochrome C6 | −93 | ATGTGACAAAGAGCGTAT |  | no/no | 5 | ||
| Unknown function | |||||||||
| DR0219 | DRO_0219 | ddrF | hypothetical protein | −43 | TGTTATGTTATATACGTAAA |  | ND | X | 1,2,3,5 |
| DR0685 | DRO_0681 | hypothetical protein | −37 | TCTTATGTTCTGAACGCTTT |  | no/no | |||
| DR1143 | DRO_1140 | hypothetical protein | −47 | GTTATGTTTTAAGCGTAAA |  | ND | X | 1,2,3,5 | |
| DR1571 | DRO_1551 | ABC transporter | −290 | GTTCTGCCAGAGACATAAA |  | no/no | |||
| DR2173 | DRO_2144 | hypothetical protein | −17 | GTTATGCTCATGACATATG |  | yes/yes | 3,5 | ||
| DRA0165 | DRO_A0167 | Putatively In operon with drA0166 | Conserved hypothetical protein | −83 | GGTTATGCTATTTACATAAC |  | yes/yes | 5 | |
| DRC0023 | DRO_C0021 | hypothetical protein | −193 | CTTTGTTCTGTTAGCCTAAC |  | no protein observed | 3 | ||
| ambiguous | |||||||||
| DRC0017 | DRO_C0017 | Putatively In operon with drC0018 | transposase | −248 | TAGTATGCTTCTGGCGTAGT |  | no protein observed | ||
| DR1296/DRC0033 | DRO_1287/ DRO_C0033 | transposase | −52 | AATATGTAAATAGCATAGT |  | no protein observed | |||
| Consensus motif TATGYYHTTARCRKA |  | ||||||||
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blasius, M.; Hubscher, U.; Sommer, S. Deinococcus radiodurans: What belongs to the survival kit? Crit. Rev. Biochem. Mol. Biol. 2008, 43, 221–238. [Google Scholar] [CrossRef]
- Confalonieri, F.; Sommer, S. Bacterial and archael resistance to ionizing radiation. J. Phys. 2011, 261, 012005. [Google Scholar] [CrossRef]
- Daly, M.J. Death by protein damage in irradiated cells. DNA Repair 2012, 11, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [Green Version]
- Zahradka, K.; Slade, D.; Bailone, A.; Sommer, S.; Averbeck, D.; Petranovic, M.; Lindner, A.B.; Radman, M. Reassembly of shattered chromosomes in Deinococcus radiodurans. Nature 2006, 443, 569–573. [Google Scholar] [CrossRef]
- Bouthier de la Tour, C.; Boisnard, S.; Norais, C.; Toueille, M.; Bentchikou, E.; Vannier, F.; Cox, M.M.; Sommer, S.; Servant, P. The deinococcal DdrB protein is involved in an early step of DNA double strand break repair and in plasmid transformation through its single-strand annealing activity. DNA Repair 2011, 10, 1223–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, D.; Lindner, A.B.; Paul, G.; Radman, M. Recombination and replication in DNA repair of heavily irradiated Deinococcus radiodurans. Cell 2009, 136, 1044–1055. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Lu, H.; Wang, L.; Chen, H.; Xu, Z.; Hu, Y.; Tian, B.; Hua, Y. DdrB stimulates single-stranded DNA annealing and facilitates RecA-independent DNA repair in Deinococcus radiodurans. DNA Repair 2010, 9, 805–812. [Google Scholar] [CrossRef]
- Floc’h, K.; Lacroix, F.; Servant, P.; Wong, Y.S.; Kleman, J.P.; Bourgeois, D.; Timmins, J. Cell morphology and nucleoid dynamics in dividing Deinococcus radiodurans. Nat. Commun. 2019, 10, 3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, J.M.; Battista, J.R. A ring-like nucleoid is not necessary for radioresistance in the Deinococcaceae. BMC Microbiol. 2005, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Kiang, J.G.; Fukumoto, R.; Lee, D.Y.; Wehr, N.B.; Viteri, G.A.; Berlett, B.S.; Levine, R.L. Small-molecule antioxidant proteome-shields in Deinococcus radiodurans. PLoS ONE 2010, 5, e12570. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Omelchenko, M.V.; Beliaev, A.S.; Venkateswaran, A.; Stair, J.; Wu, L.; Thompson, D.K.; Xu, D.; Rogozin, I.B.; et al. Transcriptome dynamics of Deinococcus radiodurans recovering from ionizing radiation. Proc. Natl. Acad. Sci. USA 2003, 100, 4191–4196. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Earl, A.M.; Howell, H.A.; Park, M.J.; Eisen, J.A.; Peterson, S.N.; Battista, J.R. Analysis of Deinococcus radiodurans’s transcriptional response to ionizing radiation and desiccation reveals novel proteins that contribute to extreme radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzer, K.N. DNA damage responses in prokaryotes: Regulating gene expression, modulating growth patterns, and manipulating replication forks. Cold Spring Harb. Perspect Biol. 2013, 5, a012674. [Google Scholar] [CrossRef]
