1. Introduction
A diverse range of factors influence the availability of macronutrients in soils of native grasslands, often with a large degree of interaction among factors. The availability of macronutrients, and the forms of macronutrients, within soils of native grasslands generally reflects the nature of parent materials from which the soils evolved, and is further affected by plant communities, climate, position within landscapes, and management [
1,
2,
3]. These factors result in variable spatial and temporal distribution patterns for plant-available macronutrients in soils of ecosystems dominated by perennial grasses [
1,
3]. Hence, the importance of macronutrients to landscape organization and biological function of grasslands [
4,
5,
6].
Cattle can affect the distribution of plant-available macronutrients in grassland ecosystems by grazing plant biomass and; removing macronutrients from paddocks in weight gain (via consumption, rumination and conversion of macronutrients to animal mass), and by redistributing unincorporated macronutrients in excreta within paddocks in heterogeneous patterns [
3,
7,
8,
9]. The behavior of cattle and their preferential use of different zones of the landscape within paddocks are factors that can drive redistribution of macronutrients within paddocks [
1,
10,
11]. The redistribution of macronutrients through excreta can be non-uniform, resulting in high concentrations of more labile forms in localized areas of paddocks [
12,
13]. There have been reported increases in amounts of labile forms of N, P, K, and S in areas near watering facilities, corners, and other structures within paddocks of grazed rangeland and perennial grasslands [
9,
14,
15].
Some systems of stocking are thought to be capable of altering how cattle redistribute more labile forms of macronutrients, primarily N, P, K, and S within grazed landscapes [
3,
15,
16]. This impact is of particular interest in cases where perennial grasslands are managed by different methods of rotational stocking, which may prevent high and disproportionate loadings in certain areas of paddocks [
15]. However, the natural distribution of macronutrients, and physical properties, of soils in production-scale paddocks tend to be variable without overlaying the effects of applied grazing systems [
1].
Given the naturally variable landscapes within production-scale paddocks, an important question is whether stocking methods can influence distributions of plant-available macronutrients within managed rangeland landscapes. The objective of this study was to apply a simple test for differences in levels of plant-available macronutrients in soils at two contrasting locations within paddocks (near water sources, near paddock centers) of southern tallgrass prairie managed under different methods of stocking. The working hypotheses were, that there would be no difference in the level of availability of eight macronutrients at these locations within paddocks, or differences in amounts at different times of growing seasons, in response to four stocking methods.
2. Materials and Methods
This study was conducted at the USDA-ARS Grazinglands Research Laboratory (35°33′29″ N, 98°1′50″ W; 435 m elevation) in central Oklahoma, USA. The entire site was located in a rolling upland landscape, with a range of features present. Included were local easterly and westerly-facing slopes (3 to 6%) on riser positions, and toe and tread slope positions (0 to 2% slopes), bordering the risers [
17]. The long-term average (LTA; 1977 to 2012) precipitation [±1 standard deviation [SD) during calendar years was 941(±174) mm. The majority of precipitation received annually had a bimodal distribution with maxima in April through June (334 ± 58 mm), and September through October (175 ± 54 mm). Long-term monthly minimum and maximum temperatures (±1 s.d.) were recorded in January [2.8(± 2.7) °C; −4.0 to 8.6 °C] and July [28.1(± 1.5) °C; 25.6 to 32.4 °C].
The study was located within production-scale paddocks on 346 ha of native grassland defined as southern tallgrass prairie. The grasslands within the paddocks are remnants of the original southern tallgrass ecosystem that existed in central Oklahoma prior to settlement, and were never cultivated or replanted to native grasses following cultivation. The plant community was identified as a Loamy Prairie ecological site ([
17], ecological site number 080AY056OK). Dominant grasses were the perennials big bluestem (
Andropogon gerardii), little bluestem (
Schizachyrium scoparium), and Indiangrass (
Sorghastrum nutans). These three indigenous species generate 60% to 80% of the total above-ground biomass produced annually within the area that was included in the study [
18]. The area was historically (1977 to 2009) used to support cow herds that were managed to generate calves of different breed-types and crosses for research projects on production systems applied to yearling stocker cattle [
19,
20]. Stocking methods that were applied during the historical period were changed regularly to meet management requirements for yearling cattle used in research projects. Included were periods of management under either, or both, continuous and rotational stocking applied to portions of the area during growing seasons (April to September) at low densities (2.0 to 3.0 ha cow/calf pair
−1 growing season
−1).
