Feed Value of Barn-Dried Hays from Permanent Grassland: A Comparison with Fresh Forage

Abstract
:1. Introduction
2. Materials and Methods
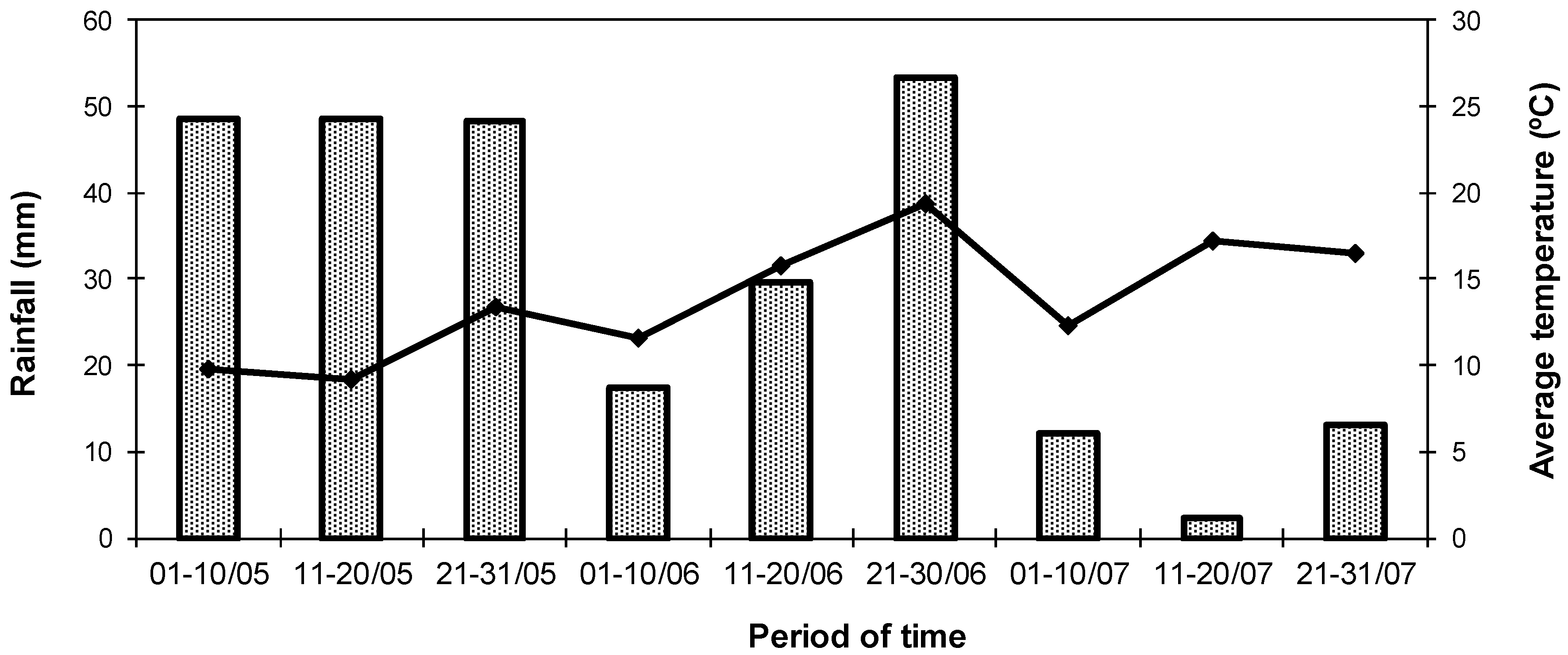
2.1. Forages and Climatic Conditions
2.2. In Vivo Digestibility Trials
2.3. Samples
2.4. Analyses
2.5. Calculations
2.6. Statistical Analysis
3. Results
3.1. Botanical Composition and Phenological Stage
3.2. Chemical Composition
3.3. Digestibility and Intake
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bruinenberg, M.H.; Valk, H.; Korevaar, H.; Struik, P.C. Factors affecting digestibility of temperate forages from seminatural grasslands: A review. Grass Forage Sci. 2002, 57, 292–301. [Google Scholar] [CrossRef]
- Andueza, D.; Cruz, P.; Farruggia, A.; Baumont, R.; Picard, F.; Michalet-Doreau, B. Nutritive value of two meadows and relationships with some vegetation traits. Grass Forage Sci. 2010, 65, 325–334. [Google Scholar] [CrossRef]
- Andueza, D.; Picard, F.; Jestin, M.; Aufrere, J. The effect of feeding animals ad libitum vs. at maintenance level on the in vivo digestibility of mown herbage from two permanent grasslands of different botanical composition. Grass Forage Sci. 2013, 68, 418–426. [Google Scholar] [CrossRef]
- Rotz, C.A.; Muck, R.E. Changes in forage quality during harvest and storage. In Forage Quality, Evaluation, and Utilization; Fahey, G.C., Collins, M., Mertens, D.R., Moser, L.E., Eds.; American Society of Agronomy, Inc.; Crop Science Society of America, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; pp. 828–868. [Google Scholar]
- Pasha, T.N.; Prigge, E.C.; Russell, R.W.; Bryan, W.B. Influence of moisture-content of forage diets on intake and digestion by sheep. J. Anim. Sci. 1994, 72, 2455–2463. [Google Scholar] [CrossRef]
- Archimede, H.; Poncet, C.; Boval, M.; Nipeau, F.; Philibert, L.; Xande, A.; Aumont, G. Comparison of fresh and dried Digitaria decumbens grass intake and digestion by Black-belly rams. J. Agric. Sci. 1999, 133, 235–240. [Google Scholar] [CrossRef]
- Dulphy, J.P.; Rouel, J. Effect of wilting on changes in the voluntary feed-intake in cattle compared to sheep. Ann. Zootech. 1988, 37, 31–41. [Google Scholar] [CrossRef]
- Demarquilly, C. Effect of low temperature-dehydration on forage feed-value. Ann. Zootech. 1970, 19, 45–51. [Google Scholar] [CrossRef]
- Delaby, L.; Peccatte, J.R. Feeding value of ventilated hay from multi-specific pastures. Fourrages 2008, 195, 354–356. [Google Scholar]
