

Smilax aspera L. Leaf and Fruit Extracts as Antibacterial Agents for Crop Protection

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Chemicals
2.2. Bacterial Strains
2.3. Preparation of Leaf and Fruit Extracts
2.4. Characterization Procedures
2.5. Evaluation of Antibacterial Activity
2.5.1. In Vitro Tests
2.5.2. In Vivo Tests
2.6. Statistical Analysis
3. Results
3.1. Vibrational Spectroscopy
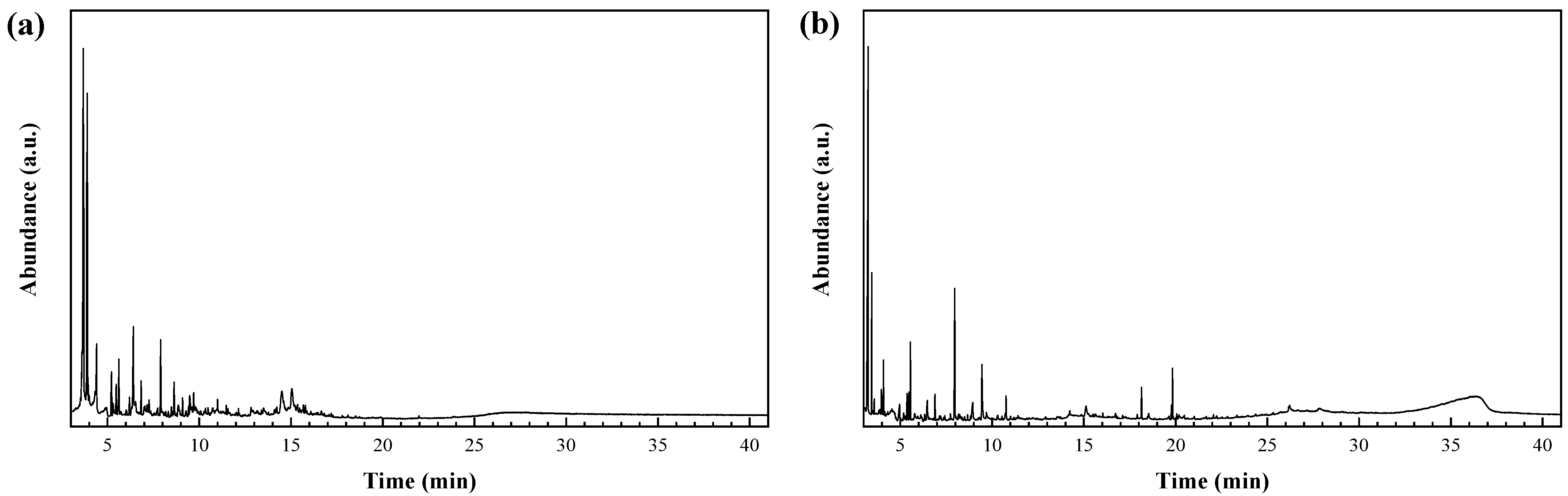
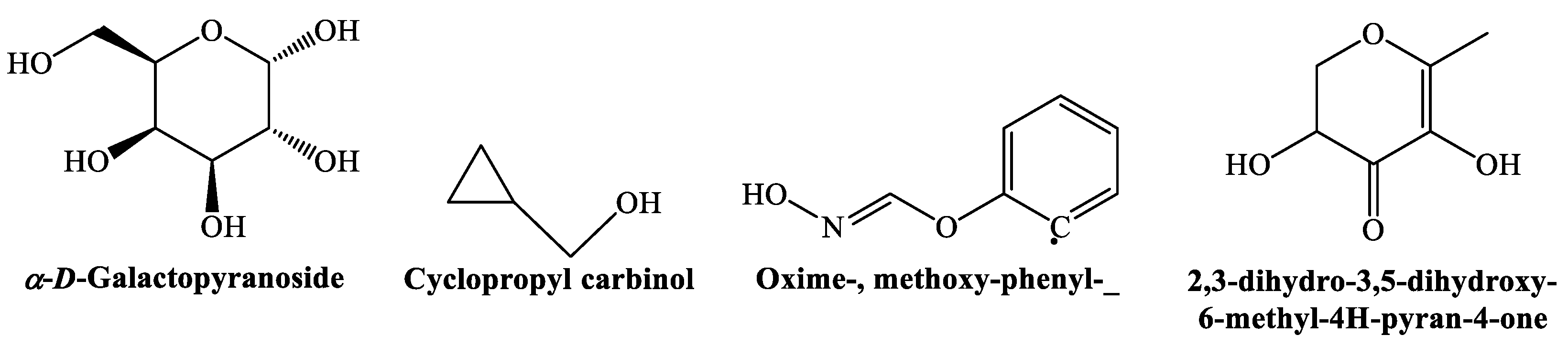
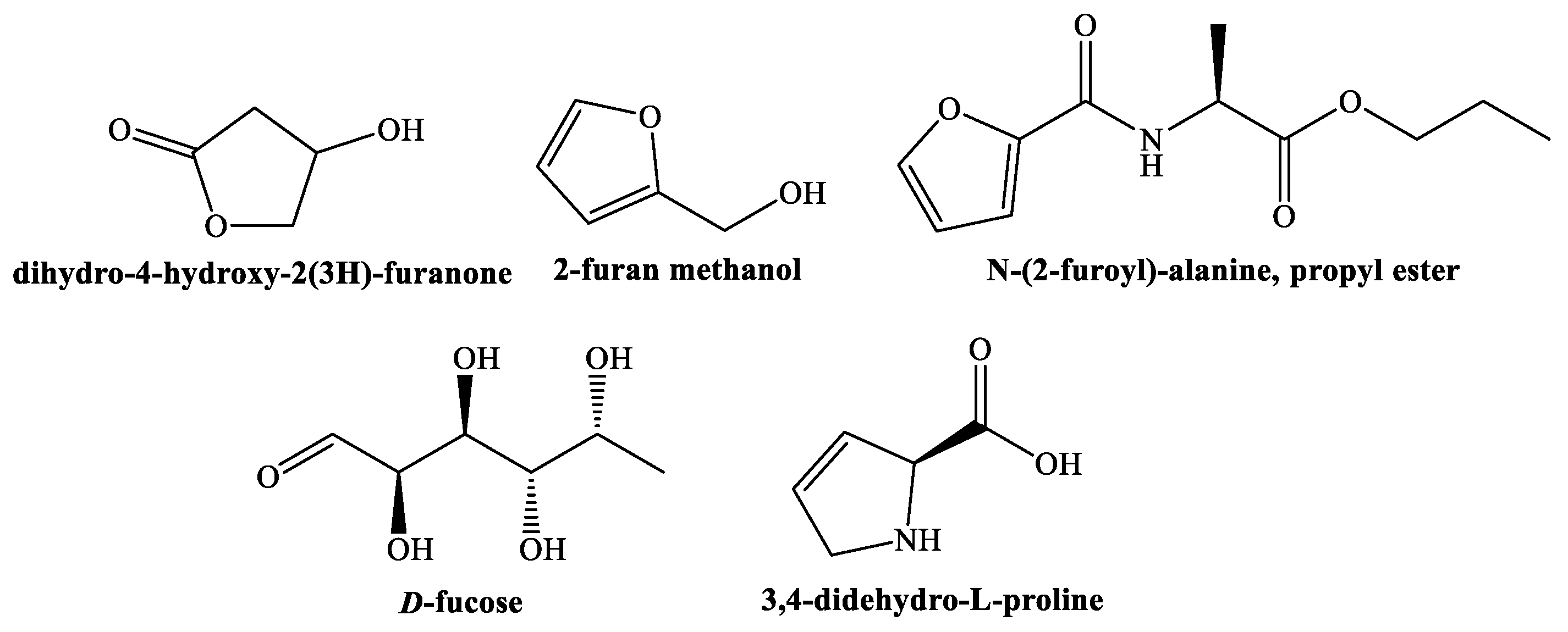
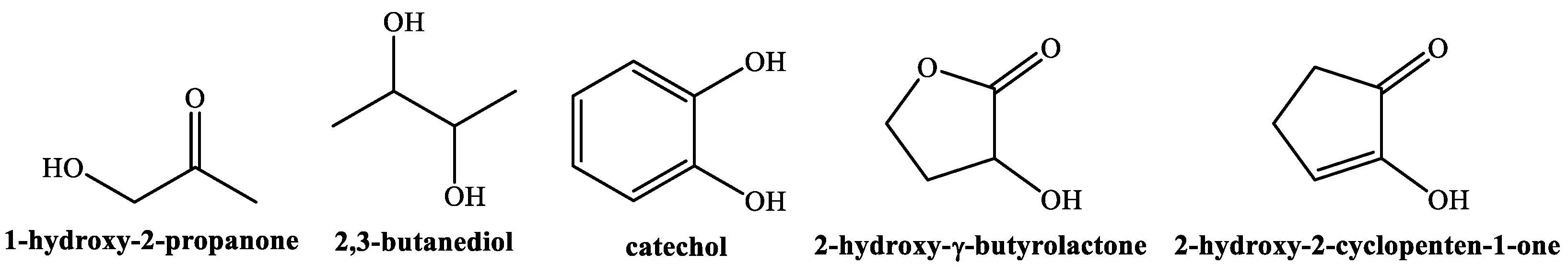
3.2. Phytochemicals Identification
3.3. Antibacterial Activity
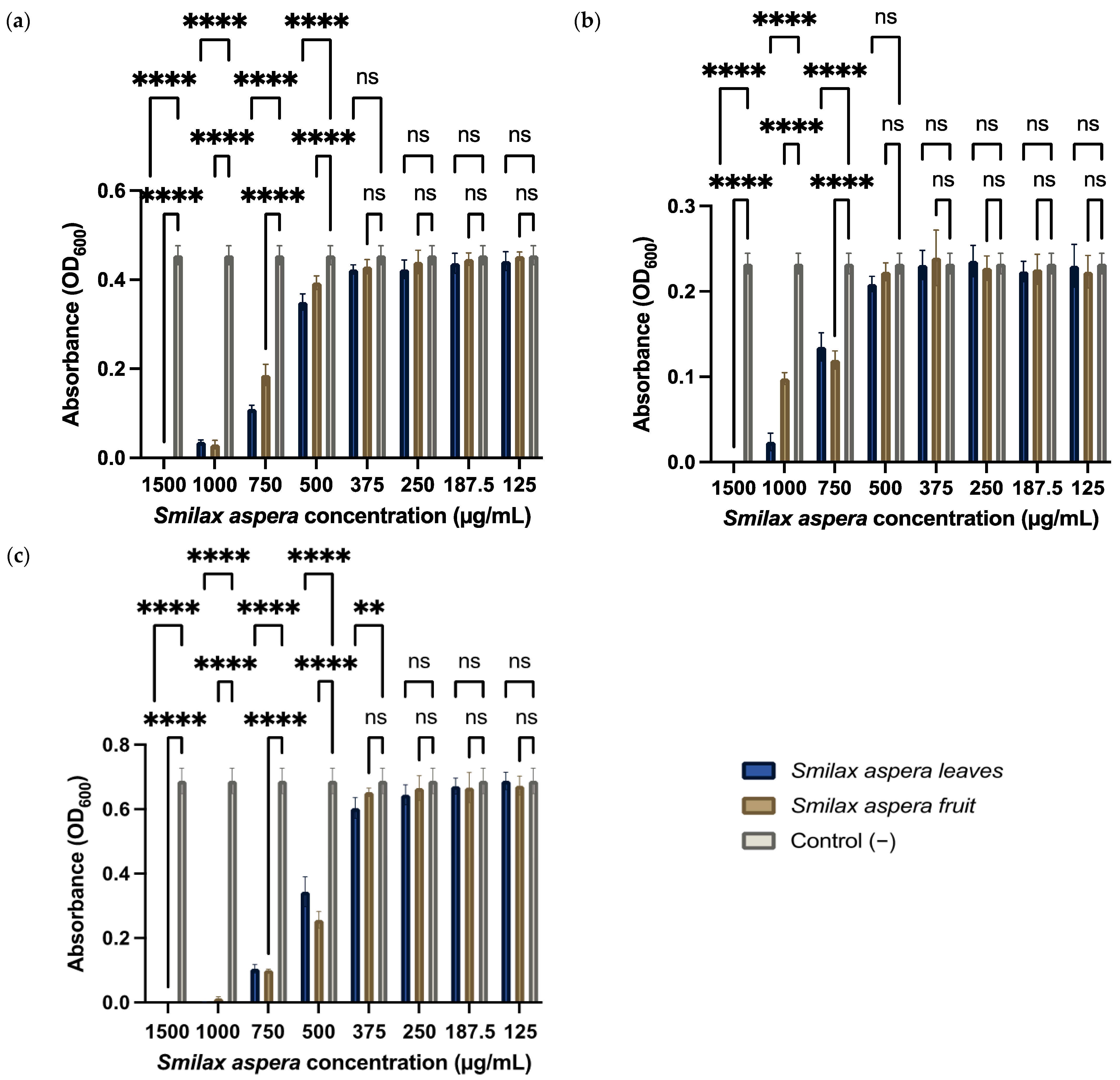
3.3.1. In Vitro Antibacterial Activity
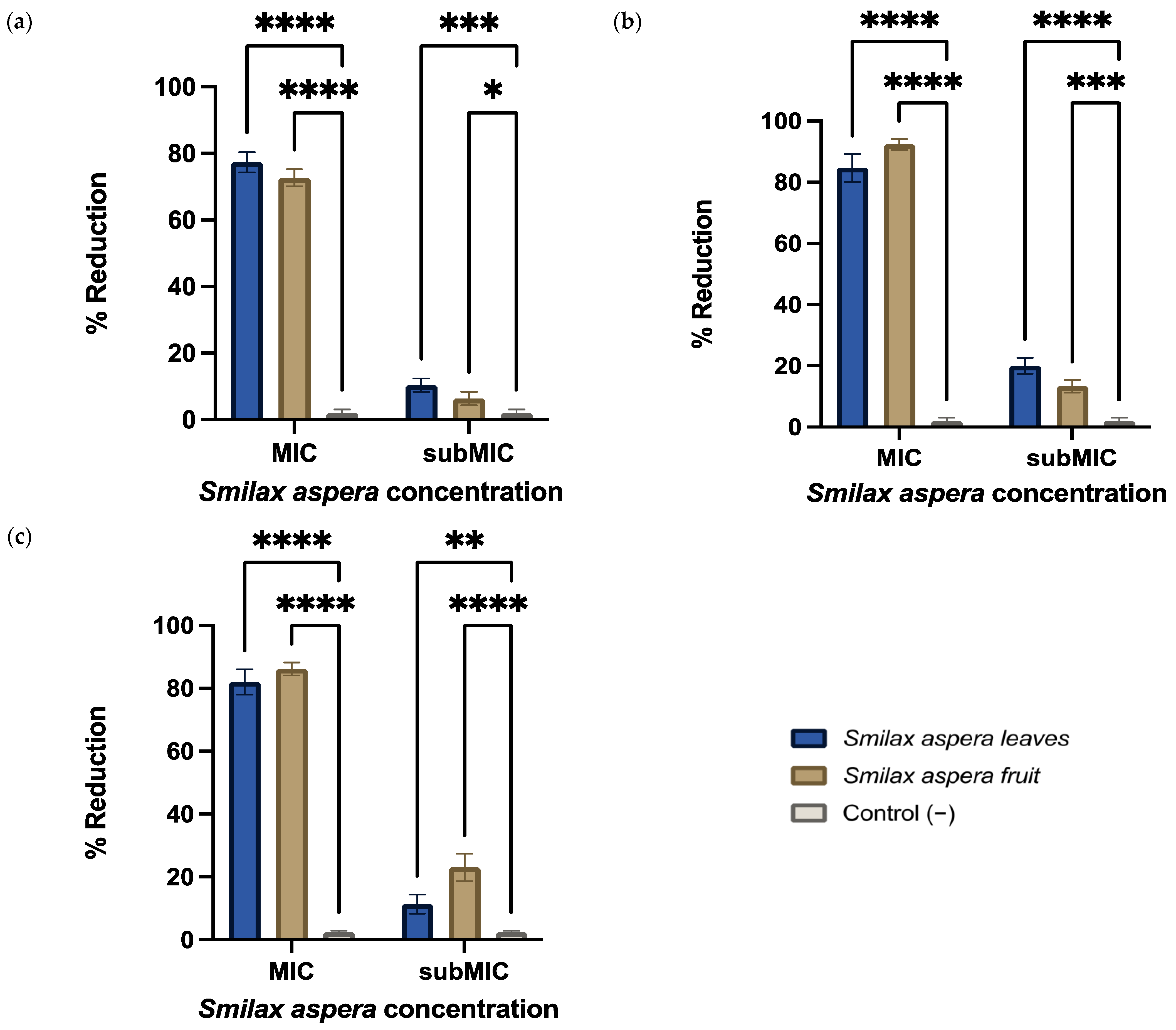
3.3.2. Anti-Biofilm Activity
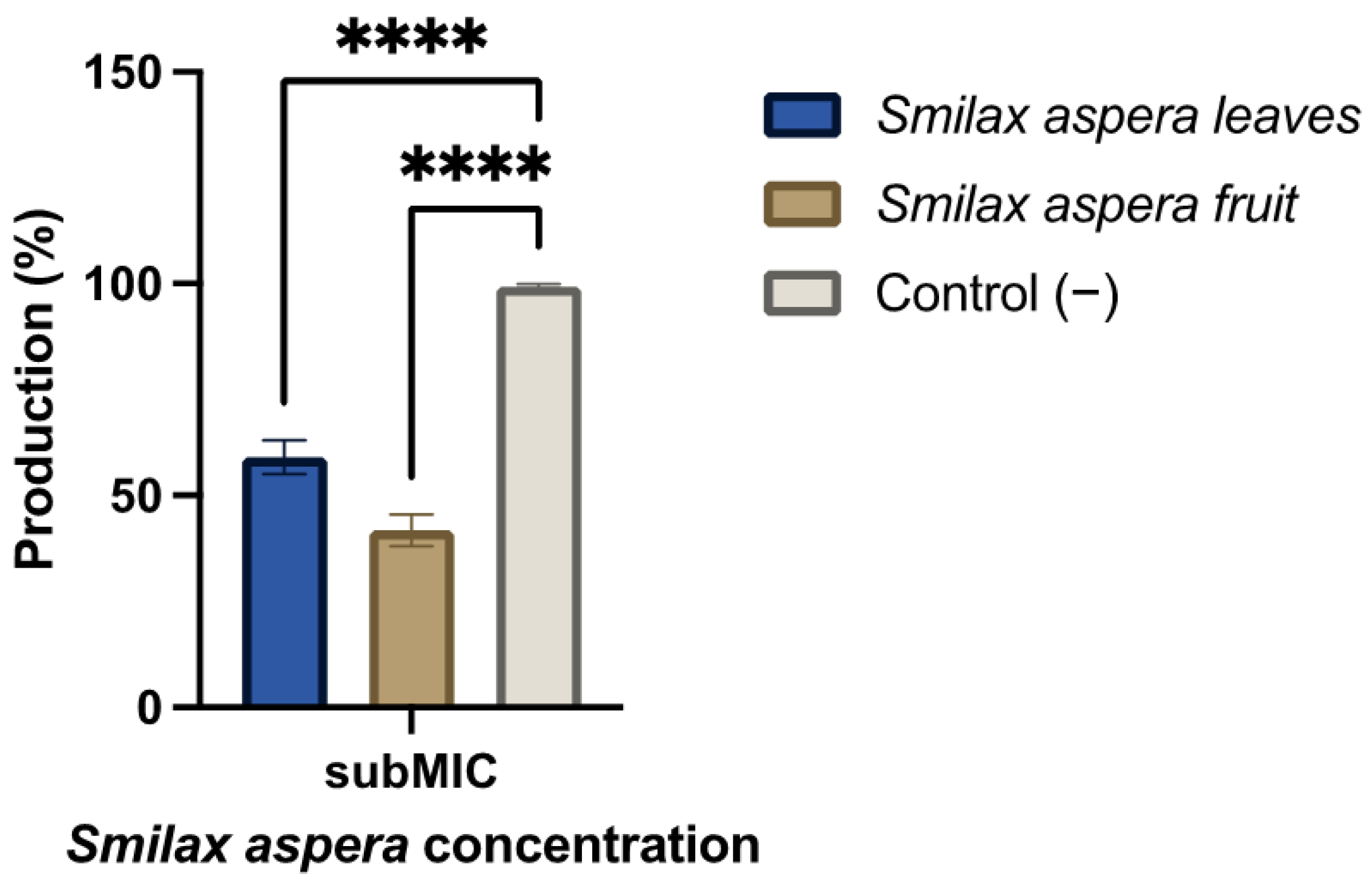
3.3.3. Amylovoran Production
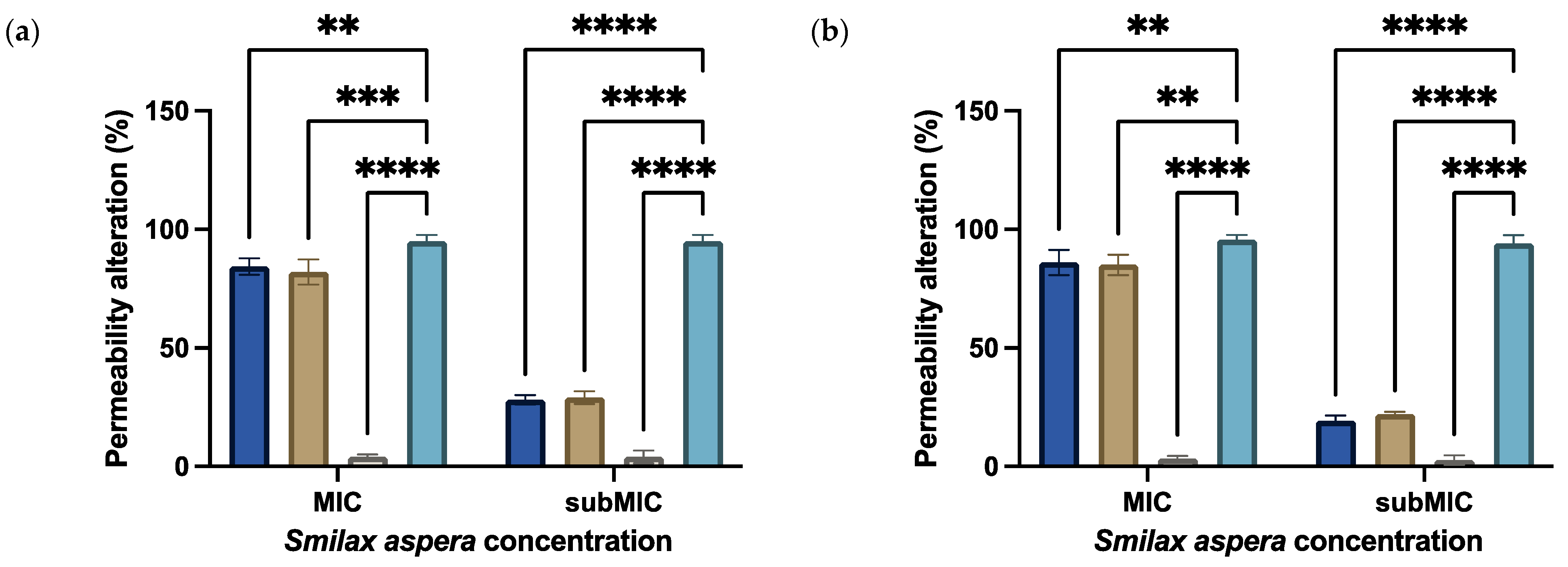
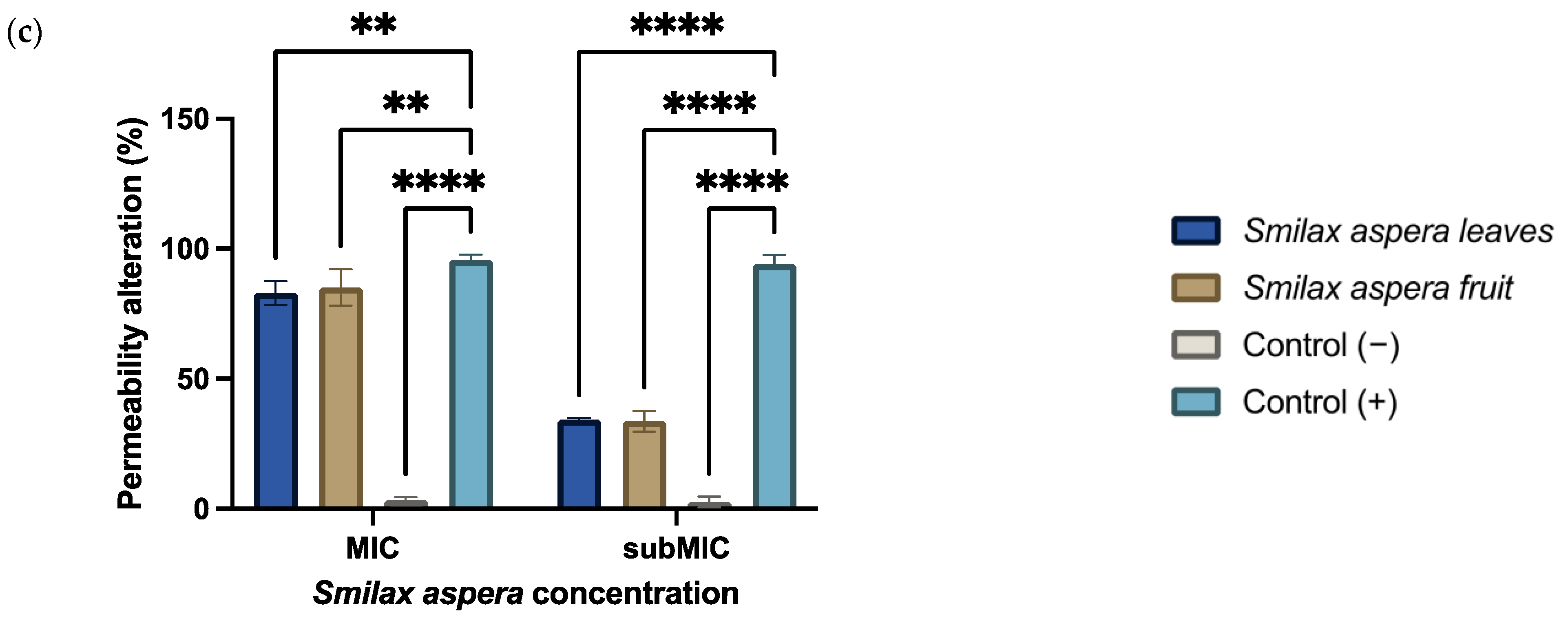
3.3.4. Permeability Alteration
3.4. In Vivo Antibacterial Activity
4. Discussion
4.1. Phytochemical Profile
4.2. On the Antibacterial Activity
4.2.1. Mechanism of Action
4.2.2. Activity of S. aspera and Other Smilax spp. Extracts
4.2.3. Comparison with Synthetic Antimicrobials
4.2.4. Comparison with Other Products for Pear Tree Protection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatta, S.; Joshi, L.R.; Kunwar, R.M.; Bussmann, R.W.; Paniagua-Zambrana, N.Y. Smilax aspera L., Smilax ovalifolia Roxb. Smilacaceae. In Ethnobotany of the Himalayas; Kunwar, R.M., Sher, H., Bussmann, R.W., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 1869–1879. [Google Scholar] [CrossRef]
