Biostimulants and Microorganisms Boost the Nutritional Composition of Buckwheat (Fagopyrum esculentum Moench) Sprouts


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Sampling Preparation and Chemical Analysis
hemicellulose (HCEL) = NDF − ADF
2.3. Statistical Analyses
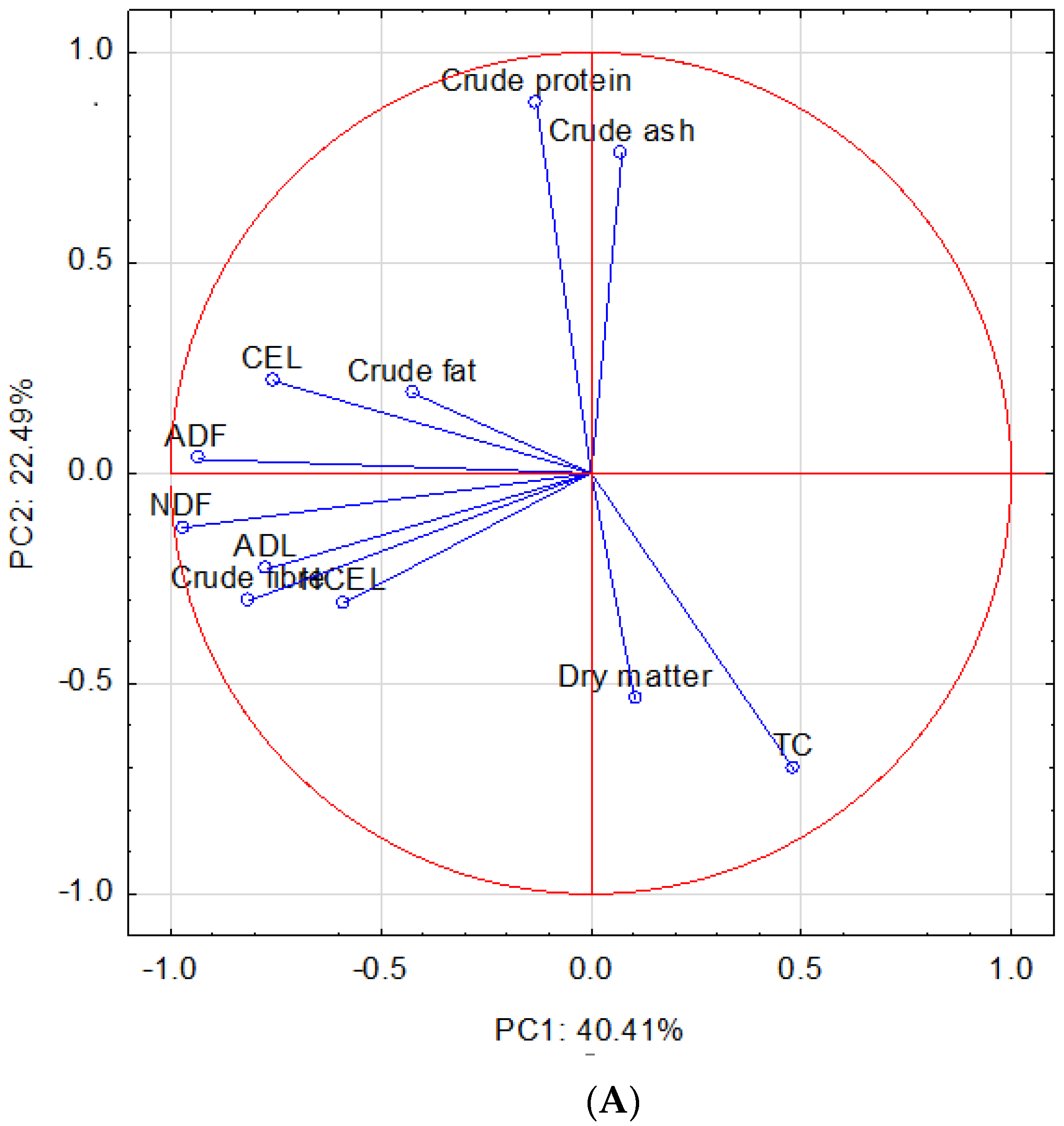
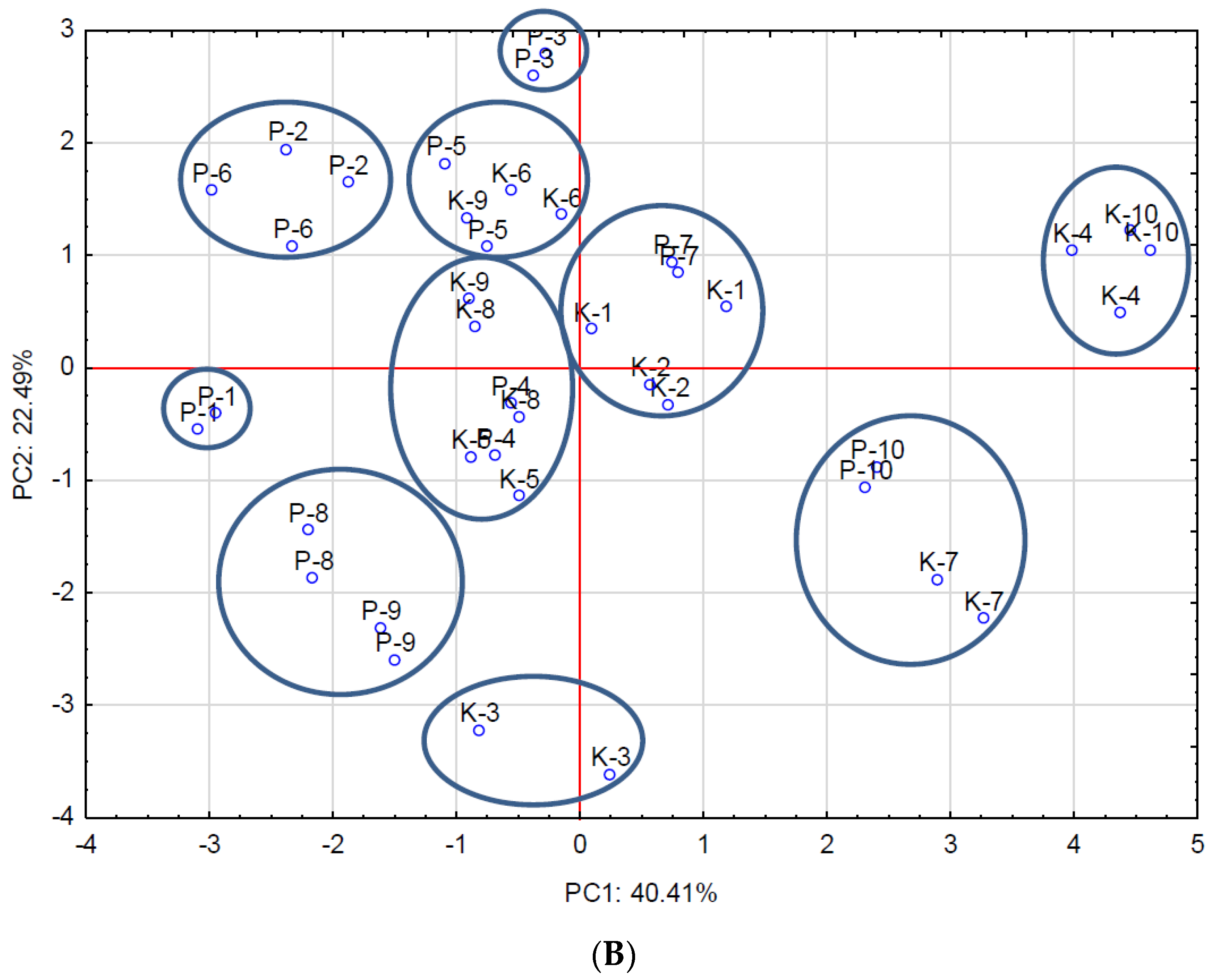
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ebert, A.W. Potential of underutilized traditional vegetables and legume crops to contribute to food and nutritional security, income and more sustainable production systems. Sustainability 2014, 6, 319–335. [Google Scholar] [CrossRef]
- Kahane, R.; Hodgkin, T.; Jaenicke, H.; Hoogendoorn, C.; Hermann, J.; Keatinge, J.; Hughes, J.D.; Looney, N. Agrobiodiversity for food security, health and income. Agron. Sustain. Dev. 2013, 33, 671–693. [Google Scholar] [CrossRef] [Green Version]
- Westhoek, H.; Lesschen, P.J.; Rood, T.; Wagner, S.; De Marco, A.; Murphy-Bokern, D.; Leip, A.; van Grinsven, H.; Sutton, M.A.; Oenema, O. Food choices, health and environment: Effects of cutting Europe’s meat and dairy intake. Global Environ. Chang. 2014, 26, 196–205. [Google Scholar] [CrossRef]
- Ebert, A.W. Sprouts, Microgreens, and Edible Flowers: The Potential for High Value Specialty Produce in Asia. In Proceedings of the SEAVEG Regional Symposium. High Value Vegetables in Southeast Asia: Production, Supply and Demand, Chiang Mai, Thailand, 24–26 January 2012. [Google Scholar]
- Di Gioia, F.; Santamaria, P. The nutritional properties of microgreens. In Microgreens; Di Gioia, F., Santamaria, P., Eds.; Eco-Logica: Bari, Italy, 2015; p. 47. [Google Scholar]
- Kim, S.L.; Kim, S.K.; Park, C.H. Introduction and nutritional evaluation of buckwheat sprout as a new vegetable. Food Res. Int. 2004, 37, 319–327. [Google Scholar] [CrossRef]
- Marton, M.; Mandoki, Z.; Csapo-Kiss, Z.; Csapo, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae Aliment. 2010, 3, 81–117. [Google Scholar]