- Andueza, D.; Delgado, I.; Muñoz, F. Effect of lucerne preservation method on the feed value of forage. J. Sci. Food Agric. 2009, 89, 1991–1996. [Google Scholar] [CrossRef]
- Demarquilly, C.; Chenost, M.; Giger, S. Pertes fécales et digestibilité des aliments et des rations. In Nutrition des Ruminants Domestiques. Ingestion et Digestion; Jarrige, R., Ruckebusch, Y., Demarquilly, C., Farce, M.H., Journet, M., Eds.; INRA Éditions: Paris, France, 1995; pp. 601–647. [Google Scholar]
- Moore, K.J.; Moser, L.E.; Vogel, K.P.; Waller, S.S.; Johnson, B.E.; Pedersen, J.F. Describing and quantifying growth-stages of perennial forage grasses. Agron. J. 1991, 83, 1073–1077. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990; p. 1298. [Google Scholar]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber neutral detergent fiber, and nonstarch poysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B. Systems of Analysis for Evaluating Fibrous Feeds; Pidgen, W.J., Balch, C.C., Graham, M., Eds.; IDRC No 134; International Development Research Centre: Ottawa, ON, USA, 1980; pp. 49–60. [Google Scholar]
- Otto, S.; Masin, R.; Chiste, G.; Zanin, G. Modelling the correlation between plant phenology and weed emergence for improving weed control. Weed Res. 2007, 47, 488–498. [Google Scholar] [CrossRef]
- SAS. SAS/STAT UsersGuide, Version 6.12; Statistical Analysis System Institute: Cary, NC, USA, 1998. [Google Scholar]
- Andueza, D.; Rodrigues, A.M.; Picard, F.; Rossignol, N.; Baumont, R.; Cecato, U.; Farruggia, A. Relationships between botanical composition, yield and forage quality of permanent grasslands over the first growth cycle. Grass Forage Sci. 2016, 71, 366–378. [Google Scholar] [CrossRef]
- Todorova, P.A.; Kirilov, A.P. Changes in the permanent grassland composition and feeding value during the growing season. Grassl. Sci. Eur. 2002, 7, 170–171. [Google Scholar]
- Rees, D.V.H. A discussion of sources of dry-matter loss during the process of haymaking. J. Agric. Eng. Res. 1982, 27, 469–479. [Google Scholar] [CrossRef]
- Dulphy, J.P. Fenaison: Pertes en cours de récolte et de conservation. In Les Fourrages Secs: Récolte, Traitement, Utilisation; Demarquilly, C., Ed.; INRA: Paris, France, 1987; pp. 103–124. [Google Scholar]
- McGechan, M.B. A review of losses arising during conservation of grass forage. 1. Field losses. J. Agric. Eng. Res. 1989, 44, 1–21. [Google Scholar] [CrossRef]