- Quattrocchi, U. CRC World Dictionary of Medicinal and Poisonous Plants: Common Names, Scientific Names, Eponyms, Synonyms, and Etymology; CRC: Boca Raton, FL, USA, 2012; p. 3960. [Google Scholar]
- Kakouri, E.; Hatziagapiou, K.; Kanakis, C.; Nikola, O.; Lambrou, G.I.; Trigas, P.; Kanaka-Gantenbein, C.; Tarantilis, P.A. Cytotoxic and antioxidant activity of a chemically characterized extract of Smilax aspera leaves and stems. Appl. Sci. 2023, 13, 4784. [Google Scholar] [CrossRef]
- Salas-Coronado, R.; Hernández-Carlos, B.; Llaguno-Guilberto, J.; Santos-Sánchez Norma, F. Phenolic compounds in genus Smilax (Sarsaparilla). In Phenolic Compounds: Natural Sources, Importance, and Applications; Marcos, S.-H., Mariana, P.-T., Maria del Rosario, G.-M., Eds.; IntechOpen: Rijeka, Croatia, 2017; pp. 233–260. [Google Scholar] [CrossRef]
- Longo, L.; Vasapollo, G. Extraction and identification of anthocyanins from Smilax aspera L. berries. Food Chem. 2006, 94, 226–231. [Google Scholar] [CrossRef]
- Delgado-Pelayo, R.; Hornero-Méndez, D. Identification and quantitative analysis of carotenoids and their esters from sarsaparilla (Smilax aspera L.) berries. J. Agric. Food. Chem. 2012, 60, 8225–8232. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.G.; Nunes da Silva, M.; Vasconcelos, M.W.; Carvalho, S.M.P. Scientific and technological advances in the development of sustainable disease management tools: A case study on kiwifruit bacterial canker. Front. Plant Sci. 2024, 14, 1306420. [Google Scholar] [CrossRef]
- Piqué, N.; Miñana-Galbis, D.; Merino, S.; Tomás, J. Virulence factors of Erwinia amylovora: A review. Int. J. Mol. Sci. 2015, 16, 12836–12854. [Google Scholar] [CrossRef]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balestra, G.M.; Spinelli, F. Pseudomonas syringae pv. actinidiae: Ecology, infection dynamics and disease epidemiology. Microb. Ecol. 2020, 80, 81–102. [Google Scholar] [CrossRef]