- Yang, Y.; Meier, F.; Ann Lo, J.; Yuan, W.; Lee Pei Sze, V.; Chung, H.J.; Yuk, H.G. Overview of recent events in the microbiological safety of sprouts and new intervention technologies. Compr. Rev. Food Sci. Food Saf. 2013, 12, 265–280. [Google Scholar] [CrossRef]
- Frederiks, C.; Wesseler, J.H.H. A comparison of EU and US regulatory frameworks for the active substance registration of microbial biological control agents. Pest. Manag. Sci. 2019, 78, 87–103. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Bulgari, R.; Trivellini, A.; Ferrante, A. Effects of two doses of organic extract-based biostimulant on greenhouse lettuce grown under increasing NaCl concentrations. Front. Plant Sci. 2019, 9, 1870. [Google Scholar] [CrossRef]
- Kałużewicz, A.; Gąsecka, M.; Spiżewski, T. Influence of biostimulants on phenolic content in broccoli heads directly after harvest and after storage. Folia Hort. 2017, 29, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Briatia, X.; Jomduang, S.; Park, C.H.; Lumyong, S.; Kanpiengjai, A.; Khanongnuch, C. Enhancing growth of buckwheat sprouts and mircogreens by endophytic bacterium inoculation. Int. J. Agric. Biol. 2017, 19, 374–380. [Google Scholar] [CrossRef]
- Gerbore, J.; Vallance, J.; Yacob, A.; Delmotte, F.; Grizard, D.; Regnault-Roger, C.; Rey, P. Characterization of Phytium oligandrum populations that colonize the rhizosphere of vines form Bordeaux region. FEMS Microbiol. Ecol. 2014, 90, 153–167. [Google Scholar] [CrossRef]
- Singh, A.; Gupta, R.; Pandey, R. Rice seed priming with picomolar rutin enhance rhizospheric Bacillus subtilis CIM colonization and plant growth. PLoS ONE 2016, 11, e01466013. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Hortic. Sci. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Hashem, H.A.; Mansour, H.A.; El-Khawas, S.A.; Hassanein, R.A. The potentiality of marine macro-algae as bio-fertilizers to improve the productivity and salt stress tolerance of canola (Brassica napus L.) plants. Agronomy 2019, 9, 146. [Google Scholar]