A range of soils exists within the area [
17,
21], based on position within the landscape. Norge series silt loams (fine-silty, mixed, active thermic Udic Paleustolls), situated on riser (mid-slope) positions of the landscape were the most common soil [
17]. The Norge series has three sub-types based on the location along risers, and degree of slope. Renfrow or Kirkland silt loams (fine, mixed-superactive, thermic Udertic Paleustolls) on tread positions (summit locations) and Port silt loams (fine-silty, mixed-superactive, thermic Cumulic Haplustolls) at toe positions bounded the Norge series within the study site. Up to six related soils are present as inclusions within the boundaries of each of these soils [
17]. The surface soils of these series have near-neutral but variable pH levels (6.7 ± 0.6), a water-holding capacity of 3 ± 1 mm cm
−1 soil, and permeability rates of 33 ± 17 mm h
−1 [
17]. These soils evolved from parent material, defined as Permian-aged Dog Creek shale, comprised of reddish-brown shale with thin inter-beds of siltstone and sandstone [
21].
Two replicate 61(± 2) ha paddocks, managed by continuous yearlong stocking, were included in the study as controls. Additional sets of sub-paddocks that were components of two replicate 80(± 2) ha paddocks (10 sub-paddocks each) of rotationally-stocked rangeland were also included. The design and organization of paddocks and sub-paddocks used in this study were variable in size, shape, and dimensions. Two sets of additional small paddocks (0.4 and 0.2 ha) were also established within the areas of the continuous paddocks, with one set each at toe and tread positions. These small units were used to mimic the application of high-density short-duration rotational stocking (HDRG), known as mob stocking [
22], and their impacts on plant communities and soil properties.
Grazing pressure was achieved by herds of cow–calf pairs (~500 kg cows and up to 249 kg calves at weaning) as animal units (AU) that were assigned to replicate experimental units receiving continuous and rotational stocking. Daily forage allotments per AU were 14 kg d
−1, or ~5.1 Mg AU
−1 yr
−1. Herd sizes in the continuous paddocks and sets of rotational-stocked sub-paddocks were 18(± 3.4) and 26(± 2.5) AU yr
−1 respectively, during 2009–2015. The total grazing pressure applied to the continuous and rotational paddocks averaged roughly 108 and 119 animal unit days (AUD) ha
−1 yr
−1. The cow herds assigned to continuous-stocked paddocks were used to apply the high-density short-duration stocking treatments, which are also known as mob stocking. These paddocks were grazed once per year for 24-h, with the timing of grazing varied annually. Cattle from the two continuous-stocked paddocks were randomly applied to one of the HDRG treatments on each replicate for 24 h, returned to the continuous paddocks for 48 h, and then applied to the remaining HDRG treatment for 24 h. The mob-stocked paddocks served as examples of two levels of high-density rapid-rotation stocking [18(± 3.4 head) for 24 h on 0.8 ha (HDRG-1×) and 0.4 ha (HDRG-2×) units, which applied 23 and 45 AUD ha
−1 yr
−1 of grazing pressure] that have garnered producer interest in Oklahoma, USA, in recent years. Cow–calf pairs assigned to the rotational-stocked paddocks grazed sub-paddocks in 7 to 10-day grazing bouts, up to four times per year, with the number and timing of bouts varied annually. Therefore, the applied rotational and HDRG systems were representative of different adaptive forms of rotational stocking [
22]. All paddocks in this study were managed under their assigned stocking methods from 2009 through 2015.