- Bonacossa de Almeida, C.; Coste, G.; Sommer, S.; Bailone, A. Quantification of RecA protein in Deinococcus radiodurans reveals involvement of RecA, but not LexA, in its regulation. Mol. Genet. Genom. 2002, 268, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Narumi, I.; Satoh, K.; Kikuchi, M.; Funayama, T.; Yanagisawa, T.; Kobayashi, Y.; Watanabe, H.; Yamamoto, K. The LexA protein from Deinococcus radiodurans is not involved in RecA induction following gamma irradiation. J. Bacteriol. 2001, 183, 6951–6956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludanyi, M.; Blanchard, L.; Dulermo, R.; Brandelet, G.; Bellanger, L.; Pignol, D.; Lemaire, D.; de Groot, A. Radiation response in Deinococcus deserti: IrrE is a metalloprotease that cleaves repressor protein DdrO. Mol. Microbiol. 2014, 94, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Vujicic-Zagar, A.; Dulermo, R.; Le Gorrec, M.; Vannier, F.; Servant, P.; Sommer, S.; de Groot, A.; Serre, L. Crystal structure of the IrrE protein, a central regulator of DNA damage repair in deinococcaceae. J. Mol. Biol. 2009, 386, 704–716. [Google Scholar] [CrossRef]
- Blanchard, L.; Guerin, P.; Roche, D.; Cruveiller, S.; Pignol, D.; Vallenet, D.; Armengaud, J.; de Groot, A. Conservation and diversity of the IrrE/DdrO-controlled radiation response in radiation-resistant Deinococcus bacteria. Microbiologyopen 2017, 6, e00477. [Google Scholar] [CrossRef]
- Devigne, A.; Ithurbide, S.; Bouthier de la Tour, C.; Passot, F.; Mathieu, M.; Sommer, S.; Servant, P. DdrO is an essential protein that regulates the radiation desiccation response and the apoptotic-like cell death in the radioresistant Deinococcus radiodurans bacterium. Mol. Microbiol. 2015, 96, 1069–1084. [Google Scholar] [CrossRef]
- Bouthier de la Tour, C.; Mathieu, M.; Meyer, L.; Dupaigne, P.; Passot, F.; Servant, P.; Sommer, S.; Le Cam, E.; Confalonieri, F. In vivo and in vitro characterization of DdrC, a DNA damage response protein in Deinococcus radiodurans bacterium. PLoS ONE 2017, 12, e0177751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, A.M.; Mohundro, M.M.; Mian, I.S.; Battista, J.R. The IrrE protein of Deinococcus radiodurans R1 is a novel regulator of recA expression. J. Bacteriol. 2002, 18, 6216–6224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.; Narumi, I.; Gao, G.; Tian, B.; Satoh, K.; Kitayama, S.; Shen, B. PprI: A general switch responsible for extreme radioresistance of Deinococcus radiodurans. Biochem. Biophys. Res. Commun. 2003, 306, 354–360. [Google Scholar] [CrossRef]
- Lu, H.; Chen, H.; Xu, G.; Shah, A.M.; Hua, Y. DNA binding is essential for PprI function in response to radiation damage in Deinococcus radiodurans. DNA Repair 2012, 11, 139–145. [Google Scholar] [CrossRef]
- Magerand, R.; Rey, P.; Blanchard, L.; de Groot, A. Redox signaling through zinc activates the radiation response in Deinococcus bacteria. Sci. Rep. 2021, 11, 4528. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Q.; Lu, H.; Lin, L.; Wang, L.; Xu, H.; Cui, X.; Zhang, H.; Li, T.; Hua, Y. Protease activity of PprI facilitates DNA damage response: Mn2+-dependence and substrate sequence-specificity of the proteolytic reaction. PLoS ONE 2015, 10, e0122071. [Google Scholar] [CrossRef]
- Narasimha, A.; Basu, B. New insights into the activation of radiation desiccation response regulon in Deinococcus radiodurans. J. Biosci. 2021, 46, 10. [Google Scholar] [CrossRef]