- Chanthini, K.M.-P.; Stanley-Raja, V.; Thanigaivel, A.; Karthi, S.; Palanikani, R.; Shyam Sundar, N.; Sivanesh, H.; Soranam, R.; Senthil-Nathan, S. Sustainable agronomic strategies for enhancing the yield and nutritional quality of wild tomato, Solanum Lycopersicum (l) var Cerasiforme Mill. Agronomy 2019, 9, 311. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hort. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Baranowska, A.J. Impact of Growth Biostimulators and Herbicide on Edible Potato Yield. Acta Agroph. 2018, 25, 385–396. [Google Scholar] [CrossRef]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants Research in Some Horticultural Plant Species—A Review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 19th ed.; AOAC: Washington, DC, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Liu, C.L.; Chen, Y.S.; Yang, J.H.; Chiang, B.H. Antioxidant activity of tartary (Fagopyrum tataricum (L.) Gaertn.) and common (Fagopyrum esculentum Moench) buckwheat sprouts. J. Agric. Food Chem. 2008, 56, 173–178. [Google Scholar] [CrossRef]
- Kim, S.L.; Son, Y.K.; Hwang, J.J.; Kim, S.K.; Hur, H.S.; Park, C.H. Development and utilization of buckwheat sprouts as functional vegetables. Fagopyrum 2001, 18, 49–54. [Google Scholar]
- Paredes-Lopez, O.; Mora-Escobedo, R. Germination of amaranth seeds: Effects on nutrient composition and color. J. Food Sci. 1989, 54, 761–762. [Google Scholar] [CrossRef]
- Gimenez-Bastida, J.A.; Zieliński, H. Buckwheat as a functional food and its effects on health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef]
- Dziadek, K.; Kopeć, A.; Piątkowska, E.; Leszczyńska, T.; Pisulewska, E.; Witkowicz, R.; Bystrowska, B.; Francik, R. Identification of polyphenolic compounds and determination of antioxidant in extracts and infusions of buckwheat leaves. Eur. Food Res. Technol. 2017, 244, 333–343. [Google Scholar] [CrossRef]
- Guillon, F.; Champ, M. Structural and physical properties of dietary fibres, and consequences of processing on human physiology. Food Res. Int. 2000, 33, 233–245. [Google Scholar] [CrossRef]
- Linh, N.T.N.; Khoa, D.V.A.; Halas, V. Buckwheat as valuable feed and food resource. NJMBS 2014, 3, 1–8. [Google Scholar]