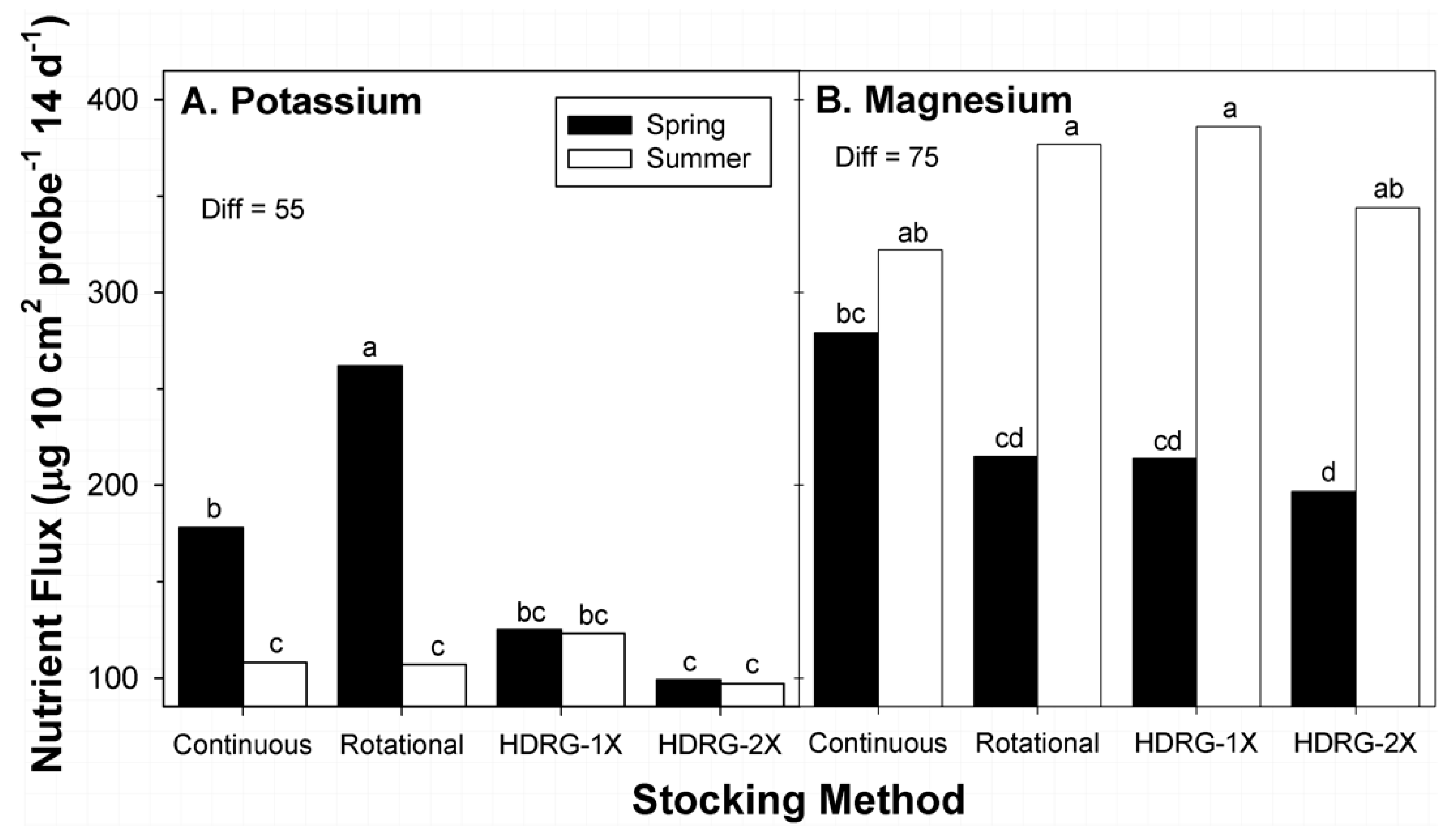
Data were collected at two times during the 2015 growing season: mid-March at the initiation of growth by native grasses, and early August, when peak living biomass occurs for southern tallgrass prairie during late summer [
18]. Sub-paddocks within the rotational stocked units that were sampled in March 2015 were last grazed in mid-August 2014, while the mob-stocked paddocks were previously grazed in early September 2014. The ~7-month delay in sampling of the rotational and mob-stocked paddocks in March 2015 occurred because there was only one set of mob-stocked paddocks per replicate, and their planned timing of grazing for 2015 was late summer. Sampling of the rotational sub-paddocks and mob-stocked units in August 2015 occurred ~7 days after grazing by cattle. This coordination of sampling within the rotational and HDRG-stocked paddocks allowed for more effective comparisons among different forms of rotational stocking.
