Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Field Weather Condition
2.2. Field Experimental Design
2.3. Growth and Yield Determination
2.4. Plant Sampling and Leaf Tissue Nutrient Concentration Analysis
2.5. Soil Sampling and Analysis
2.5.1. Soil Available Nutrients, pH and EC Values
2.5.2. Soil Enzyme Activities
2.6. Statistical Analysis
3. Results
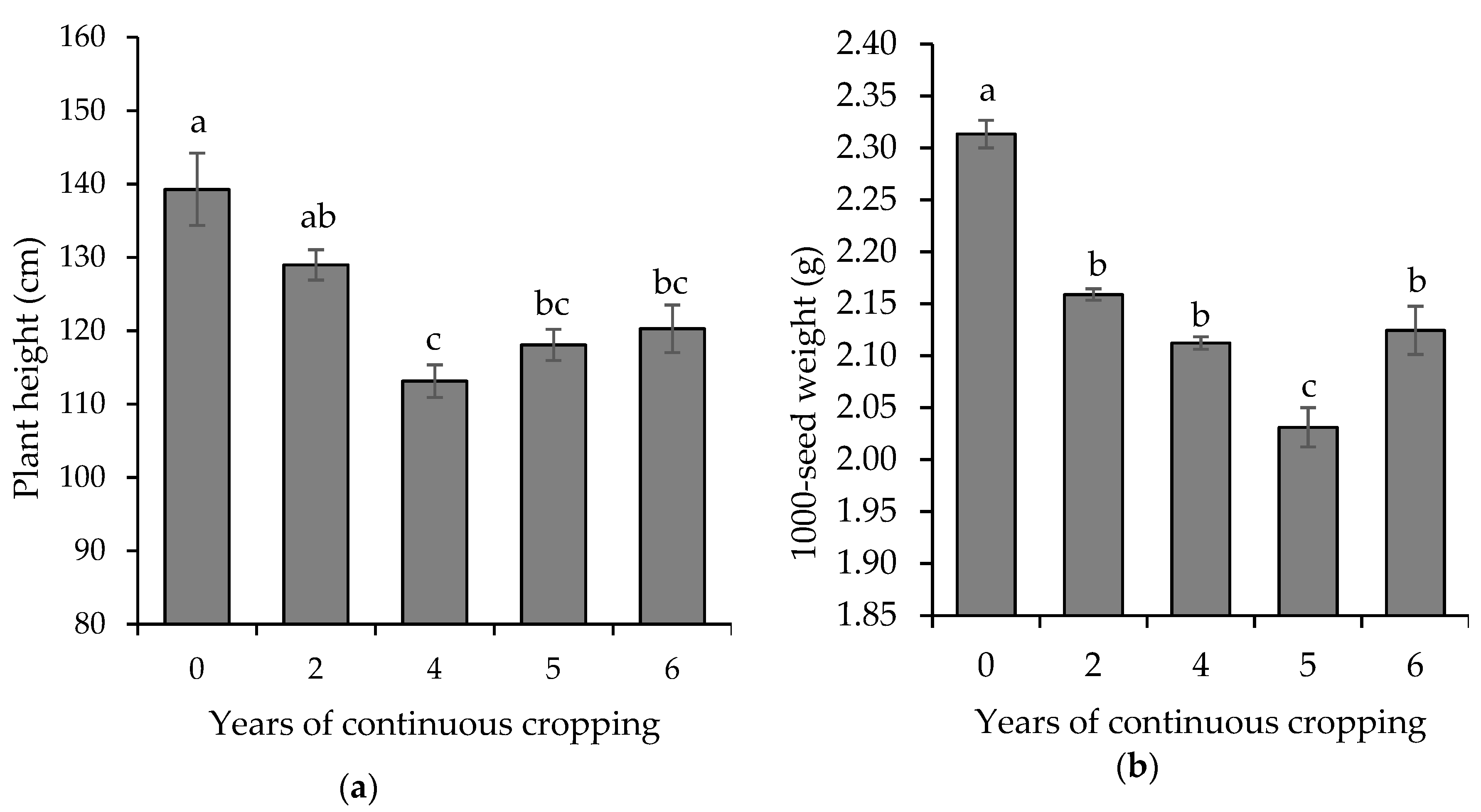
3.1. Effect of Continuous Cropping on Sesame Growth and Yield under the Different Duration of Cropping Years
3.2. Effect of Continuous Cropping on Sesame Leaf Tissue Nutrient Concentrations under the Different Duration of Cropping Years
3.3. Effect of Continuous Cropping on Soil Chemical Properties under the Different Duration of Cropping Years
3.4. Effect of Continuous Sesame Cropping on Soil Enzyme Activities under the Different Duration of Cropping Years
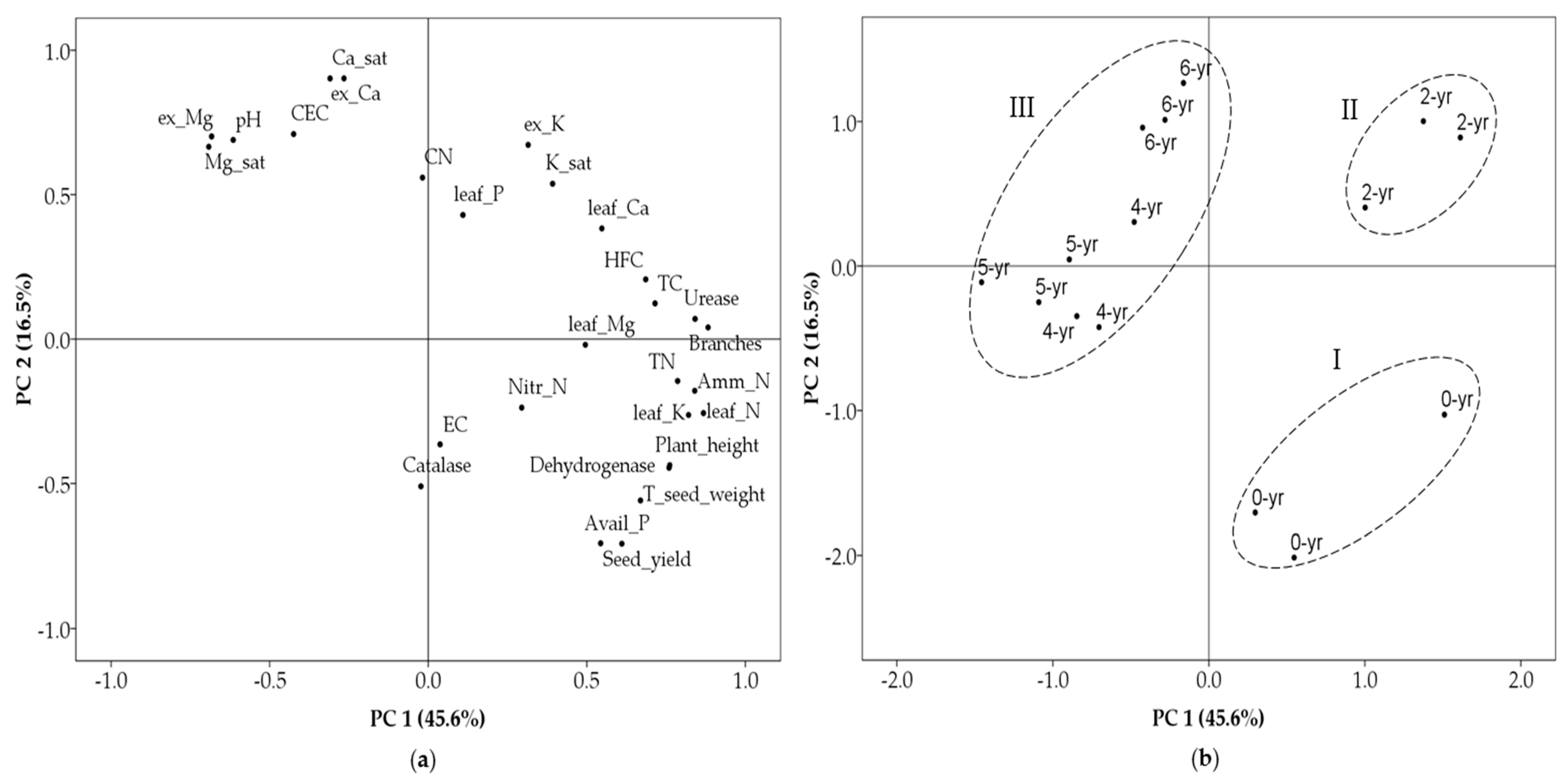
3.5. Relationship among Soil Chemical Properties, Leaf Tissue Nutrient Concentrations, Growth and Yield Decline Parameters under the Different Duration of Cropping Years
4. Discussion
4.1. Effect of Continuous Sesame Cropping on Soil Chemical Properties under the Different Duration of Cropping Years
4.2. Effect of Continuous Sesame Cropping on Leaf Tissue Nutrient Concentrations under the Different Duration of Cropping Years
4.3. Relationship between Soil Chemical Properties, Leaf Tissue Nutrient Concentrations, Growth and Yield Decline Parameters under the Different Duration of Cropping Years
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ashri, A. Sesame. In Oil Crops of the World; Robbelen, G., Downey, R.K., Ashri, A., Eds.; McGraw–Hill: New York, NY, USA, 1989; pp. 375–387. [Google Scholar]
- Yasumoto, S.; Katsuta, M. Breeding a high-lignan-content sesame cultivar in the prospect of promoting metabolic functionality. Jpn. Agric. Res. Q. 2006, 40, 123–129. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Statistical Database. 2016. Available online: http://www.fao.org/faostat/en/#data/TP (accessed on 1 March 2019).
- MAFF. FY2013 Annual Report on Food, Agriculture and Rural Areas in Japan Summary Ministry of Agriculture, Forestry and Fisheries; MAFF: Tokyo, Japan, 2014.