- Yang, H.J.; Lim, J.H.; Park, K.J.; Kang, S.; Kim, D.S.; Park, S. Methyl jasmolate treated buckwheat sprout powder enhances glucose metabolism by potentiating hepatic insulin signaling in estrogen-deficient rats. Nutrition 2016, 32, 129–137. [Google Scholar] [CrossRef]
- Yiming, Z.; Hong, W.; Linlin, C.; Xiaoli, Z.; Wen, T.; Xinli, S. Evolution of nutrient ingredients in tartary buckwheat seeds during germination. Food Chem. 2015, 186, 244–248. [Google Scholar] [CrossRef]
- Elumalai, L.K.; Rengasamy, R. Synergistic effect of seaweed manure and Bacillus sp. on growth and biochemical constituents of Vigna radiata L. J. Biofertil. Biopes. 2012, 3, 121–128. [Google Scholar] [CrossRef]
- Selvam, G.G.; Sivakumar, K. Influence of seaweed extract as an organic fertilizer on the growth and yield of Arachis hypogea L. and their elemental composition using SEM-energy dispersive spectropic analysis. Asian Pac. J. Reprod. 2014, 3, 18–22. [Google Scholar] [CrossRef]
- Vijayanand, N.; Ramya, S.S.; Rathinavel, S. Potential of liquid extracts of Sargassum wightii on growth, biochemical and yield parameters of cluster bean plant. Asian Pac. J. Reprod. 2014, 3, 150–155. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- De Gonzalo, G.; Colpa, D.I.; Habib, M.H.M.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Kellner, H.; Luis, P.; Zimdars, B.; Kiesel, B.; Buscot, F. Diversity of bacterial laccase-like multicopper oxidase genes in forest and grassland Cambisol soil samples. Soil Biol. Biochem. 2008, 40, 638–648. [Google Scholar] [CrossRef]
- Ciepiela, G.A.; Godlewska, A.; Jankowska, J. The effect of seaweed Ecklonia maxima extract and mineral nitrogen on fodder grass chemical composition. Environ. Sci. Pollut. Res. 2016, 23, 2301–2307. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Ingredient (%) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dry Matter | Crude Ash | Crude Protein | Crude Fat | TC | |||||||||||
| Panda | Kora | Mean | Panda | Kora | Mean | Panda | Kora | Mean | Panda | Kora | Mean | Panda | Kora | Mean | |
| Seaweed E. maxima extract | 91.27 d,e | 91.69 f | 91.48 e | 5.37 b,c,d | 5.74 d,e,f | 5.55 b,c | 22.5 f | 22.65 f,g | 22.57 b | 3.82 a,b,c | 3.62 a | 3.72 a,b,c | 38.93 c,d,e | 38.35 c,d,e | 38.64 a |
| Nitrophenols | 90.33 a | 91.60 f | 90.96 b,c | 5.45 c,d,e | 5.47 c,d,e | 5.46 b,c | 23.06 g | 21.99 d,e | 22.52 a,b | 3.79 a,b,c | 3.71 a,b | 3.75 a,b,c | 37.59 b | 38.8 c,d,e | 38.19 a |
| P. oligandrum | 90.48 a | 91.50 e,f | 90.99 b,c | 5.47 c,d,e | 4.8 a | 5.13 a | 23.36 g | 21.09 a,b | 22.22 a | 4.06 c | 3.57 a | 3.82 a,b,c | 36.74 a | 40.99 i | 38.86 a |
| Bacillus subtilis | 90.60 a,b | 91.11 c,d | 90.85 a,b | 5.26 b,c | 5.54 c,d,e | 5.40 b | 22.25 e,f | 22.57 f,g | 22.41 a,b | 3.94 b,c | 3.55 a | 3.75 a,b,c | 41.01 i | 39.66 g,h | 40.33 b |
| Seaweed E. maxima extract + P. oligandrum | 91.06 c,d | 91.05 c,d | 91.05 b,c | 5.48 c,d,e | 5.52 c,d,e | 5.50 b,c | 23.27 g | 21.27 b | 22.27 a,b | 4.05 c | 3.67 a,b | 3.86 c | 38.75 c,d,e | 38.88 c,d,e | 38.81 a |
| Nitrophenols + P. oligandrum | 91.13 c,d | 91.17 c,d | 91.15 c,d | 5.82 e,f | 5.56 c,d,e | 5.68 c | 22.98 g | 23.4 g | 23.19 c | 3.93 b,c | 3.55 a | 3.74 a,b,c | 38.16 c | 38.57 c,d,e | 38.36 a |
| Seaweed E. maxima extract + B. subtilis | 91.68 f | 90.99 c,d | 91.33 d,e | 5.10 a,b | 5.89 f | 5.50 b,c | 22.5 f | 22.5 f | 22.50 a,b | 3.72 a,b | 3.51 a | 3.61 a | 40.14 h | 40.67 i | 40.40 b |
| Nitrophenols + B. subtilis | 90.46 a | 91.05 c,d | 90.76 a | 5.51 c,d,e | 5.07 a,b | 5.29 a,b | 22.67 f,g | 20.81 a | 21.74 a | 4.04 c | 3.62 a | 3.83 b,c | 39.14 d,e,f | 41.74 j | 40.44 b |
| Seaweed E. maxima extract + nitrophenols | 91.61 f | 90.93 c,d | 91.27 d | 4.87 a | 5.95 f | 5.41 b | 21.64 c,d | 22.33 e,f | 21.99 a | 4.05 c | 3.56 a | 3.80 a,bc | 40.01 h | 38.58 c,d,e | 39.29 a |
| Control | 91.26 d,e | 90.8 b,c | 91.03 b,c | 5.24 b,c | 5.55 c,d,e | 5.39 b | 21.65 c,d | 22.18 e,f | 21.92 a | 3.72 a,b | 3.55 a | 3.63 a,b | 39.15 d,e,f,g | 39.58 f,g,h | 39.36 a |
| Mean | 90.98 a | 91.19 b | - | 5.35 a | 5.51 b | - | 22.59 b | 22.08 a | - | 3.91 b | 3.59 a | - | 39.0 a | 39.6 b | - |
| Treatment | Ingredient (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Crude Fiber | NDF | ADF | |||||||
| Panda | Kora | Mean | Panda | Kora | Mean | Panda | Kora | Mean | |
| Seaweed Ecklonia maxima extract | 20.88 h | 20.16 f | 20.52 e,f | 40.44 i | 35.71 d | 38.07 e | 31.16 g | 27.91 b,c,e | 29.54 c |
| Nitrophenols | 20.27 f | 19.62 d,e | 19.95 c | 38.51 f,g | 35.82 d | 37.16 d | 30.65 f,g | 28.2 c,d,e | 29.42 c |
| Pythium oligandrum | 19.67 d,e | 20.93 h | 20.30 d,e | 35.90 d | 37.56 e,f | 36.73 d | 27.83 b,c,e | 28.66 d,e,f | 28.24 b |
| Bacillus subtilis | 20.21 f | 17.11 a | 18.65 a | 38.25 f | 32.61 b | 35.43 c | 28.47 d,e | 25.00 a | 26.74 a |
| Seaweed Ecklonia maxima extract + Pythium oligandrum | 19.38 c,d | 21.51 i | 20.45 e,f | 37.73 e,f | 36.89 e | 37.31 d | 29.24 d,e,f,g | 28.69 d,e,f | 28.97 b,c |
| Nitrophenols + Pythium oligandrum | 20.26 f | 19.91 e,f | 20.08 c,d | 40.00 h,i | 36.91 e | 38.45 e | 30.53 f,g | 29.73 d,e,f,g | 30.13 c |
| Seaweed Ecklonia maxima extract + Bacillus subtilis | 19.01 c | 18.29 b | 18.6 a | 34.50 c | 31.00 a | 32.75 a | 26.04 a | 25.38 a | 25.71 a |
| Nitrophenols + Bacillus subtilis | 19.29 c,d | 18.99 c | 19.14 b | 35.94 d | 33.24 b | 34.59 b | 26.38 a,b | 26.69 a,b,c | 26.54 a |
| Seaweed Ecklonia maxima extract + nitrophenols | 20.14 f | 20.49 f | 20.31 d,e | 39.19 g,h | 37.15 e | 38.17 e | 29.35 d,e,f,g | 30.13 e,f,g | 29.74 c |
| Control | 21.02 h | 20.37 f | 20.69 f | 39.64 h,i | 37.9 e,f | 38.77 e | 30.15 e,f,g | 29.24 d,e,f,g | 29.69 c |
| Mean | 20.0 b | 19.7 a | - | 38.01 b | 35.48 a | - | 28.98 b | 27.96 a | - |
| Treatment | Ingredient (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| ADL | HCEL | CEL | |||||||
| Panda | Kora | Mean | Panda | Kora | Mean | Panda | Kora | Mean | |
| Seaweed Ecklonia maxima extract | 12.10 h | 10.43 e,f | 11.27 e | 9.28 d,e | 7.80 b,c,d,e | 8.54 b | 19.33 c,d | 17.48 a,b,c,d | 18.41 c,d |
| Nitrophenols | 11.44 g | 10.03 c,d,e | 10.74 d | 7.86 b,c,d,e | 7.62 b,c,d,e | 7.74 a,b | 19.21 c,d | 18.17 a,b,c,d | 18.69 c,d |
| Pythium oligandrum | 9.53 a,b,c | 10.74 f | 10.13 c | 8.08 b,c,d,e | 8.91 c,d,e | 8.49 b | 18.3 a,b,c,d | 17.92 a,b,c,d | 18.11 b,c |
| Bacillus subtilis | 9.48 a,b,c | 8.86 a | 9.17 a | 9.79 e | 7.60 b,c,d,e | 8.69 b | 18.99 c,d | 16.15 a | 17.57 b,c |
| Seaweed Ecklonia maxima extract + Pythium oligandrum | 9.68 b,c,d | 10.91 f | 10.29 c | 8.49 b,c,d,e | 8.20 b,c,d,e | 8.34 b | 19.57 d | 17.79 a,b,c,d | 18.68 c,d |
| Nitrophenols + Pythium oligandrum | 10.75 f | 10.27 d,e,f | 10.51 c | 9.48 e | 7.18 a,b,c,d | 8.33 b | 19.78 d | 19.46 d | 19.62 d |
| Seaweed Ecklonia maxima extract + Bacillus subtilis | 9.13 a,b | 9.06 a,b | 9.09 a | 8.46 b,c,d,e | 5.63 a | 7.04 a | 16.92 a,b,c | 16.32 a | 16.62 a |
| Nitrophenols + Bacillus subtilis | 9.94 c,d,e | 9.26 a,b | 9.60 b | 9.56 e | 6.55 a,b | 8.05 a,b | 16.45 a,b | 17.43 a,b,c,d | 16.94 a,b |
| Seaweed Ecklonia maxima extract + nitrophenols | 11.49 g | 11.82 g,h | 11.65 f | 9.84 e | 7.02 a,b,c | 8.43 b | 17.86 a,b,c,d | 18.31 a,b,c,d | 18.08 b,c |
| Control | 12.13 h | 10.4 e,f | 11.26 e | 9.49 e | 8.67 b,c,d,e | 9.08 b | 18.03 a,b,c,d | 18.84 b,c,d | 18.43 c,d |
| Mean | 10.56 b | 10.18 a | - | 9.03 b | 7.52 a | - | 18.44 b | 17.78 a | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witkowicz, R.; Biel, W.; Chłopicka, J.; Galanty, A.; Gleń-Karolczyk, K.; Skrzypek, E.; Krupa, M. Biostimulants and Microorganisms Boost the Nutritional Composition of Buckwheat (Fagopyrum esculentum Moench) Sprouts. Agronomy 2019, 9, 469. https://doi.org/10.3390/agronomy9080469
Witkowicz R, Biel W, Chłopicka J, Galanty A, Gleń-Karolczyk K, Skrzypek E, Krupa M. Biostimulants and Microorganisms Boost the Nutritional Composition of Buckwheat (Fagopyrum esculentum Moench) Sprouts. Agronomy. 2019; 9(8):469. https://doi.org/10.3390/agronomy9080469
Chicago/Turabian StyleWitkowicz, Robert, Wioletta Biel, Joanna Chłopicka, Agnieszka Galanty, Katarzyna Gleń-Karolczyk, Edyta Skrzypek, and Mateusz Krupa. 2019. "Biostimulants and Microorganisms Boost the Nutritional Composition of Buckwheat (Fagopyrum esculentum Moench) Sprouts" Agronomy 9, no. 8: 469. https://doi.org/10.3390/agronomy9080469