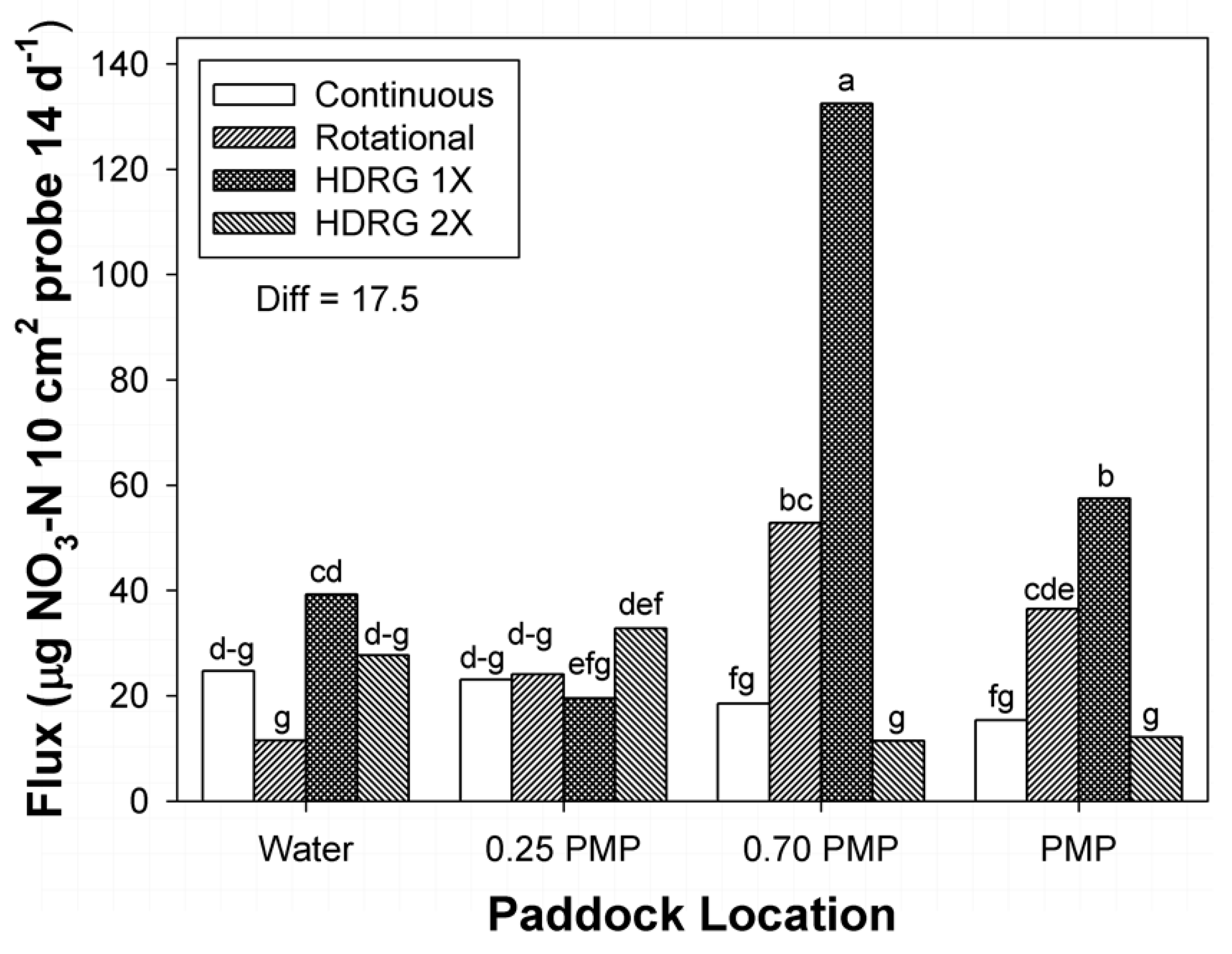
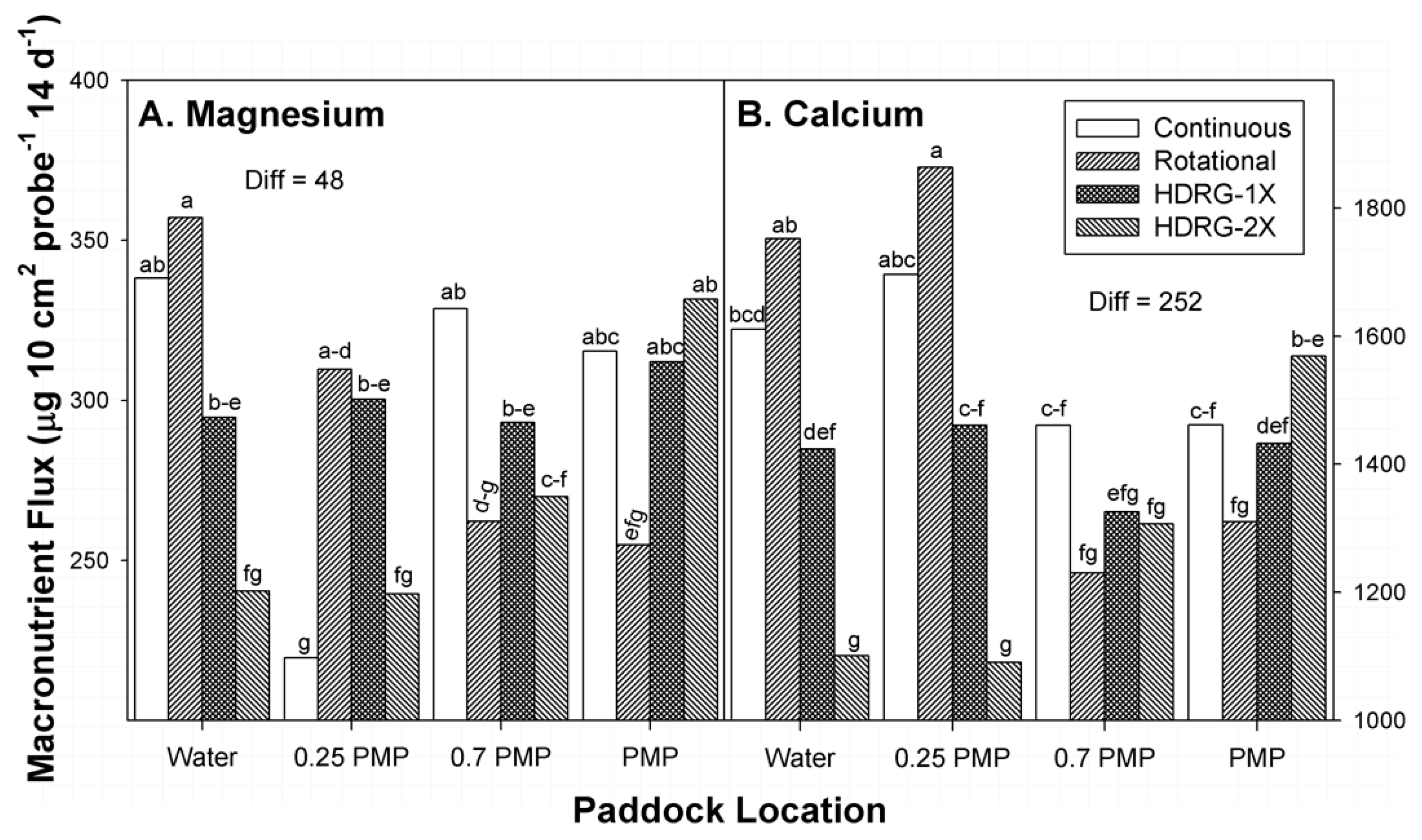
At each sampling date, the availability of macronutrients within the 0 to 7.5 cm and 7.5 to 15 cm depth increments of soil were determined at four locations within the paddocks at four locations within two replicate paddocks (or sub-paddocks) per stocking method. Locations were oriented along straight lines from water sources to centers of units. All water sources within paddocks or sub-paddocks of this study were located near paddock corners. Locations were: (1) 1.0 to 2.0 m distance from water tanks, (2) 25% of the distance between tanks and the mid-point of paddocks (0.25 PMP), (3) 70% of the distance between water and the mid-point of paddocks (0.7 PMP), and (4) at the mid-points of paddocks (PMP). The distances of 0.25 PMP, 0.7 PMP, and PMP locations from water within paddocks varied with size and shape of the individual paddocks and sub-paddocks that were sampled (
Table 1). Therefore, all locations represented random locations within each paddock or sub-paddock due to pasture size, shape, and dimensions. These locations represented areas within rangeland paddocks with two divergent capacities to attract cattle: (1) areas where animals are guaranteed to interact with the soil and plant community (locations closer to water), and (2) areas at the furthest point from water and other attractant features (fence lines, corners) within paddocks [
23].
