4.1. Effect of Biochar on Soil Properties
Biochar addition clearly improved tropical soil properties in general. Improvements of soil pH and CEC on acidic tropical soil after biochar treatment were in agreement in many biochar experiments, either in a pot [
29] or field study [
30]. Increases of total organic C in all biochar treated soils revealed that biochar contribute to the organic C pool in the soil. The result of organic C is in agreement in many findings, which showed a significant increase of the total organic C pool in the soil environment [
31]. The same observation on total soil N also matches those reported in earlier studies [
8]. Despite the palm kernel shell biochar comprised of higher initial total C and N (
Table 1) than rice husk biochar, the pairwise comparison of total organic C and N were not significant between soils treated with both biochar. On the other hand, greater organic C concentrations may lead to the elevation of soil microbial activity [
32], causing higher N demand in soil, hence immobilized and reduced available N for plant.
Insignificant changes of NH
4+—N following biochar addition in PK and RH is consistent with the data reported from the tropical soils of Zambia and Indonesia [
30]. Greater NH
4+—N concentration in FPK and FRH than PK and RH indicate that ammonification of N does not occur when biochar is applied alone. On the other hand, the association between CEC and NH
4+—N cation is not observed although the negatively charged biochar surfaces has been suggested to allow retention of cations such as NH
4+—N [
33]. This may due to the low concentration of native NH
4+—N in the investigated tropical soil. Supplementation of additional fertilizer enhanced N mineralization through ammonification process. Incorporation of fertilizer to soil with biochar amendment significantly altered NO
3−—N concentration. Significant increment of NO
3−—N in all biochar treated soil than F is likely due to the initial concentration of NO
3−—N from the biochar itself (
Table 1) and not due to mineralization of N through the nitrification of NH
4+—N, while a greater concentration in soils treated with rice husk biochar (RH and FRH) than palm kernel shell biochar (PK and FPK) is due to the double fold initial concentration of NO
3−—N. These findings further confirm the association of a high C:N ratio of organic input with N immobilization in soil.
Increased of P availability upon biochar treatment has been commonly reported, especially in the P deficient soil of the humid tropics [
29]. Biochar is capable of increasing the soil P availability through either its direct inputs or its retaining potential of P applied [
34]. However, the increment of soil P availability due to the biochar input itself is unlikely as the difference of available P in PK and RH is not significantly different. Greater P availability in FPK may suggest the retention of P from the fertilizer applied as a palm kernel shell biochar possess larger BET surface area (
Table 1), hence able to hold greater P in the soil. The surface property, such as the microporous structure of the biochar, has been demonstrated to determine its capacity in P adsorption [
35].
4.2. Effect of Biochar on Soil Microbial Properties
Although made up recalcitrant C with high stability, biochar comprises small fractions of readily mineralized labile active C, which is bioavailable for microbial utilization and serves as a substrate [
36]. Application of rice husk biochar with or without fertilization had a greater significant effect on soil microbial biomass C than the plot treated with palm kernel shell biochar. The greater microbial biomass size can be linked with the higher labile active C input of rice husk biochar compared than in the palm kernel biochar.