- de Groot, A.; Siponen, M.I.; Magerand, R.; Eugenie, N.; Martin-Arevalillo, R.; Doloy, J.; Lemaire, D.; Brandelet, G.; Parcy, F.; Dumas, R.; et al. Crystal structure of the transcriptional repressor DdrO: Insight into the metalloprotease/repressor-controlled radiation response in Deinococcus. Nucleic. Acids. Res. 2019, 47, 11403–11417. [Google Scholar] [CrossRef]
- Lu, H.; Wang, L.; Li, S.; Pan, C.; Cheng, K.; Luo, Y.; Xu, H.; Tian, B.; Zhao, Y.; Hua, Y. Structure and DNA damage-dependent derepression mechanism for the XRE family member DG-DdrO. Nucleic. Acids. Res. 2019, 47, 9925–9933. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Jung, J.H.; Blanchard, L.; de Groot, A. Conservation and diversity of radiation and oxidative stress resistance mechanisms in Deinococcus species. FEMS Microbiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchard, L.; de Groot, A. Coexistence of SOS-dependent and SOS-independent regulation of DNA repair genes in radiation-resistant Deinococcus bacteria. Cells 2021, 10, 924. [Google Scholar] [CrossRef] [PubMed]
- de Groot, A.; Dulermo, R.; Ortet, P.; Blanchard, L.; Guerin, P.; Fernandez, B.; Vacherie, B.; Dossat, C.; Jolivet, E.; Siguier, P.; et al. Alliance of proteomics and genomics to unravel the specificities of Sahara bacterium Deinococcus deserti. PLoS Genet. 2009, 5, e1000434. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Lapidus, A.; Copeland, A.; Kim, E.; Land, M.; et al. Deinococcus geothermalis: The pool of extreme radiation resistance genes shrinks. PLoS ONE 2007, 2, e955. [Google Scholar] [CrossRef] [PubMed]
- Anaganti, N.; Basu, B.; Mukhopadhyaya, R.; Apte, S.K. Proximity of radiation desiccation response motif to the core promoter is essential for basal repression as well as gamma radiation-induced gyrB gene expression in Deinococcus radiodurans. Gene 2017, 615, 8–17. [Google Scholar] [CrossRef]
- Meima, R.; Lidstrom, M.E. Characterization of the minimal replicon of a cryptic Deinococcus radiodurans SARK plasmid and development of versatile Escherichia coli-D. radiodurans shuttle vectors. Appl. Environ. Microbiol. 2000, 66, 3856–3867. [Google Scholar] [CrossRef] [Green Version]
- Mennecier, S.; Coste, G.; Servant, P.; Bailone, A.; Sommer, S. Mismatch repair ensures fidelity of replication and recombination in the radioresistant organism Deinococcus radiodurans. Mol. Genet. Genom. 2004, 272, 460–469. [Google Scholar] [CrossRef]
- Dardalhon-Samsonoff, M.; Averbeck, D. DNA-membrane complex restoration in Micrococcus radiodurans after X-irradiation: Relation to repair, DNA synthesis and DNA degradation. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1980, 38, 31–52. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome. Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Subgroup Genome Project Data Processing. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; McCarthy, S.A. BCFtools/csq: Haplotype-aware variant consequences. Bioinformatics 2017, 33, 2037–2039. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, D.; LoCascio, P.F.; Hauser, L.J.; Uberbacher, E.C. Gene and translation initiation site prediction in metagenomic sequences. Bioinformatics 2012, 28, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome. Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Solovyev, V.; Salamov, A.; Seledtsov, I.; Vorobyev, D.; Bachinsky, A. Automatic annotation of bacterial community sequences and application to infections diagnostic. In Proceedings of the International Conference on Bioinformatics Models, Rome, Italy, 26–29 January 2011; pp. 346–353. [Google Scholar] [CrossRef] [Green Version]