Availability of macronutrients within soils was determined with Plant Root Simulator TM probes (Western Ag Innovations Inc., Saskatoon, SK, Canada). The probes consist of paired sets of anion and cation exchange membranes encased in plastic housings. At each sampled location within paddocks, two sets of probes with anion and cation membranes were co-located within 30 cm areas around paddock locations, at each soil depth. The probes were buried in situ during the March sampling for a 14-day incubation period. Soil moisture approximated field capacity in March [22(±3)% volumetric water] due to precipitation events that occurred prior to incubation periods.
Soil moisture at the time of the August sampling approximated permanent wilt-point [12(±1)% volumetric water] due to a prolonged dry period during summer. Therefore, replicate (n = 4) 5.38 cm diameter soil cores for each location and soil depth were collected and removed intact to a laboratory. Collected cores were wetted to field capacity with deionized water, dissected into two sections along the long axis of cores, and probes with anion and cation membranes were each sandwiched within two separate cores for incubation. The probes were removed following incubation periods for both sampling dates, lightly washed with deionized water to remove soil, packaged in groups by paddocks, refrigerated, and sent to the probe manufacturer for analyses. The manufacturer used colorimetric analyses via automated flow injection to determine NO3-N and NH4-N, and the other macronutrients (ions) were measured using inductively-coupled plasma spectrometry.
Two additional sets of soil cores were collected at times of sampling to define physical properties of soils, within a 50 cm diameter area as the probes, at the different sampled locations. Moist bulk densities of one set of samples (
n = 8 soil depth
−1 stocking method
−1) were defined [
24], followed by analyses of particle fractions by hydrometer methods in a sodium hexa-metasulfate solution [
25]. A second set of soil samples (
n = 8 soil depth
−1 stocking method
−1) were used to define estimates of soil organic matter based on low temperature [380 °C for 16 h on oven-dried (105 °C for 24 h) samples] loss on ignition [
26].
Exploratory analyses were applied to amounts of macronutrients absorbed by the probes (
Table 2) to determine whether data transformations were required for statistical tests [
27]. We applied natural logarithm (Ln) transforms, as required, to improve the cumulative distribution functions of the populations of observations so they more closely fit a normal distribution. Transformed levels of flux of total mineral N, NO
3, NH
4, P, S, and K, and the raw values for Ca and Mg were analyzed in SAS 9.3 (SAS Institute, Cary, NC) by longitudinal (repeated) measures analyses [
28] within mixed-models (PROC MIXED). Grazing regime, soil depth, and time of growing season were main effects in analyses, while sampled location within paddock was the longitudinal effect. Particle fractions, bulk density, organic matter, and distances between water sources and sampled locations were tested as covariates, to improve function and tests of statistical models. However, all physical attributes of soil were reported as non-significant for effects on analyses (0.23 <
P < 0.88). Distances between water and sampled locations were retained as a covariate, as they differed among individual paddocks assigned to stocking methods, and tests indicated there was some influence on the function of statistical models [
29]. Therefore, means and standard deviations (SD) of particle fractions, bulk density, and organic matter are reported to provide some estimates of soil properties at the sampled locations within paddocks.
Compound symmetry (CS) variance–covariance matrices [
28] were used to account for covariance and autocorrelation among locations within paddocks as there were not enough degrees of freedom (d.f.) to utilize more complex matrix structures [
29]. All analyses were restricted to 2-way interactions among main factors and the longitudinal factor due to a lack of d.f. required to test higher-order interactions [
27]. The PDIFF procedure of SAS [
29] was used to test for differences among means of significant main and interaction effects. Reported means were back-transformed to original scales of flux [
27]. The level of significance of statistical tests was set at
P = 0.10.






{kind=link}
{kind=link}
{kind=link}