- Buxton, D.R. Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Anim. Feed Sci. Technol. 1996, 59, 37–49. [Google Scholar] [CrossRef]
- Montossi, F.; Liu, F.; Hodgson, J.; Morris, S.; Barry, T.; Risso, D. Influence of low-level condensed tannins concentrations in temperate forages on sheep performance. In Proceedings of the XVIIIth International Grassland Congress, Saskatoon, SK, Canada, 8–19 June 1997; pp. 8.1–8.2. [Google Scholar]
- Lees, G.L.; Hinks, C.F.; Suttill, N.H. Effect of high-temperature on condensed tannin accumulation in leaf tissues of big trefoil (Lotus Uliginosus Schkuhr). J. Sci. Food Agric. 1994, 65, 415–421. [Google Scholar] [CrossRef]
- Theodoridou, K.; Aufrere, J.; Andueza, D.; Le Morvan, A.; Picard, F.; Stringano, E.; Pourrat, J.; Mueller-Harvey, I.; Baumont, R. Effect of plant development during first and second growth cycle on chemical composition, condensed tannins and nutritive value of three sainfoin (Onobrychis viciifolia) varieties and lucerne. Grass Forage Sci. 2011, 66, 402–414. [Google Scholar] [CrossRef]
- Frutos, P.; Hervas, G.; Giraldez, F.J.; Mantecon, A.R. Review. Tannins and ruminant nutrition. Span. J. Agric. Res. 2004, 2, 191–202. [Google Scholar] [CrossRef]
- Scharenberg, A.; Arrigo, Y.; Gutzwiller, A.; Wyss, U.; Hess, H.D.; Kreuzer, M.; Dohme, F. Effect of feeding dehydrated and ensiled tanniferous sainfoin (Onobrychis viciifolia) on nitrogen and mineral digestion and metabolism of lambs. Arch. Anim. Nutr. 2007, 61, 390–405. [Google Scholar] [CrossRef]
- Michalet Doreau, B.; Gatel, F. Annual variations in voluntary food-intake of wethers. Ann. Zootech. 1988, 37, 151–158. [Google Scholar] [CrossRef]
- Estrada, J.I.C.; Delagarde, R.; Faverdin, P.; Peyraud, J.L. Dry matter intake and eating rate of grass by dairy cows is restricted by internal, but not external water. Anim. Feed Sci. Technol. 2004, 114, 59–74. [Google Scholar] [CrossRef]


{kind=link}
| Stage | Code | Description |
|---|---|---|
| Vegetative | ||
| Vegetative leaf development | 1.5 | |
| Stem Elongation | ||
| Elongation (beginning) | 2 | First node palpable/visible |
| Elongation | 2.5 | Nodes palpable/visible |
| Elongation (end) | 3 | Boot stage |
| Reproductive/Floral Development | ||
| Inflorescence emergence | 3.1 | First spikelet visible |
| Inflorescence | 3.3 | Spikelets fully emerged/peduncle not emerged |
| Inflorescence emerged | 3.5 | Inflorescence emerged/peduncle fully elongated |
| Anther emergence/anthesis | 3.8 | |
| Seed Development and Ripening | ||