- White, O.; Eisen, J.A.; Heidelberg, J.F.; Hickey, E.K.; Peterson, J.D.; Dodson, R.J.; Haft, D.H.; Gwinn, M.L.; Nelson, W.C.; Richardson, D.L.; et al. Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Hua, Y. Improved complete genome sequence of the extremely radioresistant bacterium Deinococcus radiodurans R1 obtained using PacBio single-molecule sequencing. Genome Announc. 2016, 4, e00886-16. [Google Scholar] [CrossRef] [Green Version]
- Merhej, J.; Frigo, A.; Le Crom, S.; Camadro, J.M.; Devaux, F.; Lelandais, G. bPeaks: A bioinformatics tool to detect transcription factor binding sites from ChIPseq data in yeasts and other organisms with small genomes. Yeast 2014, 31, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic. Acids. Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.H.; Bouthier de la Tour, C.; Toueille, M.; Vannier, F.; Sommer, S.; Servant, P. The essential histone-like protein HU plays a major role in Deinococcus radiodurans nucleoid compaction. Mol. Microbiol. 2009, 73, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, H.; Xu, X.; Wang, L.; Tian, B.; Hua, Y. Characteristics of dr1790 disruptant and its functional analysis in Deinococcus radiodurans. Braz. J. Microbiol. 2015, 46, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Basu, B.; Apte, S.K. Gamma radiation-induced proteome of Deinococcus radiodurans primarily targets DNA repair and oxidative stress alleviation. Mol. Cell Proteom. 2012, 11, M111.011734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, R.G.; Rudolph, C.J. 25 years on and no end in sight: A perspective on the role of RecG protein. Curr. Genet. 2016, 62, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Kapoor, S.; Thakur, K.G. Bacillus subtilis HelD, an RNA polymerase interacting helicase, forms amyloid-like fibrils. Front Microbiol. 2018, 9, 1934. [Google Scholar] [CrossRef] [Green Version]
- Newing, T.P.; Oakley, A.J.; Miller, M.; Dawson, C.J.; Brown, S.H.J.; Bouwer, J.C.; Tolun, G.; Lewis, P.J. Molecular basis for RNA polymerase-dependent transcription complex recycling by the helicase-like motor protein HelD. Nat. Commun. 2020, 11, 6420. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, W.; Zhao, Y.; Xu, H.; Hua, Y. Involvement of RecG in H2O2-induced damage repair in Deinococcus radiodurans. Can. J. Microbiol. 2009, 55, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.W.; Kim, M.K.; Zhao, L.; Yang, S.K.; Jung, J.H.; Lim, H.M.; Lim, S. Effects of conserved wedge domain residues on DNA binding activity of Deinococcus radiodurans RecG helicase. Front Genet. 2021, 12, 634615. [Google Scholar] [CrossRef]
- Cao, Z.; Julin, D.A. Characterization in vitro and in vivo of the DNA helicase encoded by Deinococcus radiodurans locus DR1572. DNA Repair 2009, 8, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.; Day, J.; Bowater, R. Bacterial DNA ligases. Mol. Microbiol. 2001, 40, 1241–1248. [Google Scholar] [CrossRef]
- Macek, B.; Forchhammer, K.; Hardouin, J.; Weber-Ban, E.; Grangeasse, C.; Mijakovic, I. Protein post-translational modifications in bacteria. Nat. Rev. Microbiol. 2019, 17, 651–664. [Google Scholar] [CrossRef]
- Carabetta, V.J.; Cristea, I.M. Regulation, function, and detection of protein acetylation in bacteria. J. Bacteriol. 2017, 199, e00107-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhou, A.; Li, S.; Ni, J.; Tao, J.; Lu, J.; Wan, B.; Li, S.; Zhang, J.; Zhao, S.; et al. Reversible lysine acetylation is involved in DNA replication initiation by regulating activities of initiator DnaA in Escherichia coli. Sci. Rep. 2016, 6, 30837. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Padmanabhan, B.; Anand, C.; Nagaraja, V. Lysine acetylation of the Mycobacterium tuberculosis HU protein modulates its DNA binding and genome organization. Mol. Microbiol. 2016, 100, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Toueille, M.; Mirabella, B.; Guerin, P.; Bouthier de la Tour, C.; Boisnard, S.; Nguyen, H.H.; Blanchard, L.; Servant, P.; de Groot, A.; Sommer, S.; et al. A comparative proteomic approach to better define Deinococcus nucleoid specificities. J. Proteom. 2012, 75, 2588–2600. [Google Scholar] [CrossRef]