- Pedroncelli, A.; Puopolo, G. This tree is on fire: A review on the ecology of Erwinia amylovora, the causal agent of fire blight disease. J. Plant Pathol. 2023, in press. [Google Scholar] [CrossRef]
- Vicente, J.G.; Holub, E.B. Xanthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to brassica crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Tarighi, S.; Taheri, P. Effects of plant essential oils on growth and virulence factors of Erwinia amylovora. J. Plant Pathol. 2019, 102, 409–419. [Google Scholar] [CrossRef]
- Wilson, C.; Lukowicz, R.; Merchant, S.; Valquier-Flynn, H.; Caballero, J.; Sandoval, J.; Okuom, M.; Huber, C.; Brooks, T.D.; Wilson, E.; et al. Quantitative and qualitative assessment methods for biofilm growth: A mini-review. Res. Rev. J. Eng. Technol. 2017, 6, 1–25. [Google Scholar]
- Bellemann, P.; Bereswill, S.; Berger, S.; Geider, K. Visualization of capsule formation by Erwinia amylovora and assays to determine amylovoran synthesis. Int. J. Biol. Macromol. 1994, 16, 290–296. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef]
- Langa-Lomba, N.; Sánchez-Hernández, E.; Buzón-Durán, L.; González-García, V.; Casanova-Gascón, J.; Martín-Gil, J.; Martín-Ramos, P. Activity of anthracenediones and flavoring phenols in hydromethanolic extracts of Rubia tinctorum against grapevine phytopathogenic fungi. Plants 2021, 10, 1527. [Google Scholar] [CrossRef]