| Caryopsis visible | 4 | |
| Milk | 4.1 | |
| Dough | 4.4 | |
| Endosperm hard/physiological maturity | 4.7 | |
| Endosperm dry/seed ripe | 4.9 | |
| P1 1 | P2 2 | P3 3 | |
|---|---|---|---|
| Grasses | |||
| Agrostis capillaris L. | 20.23 | 12.96 | 29.87 |
| Trisetum flavescens L. | 4.41 | 4.01 | 7.02 |
| Bromus mollis L. | 1.03 | 0.55 | 0.00 |
| Dactylis glomerata L. | 15.39 | 4.92 | 0.84 |
| Festuca rubra L. | 3.14 | 2.02 | 9.97 |
| Anthoxanthum odoratum L. | 0.19 | 0.00 | 1.61 |
| Poa pratensis L. | 6.06 | 7.37 | 8.25 |
| Holcus lanatus L. | 1.44 | 0.19 | 6.34 |
| Lolium perenne L. | 18.81 | 56.46 | 16.80 |
| Phleum pratense L. | 5.64 | 0.27 | 2.62 |
| Forbs | |||
| Cerastium fontanum Baumg. | 9.57 | 1.54 | 5.41 |
| Taraxacum officinale P.H. Wigg. | 11.64 | 2.54 | 3.16 |
| Urtica dioica L. | 0.00 | 1.01 | 0.02 |
| Sum of grasses+forbs | 97.55 | 93.84 | 91.91 |
| Others | |||
| Senescent material | 0.72 | 4.70 | 6.36 |
| MPW | 2.65 | 3.79 | 3.82 |
| Forage | Period | Significance | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FF 1 | BDF 2 | P1 3 | P2 4 | P3 5 | SEM 6 | Period | Contrast | Forage | Period × Forage | |
| CA | 80 | 79 | 82 | 71 | 84 | 10.2 | ns 10 | ns | ns | ns |
| CP | 11.9 | 12.1 | 15.6 | 10.6 | 9.9 | 0.60 | *** | L 8,* Q 9,* | ns | ns |
| NDF | 598 | 592 | 545 | 590 | 652 | 16.8 | *** | L * | ns | ns |
| ADF | 320 | 322 | 287 | 321 | 355 | 6.3 | *** | L * | ns | ns |
| ADL | 45 | 40 | 33 | 42 | 53 | 5.9 | * | L * | ns | ns |
| WSC | 104 | 92 | 99 | 105 | 90 | 11.1 | ns | ns | ns | ns |
| Fresh Forage | Barn-Dried Forage | Significance | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 1 | P2 2 | P3 3 | SEM 4 | Contrast | P1 | P2 | P3 | SEM | Contrast | F 5 | P 6 | F × P | |
| DMD | 0.64 | 0.56 | 0.41 | 0.564 | L 7,* | 0.64 | 0.55 | 0.50 | 0.013 | L * Q 8,* | † | *** | *** |
| OMD | 0.66 | 0.60 | 0.50 | 0.012 | L * | 0.66 | 0.59 | 0.52 | 0.012 | L * | ns 9 | *** | ns |
| VI | 67.07 | 50.27 | 49.75 | 4.408 | L * Q * | 80.87 | 60.96 | 53.20 | 4.408 | L * Q * | * | *** | ns |
| DOMI | 35.94 | 26.95 | 22.48 | 2.665 | L * Q * | 49.54 | 33.60 | 25.18 | 2.665 | L * Q * | ** | *** | ns |
| ST 1 | MPW | Ash | CP | NDF | ADF | ADL | WSC | DMDff | OMDff | VIff | DOMIff | DMDbdf | OMDbdf | VIbdf | DOMIbdf | Gram | Leg | for | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MPW 2 | 0.66 | ||||||||||||||||||
| Ash | 0.31 | −0.51 | |||||||||||||||||
| CP 3 | −0.87 | −0.94 | 0.19 | ||||||||||||||||
| NDF 4 | 0.99 * | 0.67 | 0.29 | −0.88 | |||||||||||||||
| ADF 5 | 0.99 † | 0.74 | 0.20 | −0.92 | 0.99 † | ||||||||||||||
| ADL 6 | 0.99 * | 0.70 | 0.26 | −0.90 | 0.99 * | 0.99 * | |||||||||||||
| WSC 7 | −0.39 | 0.44 | −0.99 † | −0.11 | −0.36 | −0.28 | −0.33 | ||||||||||||
| DMDff 8 | −0.99 * | −0.60 | −0.38 | 0.83 | −0.99 † | −0.98 | −0.99 † | 0.45 | |||||||||||
| OMDff 9 | −0.99 * | −0.63 | −0.34 | 0.86 | −0.99 * | −0.99† | −0.99 † | −0.41 | 0.99 * | ||||||||||
| VIff 10 | −0.81 | −0.97 | 0.30 | 0.99 † | −0.82 | −0.87 | −0.84 | −0.23 | 0.76 | 0.79 | |||||||||
| DOMIff 11 | −0.95 | −0.86 | −0.01 | 0.98 | −0.96 | −0.98 | −0.97 | 0.08 | 0.92 | 0.94 | 0.95 | ||||||||
| DMDbdf 12 | −0.96 | −0.84 | −0.04 | 0.97 | −0.97 | −0.98 | −0.98 | 0.12 | 0.94 | 0.95 | 0.94 | 0.99 † | |||||||
| OMDbdf 13 | −0.99 † | −0.74 | −0.20 | 0.92 | −0.99 † | −0.99 * | −0.99 * | −0.28 | 0.98 | 0.99 † | 0.87 | 0.98 | 0.99 † | ||||||
| VIbdf 14 | −0.93 | −0.88 | 0.04 | 0.99 † | −0.94 | −0.97 | −0.95 | −0.03 | 0.91 | 0.92 | 0.97 | 0.99 * | 0.99 † | 0.97 | |||||
| DOMIbdf 15 | −0.96 | −0.84 | −0.03 | 0.98 | −0.96 | −0.98 | −0.97 | 0.10 | 0.93 | 0.95 | 0.94 | 0.99 * | 0.99 * | 0.98 | 0.99 * | ||||
| Gram 16 | 0.45 | 0.97 | −0.71 | −0.83 | 0.47 | 0.55 | 0.50 | 0.65 | −0.38 | −0.42 | −0.89 | −0.70 | −0.68 | −0.55 | −0.74 | −0.69 | |||
| leg 17 | −0.05 | 0.72 | −0.96 | −0.45 | −0.02 | 0.07 | 0.01 | 0.94 | 0.12 | 0.08 | −0.55 | −0.27 | −0.23 | −0.07 | −0.31 | −0.24 | 0.87 | ||
| for 18 | −0.62 | −0.99 * | 0.54 | 0.93 | −0.64 | −0.71 | −0.67 | −0.48 | 0.56 | 0.60 | 0.96 | 0.84 | 0.81 | 0.71 | 0.86 | 0.82 | −0.98 | −0.75 | |
| sm 19 | 0.94 | 0.87 | −0.03 | −0.98 | 0.95 | 0.97 | 0.96 | −0.05 | −0.91 | −0.93 | −0.96 | −0.99 * | −0.99 * | −0.97 | −0.99 * | −0.99 * | 0.73 | 0.30 | −0.85 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andueza, D.; Picard, F.; Pradel, P.; Theodoridou, K. Feed Value of Barn-Dried Hays from Permanent Grassland: A Comparison with Fresh Forage. Agronomy 2019, 9, 273. https://doi.org/10.3390/agronomy9060273
Andueza D, Picard F, Pradel P, Theodoridou K. Feed Value of Barn-Dried Hays from Permanent Grassland: A Comparison with Fresh Forage. Agronomy. 2019; 9(6):273. https://doi.org/10.3390/agronomy9060273
Chicago/Turabian StyleAndueza, Donato, Fabienne Picard, Philippe Pradel, and Katerina Theodoridou. 2019. "Feed Value of Barn-Dried Hays from Permanent Grassland: A Comparison with Fresh Forage" Agronomy 9, no. 6: 273. https://doi.org/10.3390/agronomy9060273