- Mennecier, S.; Servant, P.; Coste, G.; Bailone, A.; Sommer, S. Mutagenesis via IS transposition in Deinococcus radiodurans. Mol. Microbiol. 2006, 59, 317–325. [Google Scholar] [CrossRef]
- Kuan, C.T.; Tessman, I. LexA protein of Escherichia coli represses expression of the Tn5 transposase gene. J. Bacteriol. 1991, 173, 6406–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Wendt, J.C.; Shanmugam, K.T. Identification of a new gene, molR, essential for utilization of molybdate by Escherichia coli. J. Bacteriol. 1990, 172, 2079–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, J.; Yakhnina, A.A.; Gitai, Z. BapE DNA endonuclease induces an apoptotic-like response to DNA damage in Caulobacter. Proc. Natl. Acad. Sci. USA 2012, 109, 18096–18101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.L.; Wang, H.H. The role of genome accessibility in transcription factor binding in bacteria. PLoS Comput. Biol. 2016, 12, e1004891. [Google Scholar] [CrossRef]
- Meyer, L.; Coste, G.; Sommer, S.; Oberto, J.; Confalonieri, F.; Servant, P.; Pasternak, C. DdrI, a cAMP receptor protein family member, acts as a major regulator for adaptation of Deinococcus radiodurans to various stresses. J. Bacteriol. 2018, 200, e00129-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using Tablet for visual exploration of second-generation sequencing data. Brief Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef] [PubMed]








| Replicon | ID | Percentage of Pairs of Orthologs * | CDS | Total CDS | Nt. | GC% | |
|---|---|---|---|---|---|---|---|
| Hua and Hua (2016) | Chr 1 | CP015081 | 96.43 | 2523 | 3079 | 2,646,742 | 67.07 |
| Chr 2 | CP015082 | 96.02 | 352 | 433,133 | 66.77 | ||
| Megaplasmid | CP015083 | 81.05 | 153 | 203,183 | 62.98 | ||
| Plasmid | CP015084 | 62.75 | 51 | 61,707 | 56.55 | ||
| White et al., (1999) | Chr 1 | DRA1 | 84.37 | 2629 | 3181 | 2,648,638 | 67.01 |
| Chr 2 | DRA2 | 85.05 | 368 | 412,348 | 66.69 | ||
| Megaplasmid | DRA3 | 77.24 | 145 | 177,466 | 63.19 | ||
| Plasmid | DRA4 | 61.54 | 39 | 45,704 | 56.15 | ||
| This work | Chr 1 | DRO | 100 | 2594 | 2,644,251 | 67.08 | |
| Chr 2 | DRO_A | 100 | 364 | 3147 | 412,138 | 66.65 | |
| Megaplasmid | DRO_B | 100 | 148 | 177,322 | 63.21 | ||
| Plasmid | DRO_C | 100 | 41 | 45,508 | 56.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eugénie, N.; Zivanovic, Y.; Lelandais, G.; Coste, G.; Bouthier de la Tour, C.; Bentchikou, E.; Servant, P.; Confalonieri, F. Characterization of the Radiation Desiccation Response Regulon of the Radioresistant Bacterium Deinococcus radiodurans by Integrative Genomic Analyses. Cells 2021, 10, 2536. https://doi.org/10.3390/cells10102536
Eugénie N, Zivanovic Y, Lelandais G, Coste G, Bouthier de la Tour C, Bentchikou E, Servant P, Confalonieri F. Characterization of the Radiation Desiccation Response Regulon of the Radioresistant Bacterium Deinococcus radiodurans by Integrative Genomic Analyses. Cells. 2021; 10(10):2536. https://doi.org/10.3390/cells10102536
Chicago/Turabian StyleEugénie, Nicolas, Yvan Zivanovic, Gaelle Lelandais, Geneviève Coste, Claire Bouthier de la Tour, Esma Bentchikou, Pascale Servant, and Fabrice Confalonieri. 2021. "Characterization of the Radiation Desiccation Response Regulon of the Radioresistant Bacterium Deinococcus radiodurans by Integrative Genomic Analyses" Cells 10, no. 10: 2536. https://doi.org/10.3390/cells10102536