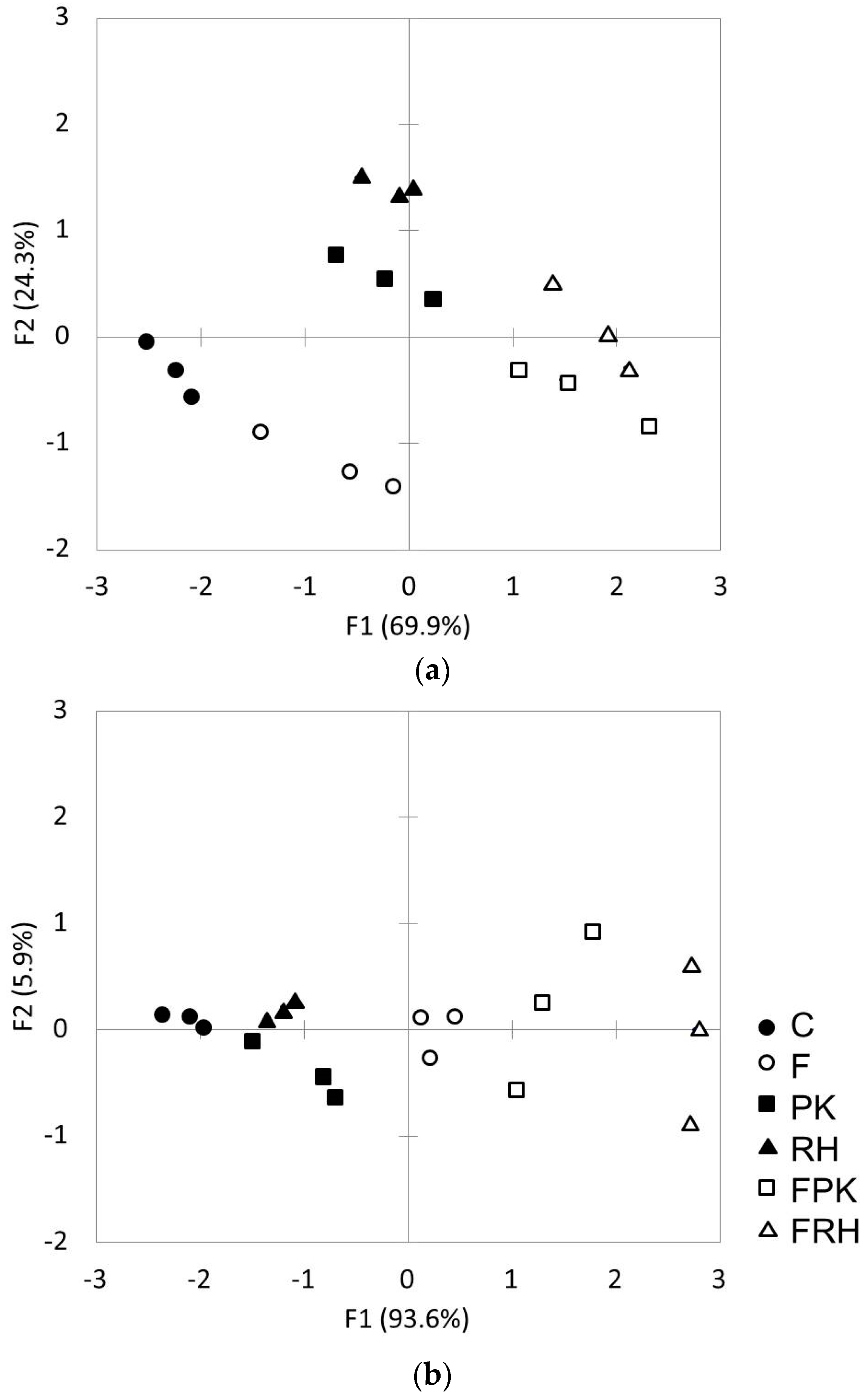
The PCA in
Figure 1 suggested a variation of the soil microbial biomass when biochar is co-applied with fertilizer, especially for microbial biomass N as indicated in component axis 1. The enhancement of microbial biomass N status of soil in FRH and FPK treatment indicate a strong stimulation of biochar in combination with the fertilizer. This finding indirectly points out the mineralization of N by microorganisms due to the fertilization input. Amendment of fertilizer reduced the C:N ratio of soil and hence stimulated N mineralization as supported by the result of NH
4+—N concentration in FPK and FRH (
Table 2). Although Spohn and Kuzyakov [
37] suggest that mineralization of P appears to have been driven by the microbial demand for C, in which organic moiety of phosphorylated organic carbon will be used as a carbon source, this reason however cannot be inferred for the microbial biomass P in this study as the carbon is not limited in the soil due to the abundant organic carbon pool of the biochar applied. The available P result however points out the possible association with microbial biomass P, which supports the possible mineralization of P (
r = 0.9129,
p < 0.01), hence, suggesting that the size of the microbial biomass P is directly associated with the availability of P to plants [
38]. In fact, the microbial biomass P can be re-mineralized in a short turnover period [
39], consequently serving as an available P for plant.
The microbial biomass ratio is considered as an important indicator of microbial nutritional balance [
10]. The soil microbial biomass data showed that the priming effect after biochar addition was higher on microbial biomass N than C, subsequently decreasing the microbial biomass C:N. High microbial biomass C:N ratio in the control soil may indicate an unbalanced soil nutritional status in which the native soil microorganisms could possibly compete for available nitrogen with the plant [
40]. This situation may interfere with plant nutritional requirements due to the nitrogen immobilization, which can become a limiting factor for the plant and the soil microorganisms. Previously, Li and co-authors [
41] quantitatively showed a negative correlation of microbial biomass C:N with plant agronomic output, where a high ratio resulted in crop nitrogen deficiency. In this study, high reduction of microbial biomass C:N was achieved when biochar was co-applied with fertilizer. The microbial biomass C:N ratio in the plot treated with both biochar and fertilizer (FPK and FRH) proposed that fertilizer application is vital to create a nutritional balance in the soil due to the high C:N content of the applied biochar (
Table 1). The statistically significant relationship found between microbial biomass N and maize biomass points out that microbial biomass can act as a significant sink and source of N [
10]. However, the increment of microbial biomass N could not actually reflect the degree of soil fertility as the greater size of microbial biomass N may also indicates N immobilization [
41], by which N is taken up by soil organisms and therefore inaccessible for crop uptake. On the other hand, the C:P and N:P are rarely systematically assessed in the agricultural soil, although the ecological stoichiometric relationships have been broadly studied in various terrestrial ecosystems [
42]. Therefore, the significance of these two molar ratios in the agricultural soil cannot be ruled out in this short-term experiment. In fact, no significant correlation of these molar ratios with a plant biomass yield was observed (
Table 6).
An available nutritional substrate found in the soil environment is utilized by the soil microorganisms through the hydrolytic degradation of the microbial enzymes. This hydrolytic process is widely regarded as a proximate indicator of elemental mineralization and nutrient cycling [
9]. The three enzymes selected in this study (β-glucosidase, urease, phosphodiesterase) are related to the transformation of soil C, N, and P [
43]. Higher β-glucosidase activity in all biochar treated soils can be linked to the boost of organic C pool in the soil and therefore may result in the increment of a distinctive subset of cellulolytic microorganisms from the soil environment. Urease activity change in response to treatment appears to result from fertilizer incorporation. Incorporation of fertilizer had a significant effect on the soil urease activity compared to when biochar was added alone, suggesting that the addition of urea in the fertilization regime enhanced the activity of many enzymes catalyzing N. However, single application of biochar without fertilization still showed significant difference with the control soil and this result is consistent with urease and phosphodiesterase activity data obtained in many previous studies [
44,
45,
46]. However, a significant difference of phosphodiesterase activity due to the phosphate addition in the biochar amended soil co-applied with fertilizer (FPK and FRH) was not observed.
Quantitative PCR has been widely used as a promising approach to quantify soil microbial community structure and function at broad taxonomic levels. The
16S rRNA and ITS are two commonly used conserved regions in enumerating the population density of the soil bacteria and fungi, respectively [
28]. Data on the abundance of the gene copy number of both bacteria and fungi shows a strong stimulation of soil microbial population density by palm kernel shell biochar. The heterogeneous characteristic of biochar, such as the porosity, has been suggested to serve as a colonizing site for microbiota, hence, acting as a potential habitat [
47]. Biochar was also discovered to be an ideal ecological niche by not only allowing microorganisms to establish themselves on the pores and surfaces, but also may retain readily metabolized nutrient [
48] and substrate [
49], therefore, stimulating the total size of soil microbial population. A number of researchers reported an increment in the abundance of these two conserved genes after the application of biochar to the soil [
50,
51], however, there is no research available reporting the effect of a biochar addition on microbial gene abundance in the acidic soil of the humid tropic.
4.3. Effect of Biochar on Maize Biomass Yield and its Relationship with Microbial Properties
Enhanced maize growths upon biochar addition in the tropical soil have been reported in a controlled glasshouse [
7] as well as in a field trial [
30]. In fact, biochar effects on crops planted in this climatic region are found to be more profound from those tested in temperate regions [
52].
Insignificant N uptake in PK and RH compared to the control is supported by the higher microbial biomass C:N in which limited available N is revealed. Enhancement of the N uptake in FPK and FRH can be reflected by the reduction of the microbial biomass C:N upon fertilizer supplementation. For P uptake, although FPK recorded a higher available P in the soil, the result showed that the P uptake in FRH is more significant. The increased microbial biomass P in the present study may indicate the immobilization of some of the P in the soil. However, most of the immobilized P may be available due to the short-term turnover of microbial biomass P [
53]. This observation is reflected by the fact that both available P and microbial biomass P did not show significant correlation in the maize biomass during the harvest stage.
Soil organic C and total N properties are commonly regarded as the main soil fertility factors in the context of crop productions [
54]. The insignificant correlation between plant biomass and all measured soil parameters (C, N and P) suggests that significant changes cannot precisely indicate the effect of the soil organic matter dynamic due to soil management practices within a short time. In the present study however, we demonstrated that no significant correlation of these two soil properties can be established with crop biomass. The microbial biomass C:N ratio was shown to be a sensitive indicator capable of assessing plant growth. Negative correlation of maize biomass with microbial biomass C:N points out that the high C:N ratios of biochar lead to nitrogen immobilization in the soil and are disadvantageous to agricultural production. This matches those observed in earlier studies, which used other types of amended organic material [
41,
55].

 ,
, 


{kind=link}