- Alexandratos, N.; Bruinsma, J. World agriculture towards 2030/2050: The 2012 Revision; ESA Working paper No. 12-03; FAO: Rome, Italy, 2012. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensi fi cation of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Song, L.; Xia, X. Plant-Soil Feedbacks and Soil Sickness: From Mechanisms to Application in Agriculture. J. Chem. Ecol. 2013, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Ventura, W.; Watanabe, I. Growth inhibition due to continuous cropping of dryland rice and other crops. Soil Sci. Plant Nutr. 2012, 37–41. [Google Scholar] [CrossRef]
- Gentry, L.F.; Ruffo, M.L.; Below, F.E. Identifying Factors Controlling the Continuous Corn Yield Penalty. Agron. J. 2013, 295–303. [Google Scholar] [CrossRef]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H.; Zhang, M.; Zhang, H. Soil Chemical Property Changes in Eggplant/Garlic Relay Intercropping Systems under Continuous Cropping. PLoS ONE 2014, 9, e111040. [Google Scholar] [CrossRef]
- Ye, X.H.; Zhao, Z.L.; Zhou, Q.; Zhao, H.B.; Guo, C.M.; Shi, L.L.; Guo, J.W. Malabsorption of mineral nutrients and effects of foliar fertilization on continuously cropped capsicum annum L. var. annuum. Pak. J. Bot. 2014, 46, 1781–1788. [Google Scholar]
- Zhou, X.; Wu, F. Changes in soil chemical characters and enzyme activities during continuous monocropping of cucumber (Cucumis sativus). Pakistan J. Bot. 2015, 47, 691–697. [Google Scholar]
- Zhong, S.; Mo, Y.; Guo, G.; Zeng, H.; Jin, Z. Effect of Continuous Cropping on Soil Chemical Properties and Crop Yield in Banana Plantation. J. Agric. Sci. Technol. 2014, 16, 239–250. [Google Scholar]
- Panda, R.; Patra, S.K. Depletion and Contribution Pattern of Available Potassium in Indian Coastal Soils under Intensive Cropping and Fertilization. Int. J. Pure Appl. Biosci. 2017, 5, 1144–1152. [Google Scholar] [CrossRef]
- Wyngaard, N.; Echeverria, H.E.; Sainz Rozas, H.R.; Divito, G.A. Fertilization and tillage effects on soil properties and maize yield in a Southern Pampas Argiudoll. Soil Tillage Res. 2012, 119, 22–30. [Google Scholar] [CrossRef]
- Kim, H.; Yun, B.; Choi, H.; Park, I.; Kim, S. Soil Chemical and Biological Characteristic of the Continuous Cropping Area of Garlic or Onions. Agric. Chem. Biotechnol. 2003, 46, 114–117. [Google Scholar]
- Gong, L.; He, G.; Liu, W. Long-Term Cropping Effects on Agricultural Sustainability in Alar Oasis of Xinjiang, China. Sustainability 2016, 8, 61. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Sherene, T. Role of soil enzymes in nutrient transformation: A review. Bio Bull. 2017, 3, 109–131. [Google Scholar]
- Salazar, S.; Sánchez, L.E.; Alvarez, J.; Valverde, A.; Galindo, P.; Igual, J.M.; Peix, A.; Santa-regina, I. Correlation among soil enzyme activities under different forest system management practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- Wang, Q.; Xiao, F.; He, T.; Wang, S. Responses of labile soil organic carbon and enzyme activity in mineral soils to forest conversion in the subtropics. Ann. For. Sci. 2013, 70, 579–587. [Google Scholar] [CrossRef]
- Hua, J.L.; Liu, G.R.; Huang, J.S. Effect of continuous cropping of sesame on rhizospheric microbial communities. Acta Ecol. Sin. 2012, 32, 2936–2942. [Google Scholar]
- Nam, S.Y.; Kim, I.J.; Kim, M.J.; Kang, H.J.; Yun, T.; Rho, C.W.; Min, K.B.; Lee, C.H. Effects of green manure crops and rotational cropping system on growth and yield of sesame (Sesamum indicum L.). Korean J. Plant Res. 2007, 20, 404–408, (In Korean with English abstract). [Google Scholar]
- Linh, T.B.; Sleutel, S.; Elsacker, S.V.; Guong, V.T.; Khoa, L.V.; Cornelis, W.M. Inclusion of upland crops in rice-based rotations affects chemical properties of clay soil. Soil Use Manag. 2015, 31, 313–320. [Google Scholar] [CrossRef]
- Zhou, W.; Lv, T.; Chen, Y.; Westby, A.P.; Ren, W. Soil Physicochemical and Biological Properties of Paddy-Upland Rotation: A Review. Sci. World J. 2014, 2014, 856352. [Google Scholar] [CrossRef]
- Nishida, M. Decline in Fertility of Paddy Soils Induced by Paddy Rice and Upland Soybean Rotation, and Measures against the Decline. Jpn. Agric. Res. Q. 2016, 50, 87–94. [Google Scholar] [CrossRef]
- Hattori, M.; Nagumo, Y.; Sato, T.; Fujita, Y.; Higuchi, Y.; Ohyama, T. Effect of continuous and long-term paddy-upland rotation on yield reduction of soybean in Niigata Prefecture, Japan. Jpn. J. Crop Sci. 2013, 82, 11–17, (In Japanese with English abstract). [Google Scholar] [CrossRef]
- Matsumoto, S.; Yoshikawa, M. Influence of continuous cropping on yield o black soybean (Glycine max Merr. Cv. Shintanbaguro) and soil chemical properties of soils in the field converted from paddy. Jpn. J. Crop Sci. 2010, 79, 268–274, (In Japanese with English abstract). [Google Scholar]
- Cavell, A.J. The colorimetric determination of phosphorus in plant materials. J. Sci. Food Agric. 1955, 6, 479–480. [Google Scholar] [CrossRef]
- Maynard, D.G.; Kalra, Y.P. Nitrate and Exchangeable Ammonium Nitrogen. In Soil Sampling and Methods of Analysis; Canadian Soceity of Soil Science: Boca Raton, FL, USA, 1993; pp. 25–33. [Google Scholar]
- Yamaki, A. A rapid UV absorption method for determination of nitrate in soil extracts. Jpn. J. Soil Sci. Plant Nutr. 2003, 74, 195–197. (In Japanese) [Google Scholar]
- Chapman, H.D. Cation–exchange capacity. In Methods of Soil Analysis–Chemical and Microbiological Properties; Black, C.A., Ed.; American Society of Agronomy: Madison, CT, USA, 1965; pp. 891–901. [Google Scholar]
- Ross, D.S.; Ketterings, Q. Recommended Methods for Determining Soil Cation Exchange Capacity. In Recommended Soil Testing Procedures for the Northeastern United States, Northeast Regional Bulletin No. 493, 3rd ed.; Wolf, A., McGrath, J., Eds.; Agricultural Experiment Stations of Connecticut: New Haven, CT, USA, 2011; pp. 75–86. [Google Scholar]
- Truog, E. The determination of the readily available phosphorous of soils. Agron. J. 1930, 22, 874–882. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Von Mersi, W.; Schinner, F. An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride. Biol. Fertil. Soils 1991, 11, 216–220. [Google Scholar] [CrossRef]
- Goldblith, S.A.; Proctor, B.E. Photometric determination of catalase activity. J. Biol. Chem. 1950, 187, 705–709. [Google Scholar]
- Sun, J.; Zou, L.; Li, W.; Wang, Y.; Xia, Q.; Peng, M. Soil microbial and chemical properties influenced by continuous cropping of banana. Sci. Agric. 2018, 75, 420–425. [Google Scholar] [CrossRef]
- Zhao, Q.; Xiong, W.; Xing, Y.; Sun, Y.; Lin, X.; Dong, Y. Long-Term Coffee Monoculture Alters Soil Chemical Properties and Microbial Communities. Sci. Rep. 2018, 8, 6116. [Google Scholar] [CrossRef]
- Kusumawardani, P.N.; Cheng, W.; Purwanto, B.H.; Utami, S.N.H. Changes in the soil pH, EC, available P, DOC and inorganic N after land use change from rice paddy in NorthEast Japan. J. Wetl. Environ. Manag. 2017, 5, 53–61. [Google Scholar] [CrossRef]
- Anderson, N.P.; Hart, J.M.; Sullivan, D.M.; Christensen, N.W.; Horneck, D.A.; Pirelli, G.J. Applying Lime to Raise Soil pH for Crop Production (Western Oregon); Oregon State University Extension Publication EM: Corvallis, OR, USA, 2013; p. 9057. [Google Scholar]
- Xiong, W.; Li, Z.; Liu, H.; Xue, C.; Zhang, R.; Wu, H. The Effect of Long-Term Continuous Cropping of Black Pepper on Soil Bacterial Communities as Determined by 454 Pyrosequencing. PLoS ONE 2015, 1–13. [Google Scholar] [CrossRef]
- Li, X.; Lewis, E.E.; Liu, Q.; Li, H.; Bai, C.; Wang, Y. Effects of long-term continuous cropping on soil nematode community and soil condition associated with replant problem in strawberry habitat. Sci. Rep. 2016, 6, 30466. [Google Scholar] [CrossRef]
- Liu, X.; Herbert, S.J.; Hashemi, A.M.; Zhang, X.; Ding, G. Effects of agricultural management on soil organic matter and carbon transformation—A review. Plant Soil Environ. 1994, 52, 531–543. [Google Scholar] [CrossRef]
- Reeves, D.W. The role of soil organic matter in maintaining soil quality in continuous cropping systems. Soil Tillage Res. 1997, 43, 131–167. [Google Scholar] [CrossRef]
- Nishimura, S.; Yonemura, S.; Sawamoto, T.; Shirato, Y.; Akiyama, H.; Sudo, S.; Yagi, K. Effect of land use change from paddy rice cultivation to upland crop cultivation on soil carbon budget of a cropland in Japan. Agric. Ecosyst. Environ. 2008, 125, 9–20. [Google Scholar] [CrossRef]
- Nie, L.; Peng, S.; Bouman, B.A.M.; Huang, J. Alleviating soil sickness caused by aerobic monocropping: Responses of aerobic rice to nutrient supply. Field Crops Res. 2008, 107, 129–136. [Google Scholar] [CrossRef]
- Horst, W.J.; Hardter, R.P. Rotation of maize with cowpea improves yield and nutrient use of maize compared to maize monocropping in an alfisol in the northern Guinea Savanna of Ghana. Plant Soil 1994, 160, 171–183. [Google Scholar] [CrossRef]
- Janssen, M.; Lennartz, B. Horizontal and vertical water and solute fluxes in paddy rice fields. Soil Tillage Res. 2007, 94, 133–141. [Google Scholar] [CrossRef]
- Katoh, M.; Murase, J.; Hayashi, M.; Matsuya, K. Nutrient leaching from the plow layer by water percolation and accumulation in the subsoil in an irrigated paddy field. Soil Sci. Plant Nutr. 2011, 50, 721–729. [Google Scholar] [CrossRef]
- Jones, C.; Jacobsen, J. Plant Nutrition and Soil Fertility; Nutrient Management Module No. 2; Montana State University, Extension Service: Bozeman, MT, USA, 2001. [Google Scholar]
- Bolan, N.S.; Adriano, D.C.; Curtin, D. Soil acidification and liming interactions with nutrient and heavy metal transformation and bioavailability. Adv. Agron. 2003, 78, 215–272. [Google Scholar]
- Schloter, M.; Dilly, O.; Munch, J.C. Indicators for evaluating soil quality. Agric. Ecosyst. Environ. 2003, 98, 255–262. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Zobeck, T.M.; Allen, V. Soil Microbial, Chemical and Physical Properties in Continuous Cotton and Integrated Crop–Livestock Systems. Soil Sci. Soc. Am. J. 2002, 68, 1875–1884. [Google Scholar] [CrossRef]
- Riedell, W.E.; Pikul, J.L.; Jaradat, A.A.; Schumacher, T.E. Crop Rotation and Nitrogen Input Effects on Soil Fertility, Maize Mineral Nutrition, Yield, and Seed Composition. Agron. J. 2009, 10, 870–879. [Google Scholar] [CrossRef]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils, 15th ed.; Pearson Education Ltd.: London, UK, 2016; p. 381. [Google Scholar]
- Kelley, K.W.; Long, J.H., Jr.; Todd, T.C. Long-term crop rotations affect soybean yield, seed weight, and soil chemical properties. Field Crops Res. 2003, 83, 41–50. [Google Scholar] [CrossRef]
- Dossa, K.; Diouf, D.; Wang, L.; Wei, X.; Zhang, Y.; Niang, M.; Fonceka, D.; Yu, J.; Mmadi, M.A.; Yehouessi, L.W.; et al. The Emerging Oilseed Crop Sesamum indicum Enters the “Omics” Era. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Biabani, A.R.; Pakniyat, H. Evaluation of seed yield-related characters in sesame (Sesamum indicum L.) using factor and path analysis. Pak. J. Biol. Sci. 2008, 11, 1157–1160. [Google Scholar] [CrossRef]
- Shehu, H.E. Uptake and agronomic efficiencies of nitrogen, phosphorous and potassium in sesame (Sesamum indicum L.). Am. J. Plant Nutr. Fertil. Technol. 2014, 4, 41–56. [Google Scholar]
- Shehu, H.E.; Kwari, J.D.; Sandabe, M.K. Nitrogen, phosphorous and potassium nutrition of sesame (Sesamum indicum) in Mubi, Nigeria. Res. J. Agron. 2009, 3, 32–36. [Google Scholar]
- Jadav, D.P.; Padamani, D.R.; Polara, K.B.; Parmar, K.B.; Babaria, N.B. Effect of different level of sulphur and potassium on growth, yield and yield attributes of sesame (Sesamum Indicum L.). Asian J. Soil Sci. 2010, 5, 106–108. [Google Scholar]
- Mitchell, G.A.; Bingham, F.T.; Yermanos, D.M. Growth, mineral composition and seed characteristics of sesame as affected by nitrogen, phosphorus and potassium nutrition. Soil Sci. Am. Proc. 1974, 38, 925–931. [Google Scholar] [CrossRef]
- Demmig, B.; Gimmler, H. Properties of the Isolated Intact Chloroplast at Cytoplasmic K Concentrations: I. Light-Induced Cation Uptake into Intact Chloroplasts is Driven by an Electrical Potential Difference. Plant Physiol. 1983, 73, 169–174. [Google Scholar] [CrossRef]
- Loide, V. About the effect of the contents and ratios of soil’ s available calcium, potassium and magnesium in liming of acid soils. Agron. Res. 2004, 2, 71–82. [Google Scholar]
- Kumar, P.; Sharma, M.K. Nutrient Deficiencies of Field Crops: Guide to Diagnosis and Management; CAB International: Wallingford, UK, 2013; p. 243. [Google Scholar]
- Hodges, S.C. Soil Fertility Basics; Soil Science Extension, North Carolina State University: Raleigh, NC, USA, 2010; pp. 1–75. [Google Scholar]
- Yoshimura, S.; Yoshida, R.; Maeda, M.; Nishio, W.; Akaji, K. Studies on the plastic greenhouse soil of eggplant (Solanum melongena L.) by continuous cropping (4). Potassium deficiency and magnesium excess. Bull. Osaka Agric. Res. Cent. 1976, 13, 17–24, (In Japanese with English abstract). [Google Scholar]
- Bolata, E.L.; Chaves, J.C.D.; Coffee, W. Enzymatic activity and mineralization of carbon and nitrogen in soil cultivated with coffee and green manures. Rev. Bras. Cienc. Solo 2010, 34, 1573–1583. [Google Scholar] [CrossRef]
- Wolinska, A.; Stepniewska, Z. Dehydrogenase Activity in the Soil Environment Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; Canuto, R.A., Ed.; Intech: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Langham, D.R. Gowth and Development of Sesame; SESACO Corporation: Austin, TX, USA, 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Year z | Height of First Capsule (cm) | Number of Branches/Plant | Seed Yield (kg ha−1) |
|---|---|---|---|
| 0 | 61.27 a | 2.37 ab | 1952.16 a |
| 2 | 64.40 a | 2.80 a | 920.29 b |
| 4 | 51.00 c | 1.77 c | 723.17 b |
| 5 | 56.43 b | 1.70 c | 617.32 b |
| 6 | 62.20 a | 1.90b c | 641.45 b |
| ANOVA (p-values) | *** | *** | *** |
| Year z | N% | P (%) | K (%) | Ca (%) | Mg (%) |
|---|---|---|---|---|---|
| 0 | 2.69 a | 0.80 a | 2.91 ab | 0.44 ab | 0.47 a |
| 2 | 2.68 a | 0.79 a | 3.13 a | 0.48 a | 0.48 a |
| 4 | 2.38 ab | 0.55 a | 2.21 abc | 0.40 ab | 0.43 a |
| 5 | 2.09 b | 0.83 a | 1.86 bc | 0.27 b | 0.37 a |
| 6 | 2.25 b | 1.21 a | 1.56 c | 0.56 a | 0.45 a |
| ANOVA (p-values) | ** | NS | ** | * | NS |
| 1:5 (H2O) | Available N | |||||||
|---|---|---|---|---|---|---|---|---|
| Yearz | pH | EC (dS m−1) | Total C (g kg−1) | Total N (g kg−1) | C/N ratio | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | Available P (mg kg−1) |
| 0 | 5.29 c | 0.053 a | 26.36 ab | 2.57 ab | 10.23 a | 9.71 ab | 13.10 a | 71.35 a |
| 2 | 5.64 bc | 0.052 a | 28.56 a | 2.71 a | 10.54 a | 10.91 a | 9.79 a | 49.88 b |
| 4 | 5.91 ab | 0.052 a | 24.83 ab | 2.40 bc | 10.36 a | 6.53 bc | 8.06 a | 45.17 b |
| 5 | 6.02 ab | 0.052 a | 25.86 ab | 2.41 bc | 10.71 a | 5.84 c | 9.58 a | 43.31 b |
| 6 | 6.30 a | 0.046 a | 24.50 b | 2.30 c | 10.67 a | 5.94 c | 11.28 a | 40.84 b |
| ANOVA (p-values) | *** | NS | * | *** | NS | ** | NS | *** |
| Exchangeable Cations | Base Saturation (%) | ||||||
|---|---|---|---|---|---|---|---|
| Year z | K (mg kg−1) | Ca (mg kg−1) | Mg (mg kg−1) | CEC (cmolc kg−1) | K | Ca | Mg |
| 0 | 157.42 b | 799.12 c | 193.47 c | 10.62 b | 3.80 bc | 37.63 c | 15.18 c |
| 2 | 250.47 a | 1478.36 a | 281.15 b | 11.20 ab | 5.73 a | 65.97 a | 20.91 b |
| 4 | 229.90 a | 1252.35 b | 349.60 a | 11.18 ab | 5.28 ab | 56.00 b | 26.06 a |
| 5 | 161.98 b | 1478.44 a | 356.68 a | 11.67 a | 3.56 c | 63.37 ab | 25.48 a |
| 6 | 217.69 ab | 1658.71 a | 381.09 a | 11.90 a | 4.70 abc | 69.74 a | 26.72 a |
| ANOVA (p-values) | ** | *** | *** | ** | ** | *** | *** |
| Year z | Urease (μg NH4+-N g−1 soil 2h−1) | Catalase (mL of 0.02M KMnO4 g−1 soil h−1) | Dehydrogenase (INTF μ moles g−1 soil 2 h−1) |
|---|---|---|---|
| 0 | 97.17 ab | 6.311 ab | 0.70 a |
| 2 | 106.18 a | 6.29 ab | 0.55 b |
| 4 | 88.13 ab | 6.89 a | 0.47 bc |
| 5 | 80.38 b | 6.30 ab | 0.39 c |
| 6 | 87.63 ab | 4.13 b | 0.50 bc |
| ANOVA (p-values) | * | * | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wacal, C.; Ogata, N.; Basalirwa, D.; Sasagawa, D.; Ishigaki, T.; Handa, T.; Kato, M.; Tenywa, M.M.; Masunaga, T.; Yamamoto, S.; et al. Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy. Agronomy 2019, 9, 184. https://doi.org/10.3390/agronomy9040184
Wacal C, Ogata N, Basalirwa D, Sasagawa D, Ishigaki T, Handa T, Kato M, Tenywa MM, Masunaga T, Yamamoto S, et al. Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy. Agronomy. 2019; 9(4):184. https://doi.org/10.3390/agronomy9040184
Chicago/Turabian StyleWacal, Cosmas, Naoki Ogata, Daniel Basalirwa, Daisuke Sasagawa, Tadashi Ishigaki, Takuo Handa, Masako Kato, Moses Makooma Tenywa, Tsugiyuki Masunaga, Sadahiro Yamamoto, and et al. 2019. "Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy" Agronomy 9, no. 4: 184. https://doi.org/10.3390/agronomy9040184