- Langa-Lomba, N.; Buzón-Durán, L.; Martín-Ramos, P.; Casanova-Gascón, J.; Martín-Gil, J.; Sánchez-Hernández, E.; González-García, V. Assessment of conjugate complexes of chitosan and Urtica dioica or Equisetum arvense extracts for the control of grapevine trunk pathogens. Agronomy 2021, 11, 976. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; González-García, V.; Palacio-Bielsa, A.; Lorenzo-Vidal, B.; Buzón-Durán, L.; Martín-Gil, J.; Martín-Ramos, P. Antibacterial activity of Ginkgo biloba extracts against Clavibacter michiganensis subsp. michiganensis, Pseudomonas spp., and Xanthomonas vesicatoria. Horticulturae 2023, 9, 461. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Li, L.; Ma, W.-H.; Han, T.; Qin, L.-P. Chemical constituents and bioactivities of the liposoluble fraction from different medicinal parts of Crocus sativus. Pharm. Biol. 2011, 49, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, Y.; Han, T.; Zheng, C.; Qin, L. A phytochemical, pharmacological and clinical profile of Paederia foetida and P. scandens. Nat. Prod. Commun. 2014, 9, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; Sánchez-Hernández, E.; Noversa, J.; Cunha, A.; Cortez, I.; Marques, G.; Martín-Ramos, P.; Oliveira, R. Antifungal activity of plant waste extracts against phytopathogenic fungi: Allium sativum peels extract as a promising product targeting the fungal plasma membrane and cell wall. Horticulturae 2023, 9, 136. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Balduque-Gil, J.; Barriuso-Vargas, J.J.; Casanova-Gascón, J.; González-García, V.; Cuchí-Oterino, J.A.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Holm oak (Quercus ilex subsp. ballota (Desf.) Samp.) bark aqueous ammonia extract for the control of invasive forest pathogens. Int. J. Mol. Sci. 2022, 23, 11882. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Balduque-Gil, J.; González-García, V.; Barriuso-Vargas, J.J.; Casanova-Gascón, J.; Martín-Gil, J.; Martín-Ramos, P. Phytochemical profiling of Sambucus nigra L. flower and leaf extracts and their antimicrobial potential against almond tree pathogens. Int. J. Mol. Sci. 2023, 24, 1154. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hernández, E.; González-García, V.; Palacio-Bielsa, A.; Casanova-Gascón, J.; Navas-Gracia, L.M.; Martín-Gil, J.; Martín-Ramos, P. Phytochemical constituents and antimicrobial activity of Euphorbia serrata L. extracts for Borago officinalis L. crop protection. Horticulturae 2023, 9, 652. [Google Scholar] [CrossRef]
- Kundu, C.; Biswas, S.; Thomas, B.S.; Appadoo, D.; Duan, A.; Bhattacharya, S. Evolution of functional group of lignocellulosic biomass and its delignified form during thermal conversion using synchrotron-based THz and laboratory-based in-situ DRIFT spectroscopy. Fuel 2023, 348, 128579. [Google Scholar] [CrossRef]
- Kim, S.; Evans, T.J.; Mukarakate, C.; Bu, L.; Beckham, G.T.; Nimlos, M.R.; Paton, R.S.; Robichaud, D.J. Furan production from glycoaldehyde over HZSM-5. ACS Sustain. Chem. Eng. 2016, 4, 2615–2623. [Google Scholar] [CrossRef]
- Xu, M.; Xue, H.; Li, X.; Zhao, Y.; Lin, L.; Yang, L.; Zheng, G. Chemical composition, antibacterial properties, and mechanism of Smilax china L. polyphenols. Appl. Microbiol. Biotechnol. 2019, 103, 9013–9022. [Google Scholar] [CrossRef]
- Islam, M.; ArİFuzzaman, A.; Rahman, M.; Rahman, M.A.; Kawsar, S.M.A.K. Novel methyl 4,6-O-benzylidene-a-D-glucopyranoside derivatives: Synthesis, structural characterization and evaluation of antibacterial activities. Hacettepe J. Biol. Chem. 2019, 47, 153–164. [Google Scholar] [CrossRef]
- Kawsar, S.M.A. Regioselective synthesis, characterization, and antimicrobial activities of some new monosaccharide derivatives. Sci. Pharm. 2014, 82, 1–20. [Google Scholar] [CrossRef]
- Misbah, M.M.; Ferdous, J.; Bulbul, M.Z.; Chowdhury, T.S.; Dey, S.; Hasan, I.; Kawsar, S.M. Evaluation of MIC, MBC, MFC and anticancer activities of acylated methyl β-D-galactopyranoside esters. Int. J. Biosci. 2020, 16, 299–309. [Google Scholar]
- Hosen, M.A.; El Bakri, Y.; Rehman, H.M.; Hashem, H.E.; Saki, M.; Kawsar, S.M.A. A computational investigation of galactopyranoside esters as antimicrobial agents through antiviral, molecular docking, molecular dynamics, pharmacokinetics, and bioactivity prediction. J. Biomol. Struct. Dyn. 2023, 42, 1015–1030. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, F.; Islam, A.U.; Mukhrish, Y.E.; Bakri, Y.E.; Ahmad, S.; Ozeki, Y.; Kawsar, S.M.A. Efficient antibacterial/antifungal activities: Synthesis, molecular docking, molecular dynamics, pharmacokinetic, and binding free energy of galactopyranoside derivatives. Molecules 2022, 28, 219. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Feng, L.; Zhang, Y.; Zhang, R.; Wang, G.; Zhang, Y. Effect of juglone against Pseudomonas syringae pv actinidiae planktonic growth and biofilm formation. Molecules 2021, 26, 7580. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, R.; Dahal, B.; Gautam, R.; Shrestha, S.; Joshi, S.; Luitel, A.; Khanal, D. Phytochemical studies traditional medicinal plants of Nepal and their formulations. Int. J. Biol. Pharm. Allied Sci. 2014, 3, 189–203. [Google Scholar]
- Mohammad Sawalha, M.K. Phytochemical screening of wild Ephedra foeminea and Smilax aspera fruits and their in-vitro pharmacological activities. M.Sc. Thesis, Al-Quds University, Jerusalem, Israel, 2018. [Google Scholar]
- Abbasołu, U.; Türköz, S. Antimicrobial activities of saponin extracts from some indigenous plants of Turkey. Int. J. Pharmacogn. 2008, 33, 293–296. [Google Scholar] [CrossRef]
- Belhouchet, Z.; Sautour, M.; Miyamoto, T.; Lacaille-Dubois, M.-A. Steroidal saponins from the roots of Smilax aspera subsp. mauritanica. Chem. Pharm. Bull. 2008, 56, 1324–1327. [Google Scholar] [CrossRef] [PubMed]
- Sautour, M.; Miyamoto, T.; Lacaille-Dubois, M.-A. Steroidal saponins from Smilax medica and their antifungal activity. J. Nat. Prod. 2005, 68, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.-W.; Zhang, Z.; Long, H.-L.; Zhang, Y.-J. Steroidal saponins from the genus Smilax and their biological activities. Nat. Prod. Bioprospect. 2017, 7, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Konopatzki Hirota, B.C.; da Silva Paula de Oliveira, C.; Merino, F.; dos Santos Verdam, M.; Da Silva, C.; Murakami, F.; Miguel, O.; Miguel, M. Larvicide and antifungal activities of sarsaparilla (Smilax larvata) extracts. Int. J. Pharm. Pharm. Sci. 2015, 7, 308–311. [Google Scholar]
- Seo, H.-K.; Lee, J.-H.; Kim, H.-S.; Lee, C.-K.; Lee, S.-C. Antioxidant and antimicrobial activities of Smilax china L. leaf extracts. Food Sci. Biotechnol. 2012, 21, 1723–1727. [Google Scholar] [CrossRef]
- Jeeno, P.; Tongban, S.; Yana, P.; Wongta, A.; Sutan, K.; Yadoung, S.; Hongsibsong, S. Tentative identification of phytochemicals from Smilax glabra and Smilax corbularia extracts by LC-QTOF/MS and their bioactive potential. Plants 2022, 11, 2089. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Rizwan, K.; Rashid, U.; Saeed, R.; Saeed, A.A.; Rasool, N.; Riaz, M. GC/MS profiling, in vitro antioxidant, antimicrobial and haemolytic activities of Smilax macrophylla leaves. Arab. J. Chem. 2017, 10, S1460–S1468. [Google Scholar] [CrossRef]
- Borkataky, M. Antimicrobial and antioxidant activity of Smilax perfoliata Lour. Der. Pharm. Lett. 2014, 6, 246–250. [Google Scholar]
- Morais, M.I.; Pinto, M.E.A.; Araújo, S.G.; Castro, A.H.F.; Duarte-Almeida, J.M.; Rosa, L.H.; Rosa, C.A.; Johann, S.; Lima, L.A.R.d.S. Antioxidant and antifungal activities of Smilax campestris Griseb. (Smilacaceae). Nat. Prod. Res. 2014, 28, 1275–1279. [Google Scholar] [CrossRef]
- Dhanya Shree, V.S.; Arbin, A.; Saema Noorain, G.K.; Sahana, B.K.; Prashith Kekuda, T.R. Preliminary phytochemical analysis, antimicrobial and antioxidant activity of Smilax zeylanica L. (Smilacaceae). J. Drug Deliv. Ther. 2018, 8, 237–243. [Google Scholar] [CrossRef]
- Rajbhandari, M.; Paneru, D.P. Phytochemical analysis and antimicrobial activity of Smilax ovalifolia Roxb. Ex D. Don. Nepal J. Sci. Technol. 2020, 19, 89–96. [Google Scholar] [CrossRef]
- Hamid, A.; Aiyelaagbe, O. The screening of phytoconstituents, antibacterial and antifungal properties of Smilax kraussiana leaves. Pharm. Sin. 2011, 2, 267–273. [Google Scholar]
- Joo, J.-H.; Han, M.-H.; Kim, J.-I.; Kim, J.-E.; Jung, K.-H.; Oh, H.S.; Chung, Y.S.; An, H.J.; Lee, J.D.; Moon, G.-S.; et al. Antimicrobial activity of Smilax china L. root extracts against the acne-causing bacterium, Cutibacterium acnes, and its active compounds. Molecules 2022, 27, 8331. [Google Scholar] [CrossRef]
- Xu, S.; Shang, M.-Y.; Liu, G.-X.; Xu, F.; