Comparative Analysis of the Transcriptional Response of Tolerant and Sensitive Wheat Genotypes to Drought Stress in Field Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Samples and RNA Isolation
2.2. RNA Deep Sequencing and Data Analysis
2.3. Identification of Differentially Expressed Genes (DEGs)
2.4. Analysis of RNA Editing Sites
3. Results and Discussion
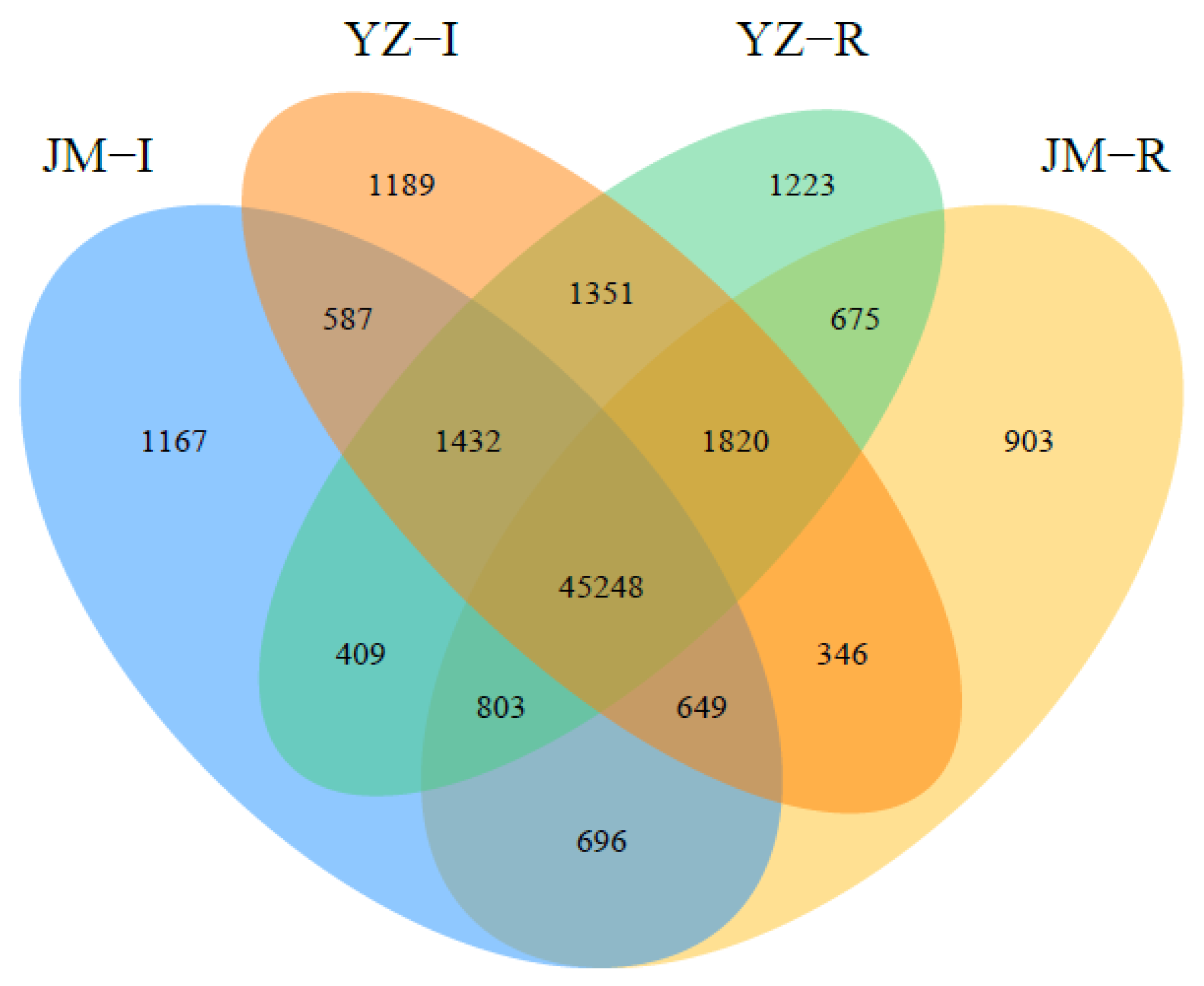
3.1. RNA-seq Analysis
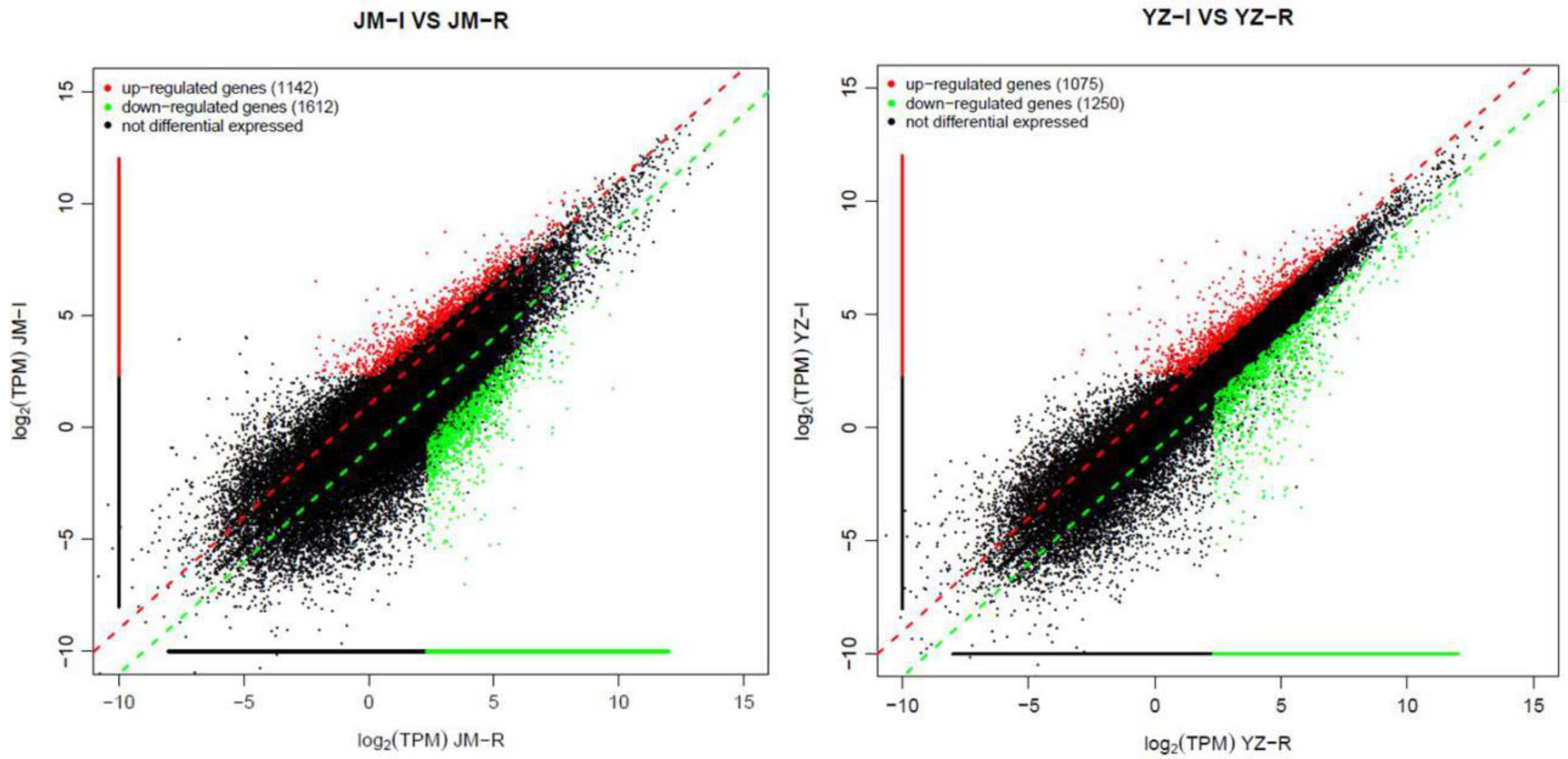
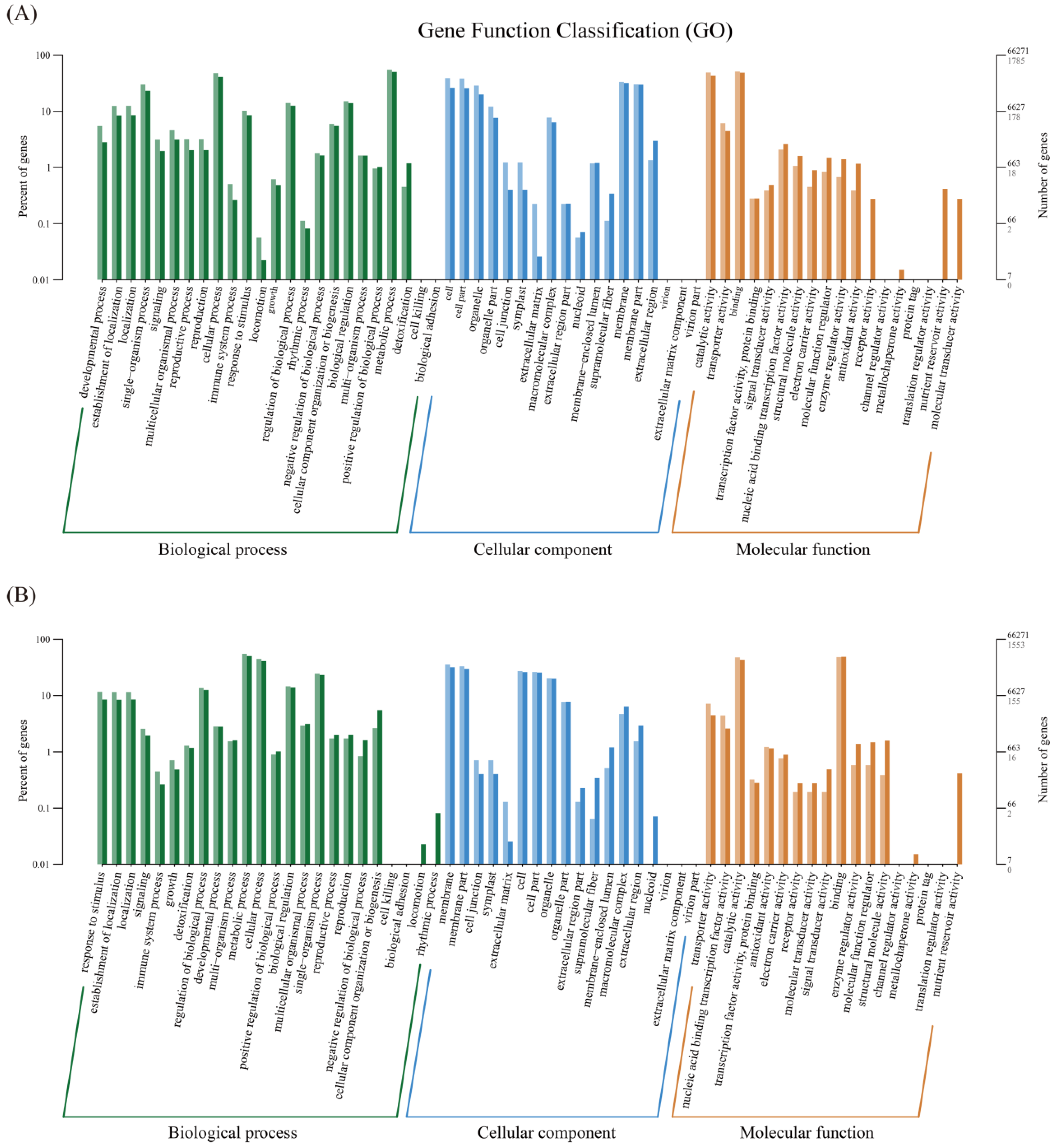
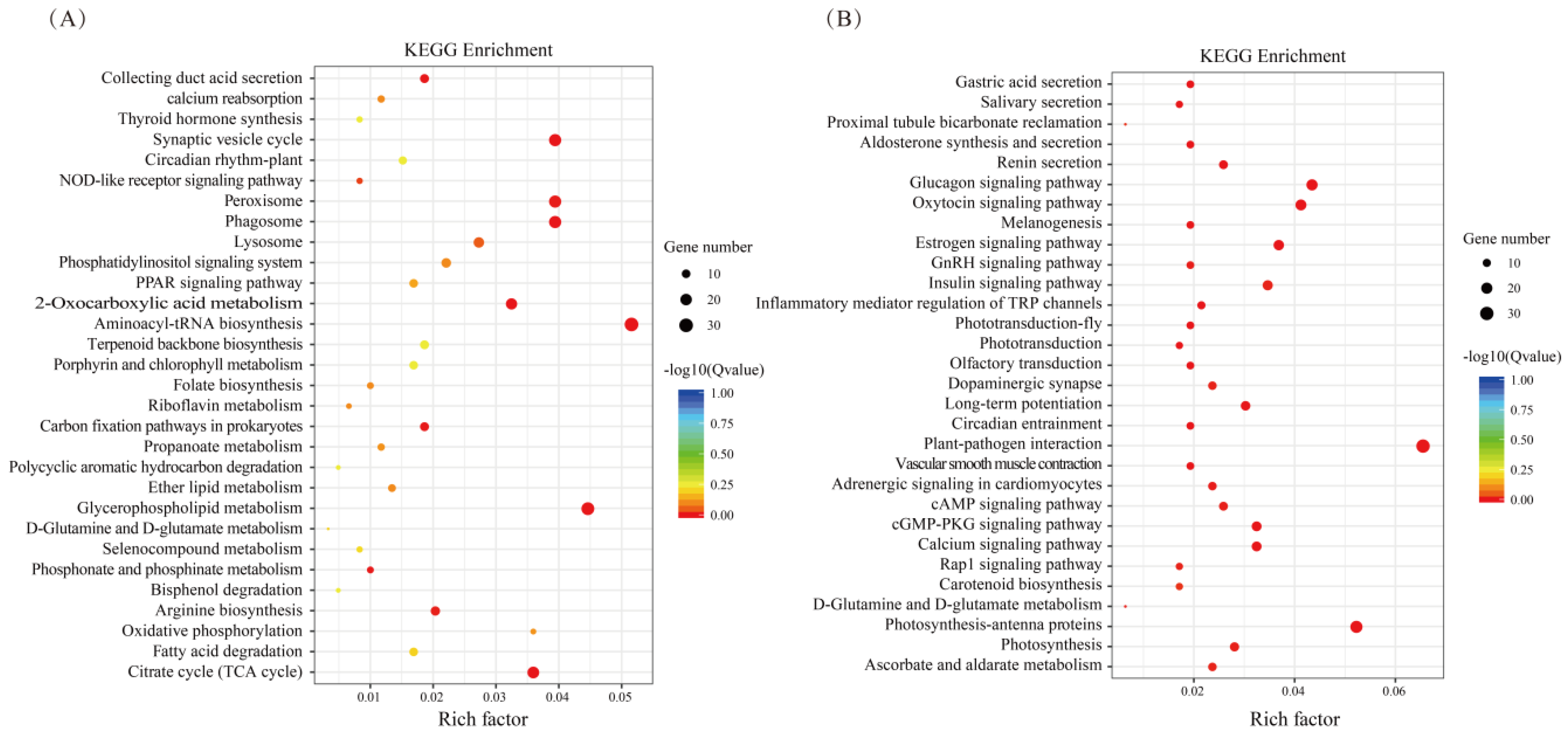
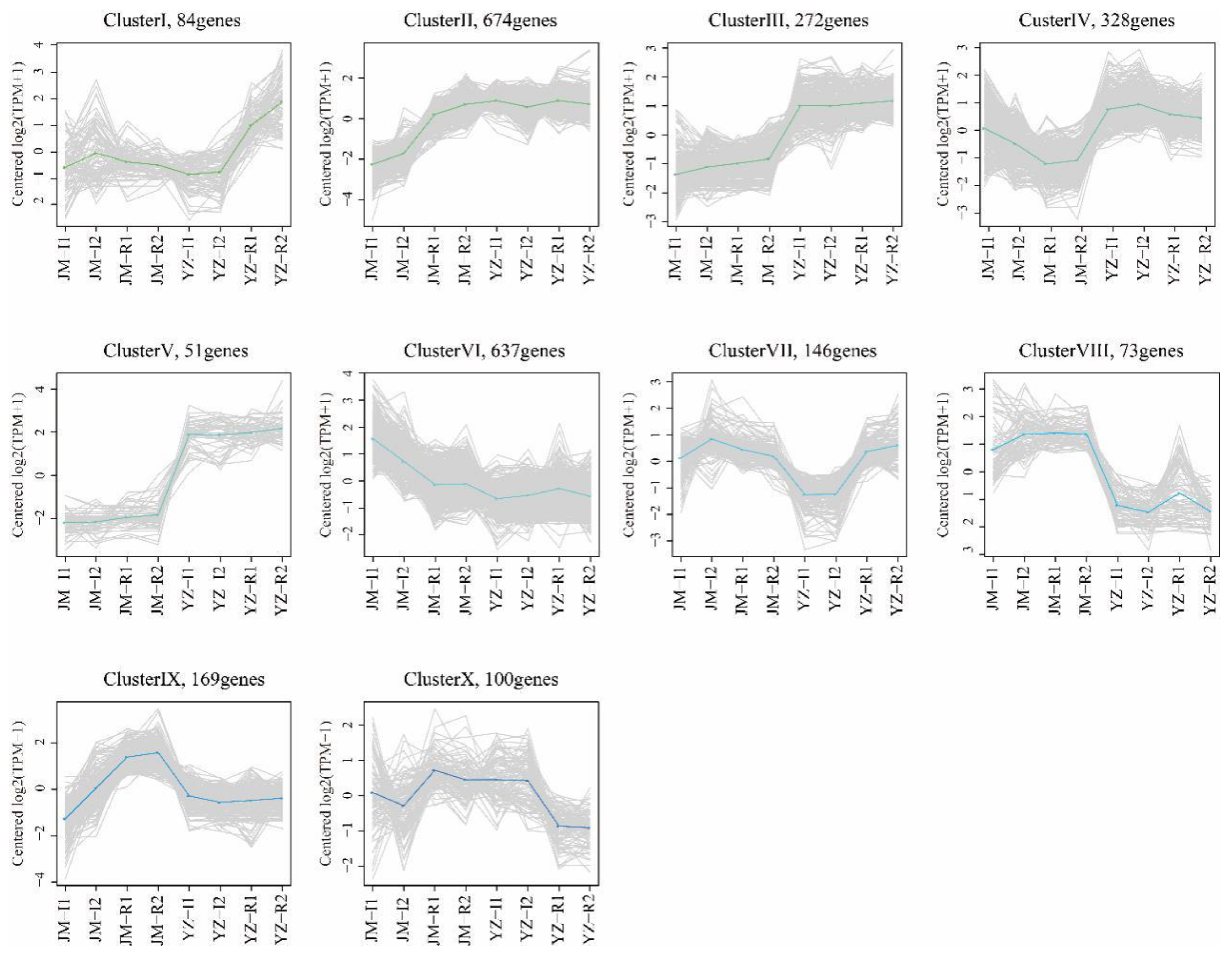
3.2. Identification of the Differentially Expressed Genes
3.3. Analysis of RNA Editing Sites
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trenberth, K.E.; Dai, A.; Schrier, G.V.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Cropland data in Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/LR (accessed on 15 September 2018).
- Thao, N.P.; Tran, L.S. Enhancement of plant productivity in the post-genomics era. Curr. Genom. 2016, 17, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Manoj, K.; Raju, S.; Satoshi, O.; Yusaku, U.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, R.; Wang, H.; Li, D.; Wang, X.; Zhang, Y.; Zhen, W.; Duan, H.; Yan, G.; Li, Y. Transcriptomics analyses reveal wheat responses to drought stress during reproductive stages under field conditions. Front. Plant Sci. 2017, 8, 592. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, D.; Pandey, A.; Choudhary, M.K.; Datta, A.; Chakraborty, S.; Chakraborty, N. Comparative proteomics analysis of differentially expressed proteins in chickpea extracellular matrix during dehydration stress. Mol. Cell. Proteom. 2007, 6, 1868–1884. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Maud, V.; Hazel, K.S.; Cohen, D.; Dewoody, J.; Trewin, H.; Steenackers, M.; Bastien, C.; Taylor, G. Adaptive mechanisms and genomic plasticity for drought tolerance identified in European black poplar (Populus nigra L.). Tree Physiol. 2016, 36, 909–928. [Google Scholar]
- Naimat, U.; Meral, Y.Z.; Neslihan, Ö.G.; Budak, H. Comparative metabolite profiling of drought stress in roots and leaves of seven Triticeae species. BMC Genom. 2017, 18, 969. [Google Scholar] [Green Version]
- Ramachandra, R.A.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar]
- Hossain, Z.; Nouri, M.Z.; Komatsu, S. Plant cell organelle proteomics in response to abiotic stress. J. Proteome Res. 2012, 11, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Moumeni, A.; Satoh, K.; Kondoh, H.; Asano, T.; Hosaka, A.; Venuprasad, R.; Serraj, R.; Kumar, A.; Leung, H.; Kikuchi, S. Comparative analysis of root transcriptome profiles of two pairs of drought-tolerant and susceptible rice near-isogenic lines under different drought stress. BMC Plant Biol. 2011, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Ramanjulu, S.; Bartels, D. Drought- and desiccation-induced modulation of gene expression in plants. Plant Cell Environ. 2002, 25, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Ergen, N.Z.; Thimmapuram, J.; Bohnert, H.J.; Budak, H. Transcriptome pathways unique to dehydration tolerant relatives of modern wheat. Funct. Integr. Genom. 2009, 9, 377–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampino, P.; Pataleo, S.; Gerardi, C.; Mita, G.; Perrotta, C. Drought stress response in wheat: Physiological and molecular analysis of resistant and sensitive genotypes. Plant Cell Environ. 2006, 29, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Kantar, M.; Kurtoglu, K.Y. Drought tolerance in modern and wild wheat. Sci. World J. 2013, 2013, 548246. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.X.; Wang, Y.P.; He, Q.; Li, H.J.; Zhang, X.J.; Zhang, F. Comparative proteomics illustrates the complexity of drought resistance mechanisms in two wheat (Triticum aestivum L.) cultivars under dehydration and rehydration. BMC Plant Biol. 2016, 16, 188. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Appels, R.; Botha-Oberholster, A.M.; Buell, C.R.; Bennetzen, J.L.; Chalhoub, B.; Chumley, F.; Dvorák, J.; Iwanaga, M.; Keller, B.; et al. A workshop report on wheat genome sequencing: International genome research on wheat consortium. Genetics 2014, 168, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.; Rogers, J.; Doležel, J.; Pozniak, C.; Eversole, K.; Feuillet, C.; Gill, B.; Friebe, B.; Lukaszewski, A.J.; Sourdille, P.; et al. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Zampieri, M.; Ceglar, A.; Dentener, F.; Toreti, A. Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. 2017, 12, 064008. [Google Scholar] [CrossRef] [Green Version]
- Manes, Y.; Gomez, H.; Puhl, L.; Reynolds, M.; Braun, H.; Trethowan, R. Genetic yield gains of the CIMMYT international semi-arid wheat yield trials from 1994 to 2010. Crop Sci. 2012, 52, 1543–1552. [Google Scholar] [CrossRef]
- Reynolds, M.; Foulkes, M.J.; Slafer, G.A.; Berry, P.; Parry, M.A.; Snape, J.W.; Angus, W.J. Raising yield potential in wheat. J. Exp. Bot. 2009, 60, 1899–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morran, S.; Eini, O.; Pyvovarenko, T.; Parent, B.; Singh, R.; Ismagul, A.; Eliby, S.; Shirley, N.; Langridge, P.; Lopato, S. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J. 2011, 9, 230–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.M.; Wang, Y.F.; Xu, P.; Zhang, Z.B. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, B.A.; Avsar, B.; Lucas, S.J.; Budak, H. Plant abiotic stress signaling. Plant Signal. Behav. 2012, 7, 1450–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Yin, Y.J.; Liu, X.Y.; Tong, S.M.; Xing, J.W.; Zhang, Y.; Pudake, R.N.; Izquierdo, E.M.; Peng, H.R.; Xin, M.M.; et al. The E3 ligase TaSAP5 alters drought stress responses by promoting the degradation of DRIP proteins. Plant Physiol. 2017, 175, 1878–1892. [Google Scholar] [CrossRef] [PubMed]
- Rahaie, M.; Xue, G.P.; Naghavi, M.R.; Alizadeh, H.; Schenk, P.M. A MYB gene from wheat (Triticum aestivum L.) is up-regulated during salt and drought stresses and differentially regulated between salt-tolerant and sensitive genotypes. Plant Cell Rep. 2010, 29, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Dalal, M.; Sahu, S.; Tiwari, S.; Rao, A.R.; Gaikwad, K. Transcriptome analysis reveals interplay between hormones, ROS metabolism and cell wall biosynthesis for drought-induced root growth in wheat. Plant Physiol. Biochem. 2018, 130, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Xie, Y.; Fan, S.; Wang, Z.; Wang, F.; Zhang, B.; Li, H.; Song, J.; Kong, L. Comparative analysis of root transcriptome profiles between drought-tolerant and susceptible wheat genotypes in response to water stress. Plant Sci. 2018, 272, 276–293. [Google Scholar] [CrossRef] [PubMed]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Ford, K.L.; Cassin, A.; Bacic, A. Quantitative proteomic analysis of wheat cultivars with differing drought stress tolerance. Front. Plant Sci. 2011, 2, 44. [Google Scholar] [CrossRef] [PubMed]
- Genome Sequence Archive (GSA) Database. Available online: http://bigd.big.ac.cn/gsa (accessed on 1 October 2018).
- Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Brock, G.; Datta, S.; Pihur, V.; Datta, S. clValid: An R package for cluster validation. J. Stat. Softw. 2008, 25, 1–22. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.Y.; You, Q.; Yi, X.; Du, Z.; Xu, W.Y.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Lu, Y.L.; Yan, S.J.; Xing, Q.H.; Tian, W.D. SPRINT: An SNP-free toolkit for identifying RNA editing sites. Bioinformatics 2017, 3, 3538–3548. [Google Scholar] [CrossRef] [PubMed]
- BWA (Burrows-Wheeler Aligner). Available online: http://bio-bwa.sourceforge.net/ (accessed on 1 September 2018).
- Samtools. Available online: http://www.htslib.org. (accessed on 1 September 2018).
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; van Ex, F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Härtel, B.; Brennicke, A. RNA editing in plants and its evolution. Annu. Rev. Genet. 2013, 47, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, H.; Ge, L.; Xing, G.; Wang, M.; Weining, S.; Nie, X. Identification and analysis of RNA editing sites in the chloroplast transcripts of Aegilops tauschii L. Genes 2016, 30, 8. [Google Scholar] [CrossRef] [PubMed]
- Kumbhar, F.; Nie, X.; Xing, G.; Zhao, X.; Lin, Y.; Wang, S.; Weining, S. Identification and characterization of RNA editing sites in chloroplast transcripts of einkorn wheat (Triticum monococcum). Ann. Appl. Biol. 2018, 172, 197–207. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Reads | Raw Base (bp) | Clean Reads | Clean Bases (bp) | Total Mapped to Wheat Genome | Multiple Mapped | Uniquely Mapped | Reads Mapped in Proper Pairs | |
|---|---|---|---|---|---|---|---|---|
| JM-I2 | 54,939,280 | 7,913,976,131 | 50,542,262 | 7,278,085,728 | 40,962,623 (81.05%) | 1,908,065 (3.78%) | 39,054,558 (77.27%) | 34,136,584 (67.54%) |
| JM-I1 | 49,544,428 | 7,152,858,915 | 47,469,372 | 6,835,589,568 | 38,093,160 (80.25%) | 1,458,216 (3.07%) | 36,634,944 (77.18%) | 32,711,652 (68.91%) |
| YZ-R1 | 52,387,108 | 7,582,519,380 | 51,913,178 | 7,475,497,632 | 41,618,432 (80.17%) | 1,576,802 (3.04%) | 40,041,630 (77.13%) | 36,267,234 (69.86%) |
| YZ-I1 | 53,867,378 | 7,742,552,474 | 51,768,216 | 7,454,623,104 | 41,280,310 (79.74%) | 1,858,852 (3.59%) | 39,421,458 (76.15%) | 34,636,332 (66.91%) |
| YZ-I2 | 53,900,538 | 7,810,800,148 | 51,550,574 | 7,474,833,230 | 41,415,518 (80.34%) | 1,488,853 (2.89%) | 39,926,665 (77.45%) | 35,443,404 (68.75%) |
| YZ-R2 | 53,939,098 | 7,789,354,276 | 47,698,318 | 6,868,557,792 | 38,634,459 (81.00%) | 1,698,950 (3.56%) | 36,935,509 (77.44%) | 32,060,558 (67.22%) |
| JM-R2 | 55,766,550 | 8,024,037,795 | 44,623,214 | 6,425,742,816 | 36,373,597 (81.51%) | 2,175,078 (4.87%) | 34,198,519 (76.64%) | 28,892,192 (64.75%) |
| JM-R1 | 51,561,402 | 7,422,811,668 | 41,653,874 | 5,998,157,856 | 34,227,750 (82.17%) | 2,045,728 (4.91%) | 32,182,022 (77.26%) | 27,307,370 (65.56%) |
| Average | 53,238,223 | 7,679,863,848 | 48,402,376 | 6,976,385,966 | ||||
| Total | 425,905,782 | 61,438,910,787 | 387,219,008 | 55,811,087,726 |
| Gene ID | Genotype a | Position b | Samples c | Editing Site d | NR_annotation | |||
|---|---|---|---|---|---|---|---|---|
| I1 | I2 | R1 | R2 | |||||
| TraesCS3D01G002100.1 | JM | 912 | A/7 | A/5 | G/5 | G/9 | acA/acG | Receptor kinase-like protein |
| TraesCS3D01G002100.1 | JM | 507 | G/9 | G/5 | C/5 | C/6 | gtG/gtC | Receptor kinase-like protein |
| TraesCS3D01G517100.1 | JM | 660 | G/122 | G/99 | T/76 | T/144 | gcC/gcA | Abscisic stress ripening protein |
| TraesCS4B01G220200.1 | JM | 843 | C/5 | C/7 | T/5 | T/6 | - | Phosphoenolpyruvate carboxykinase (ATP)-like |
| TraesCS4B01G280800.1 | JM | 417 | G/6 | G/8 | C/5 | C/11 | - | 40S ribosomal protein S27 |
| TraesCS4B01G287800.1 | JM | 1356 | C/13 | C/24 | A/16 | A/21 | ggC/ggA | Sucrose transporter |
| TraesCS4B01G292500.1 | JM | 735 | G/9 | G/7 | A/10 | A/13 | - | Transcription initiation factor IIE subunit beta |
| TraesCS4B01G293600.3 | JM | 1893 | G/11 | G/25 | A/19 | A/18 | - | Copper-transporting ATPase |
| TraesCS4B01G293600.3 | JM | 1221 | C/53 | C/78 | T/37 | T/89 | gCa/gTa | Copper-transporting ATPase |
| TraesCS4B01G293600.3 | JM | 135 | C/64 | C/86 | T/46 | T/85 | Cca/Tca | Copper-transporting ATPase |
| TraesCS4D01G268600.1 | JM | 1215 | C/9 | C/21 | A/6 | A/35 | agG/agT | Ribosomal large subunit pseudouridine synthase B |
| TraesCS5A01G232200.1 | JM | 498 | G/5 | G/6 | A/4 | A/7 | tCg/tTg | RING/U-box superfamily protein, putative |
| TraesCS5A01G279200.1 | JM | 1437 | C/5 | C/13 | A/5 | A/8 | - | Basic helix-loop-helix transcription factor |
| TraesCS5A01G501900.1 | JM | 819 | C/5 | C/32 | A/6 | A/35 | Gcc/Tcc | ATP phosphoribosyltransferase |
| TraesCS5A01G509700.1 | JM | 795 | C/8 | C/10 | T/6 | T/13 | cCg/cTg | Thyroid adenoma-associated protein-like protein |
| TraesCS5A01G518300.1 | JM | 213 | G/49 | G/142 | A/50 | A/158 | ggC/ggT | Aquaporin-like protein |
| TraesCS5A01G518300.1 | JM | 1071 | G/5 | G/12 | A/5 | A/15 | - | Aquaporin-like protein |
| TraesCS5D01G043500.1 | JM | 909 | C/5 | C/5 | T/5 | T/11 | gCg/gTg | Receptor-like protein kinase |
| TraesCS5D01G045300.1 | JM | 618 | T/10 | T/45 | C/9 | C/48 | aTg/aCg | NRT1/PTR family protein 2.2 |
| TraesCS6B01G012800.1 | JM | 6282 | T/5 | T/9 | C/5 | C/5 | taC/taT | MYB family transcription factor-like protein |
| TraesCS6B01G069800.1 | JM | 1533 | C/17 | C/5 | T/3 | T/6 | acG/acA | Kinase family protein |
| TraesCS6B01G069800.1 | JM | 1185 | T/8 | T/5 | C/8 | C/16 | - | Kinase family protein |
| TraesCS6B01G078200.1 | JM | 1473 | G/5 | G/9 | A/5 | A/31 | - | Histone H4 |
| TraesCS6B01G078200.1 | JM | 834 | T/8 | T/16 | C/5 | C/50 | - | Histone H4 |
| TraesCS6B01G079400.1 | JM | 969 | A/9 | A/23 | G/5 | G/36 | gTg/gCg | Syntaxin, putative |
| TraesCS6B01G079400.1 | JM | 957 | G/5 | G/23 | T/5 | T/20 | - | Syntaxin, putative |
| TraesCS6B01G079400.1 | JM | 2106 | T/5 | T/21 | C/5 | C/19 | - | Syntaxin, putative |
| TraesCS6B01G434200.1 | JM | 1665 | T/6 | T/5 | C/5 | C/7 | gAg/gGg | Thioredoxin |
| TraesCS7A01G437900.1 | JM | 408 | G/5 | G/14 | C/10 | C/5 | - | Histone H2A |
| TraesCS7D01G399500.1 | JM | 816 | C/8 | C/7 | T/6 | T/7 | gaC/gaT | Eukaryotic aspartyl protease family protein |
| TraesCS6B01G079400.1 | YZ | 1122 | C/8 | C/12 | T/8 | T/15 | - | Syntaxin, putative |
| TraesCS5A01G509700.1 | YZ | 795 | C/11 | C/5 | T/5 | T/8 | cCg/cTg | Thyroid adenoma-associated protein-like protein |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, S.; Feng, K.; Peng, S.; Wang, J.; Zhang, Y.; Bian, J.; Nie, X. Comparative Analysis of the Transcriptional Response of Tolerant and Sensitive Wheat Genotypes to Drought Stress in Field Conditions. Agronomy 2018, 8, 247. https://doi.org/10.3390/agronomy8110247
Lv S, Feng K, Peng S, Wang J, Zhang Y, Bian J, Nie X. Comparative Analysis of the Transcriptional Response of Tolerant and Sensitive Wheat Genotypes to Drought Stress in Field Conditions. Agronomy. 2018; 8(11):247. https://doi.org/10.3390/agronomy8110247
Chicago/Turabian StyleLv, Shuzuo, Kewei Feng, Shaofeng Peng, Jieqiong Wang, Yuanfei Zhang, Jianxin Bian, and Xiaojun Nie. 2018. "Comparative Analysis of the Transcriptional Response of Tolerant and Sensitive Wheat Genotypes to Drought Stress in Field Conditions" Agronomy 8, no. 11: 247. https://doi.org/10.3390/agronomy8110247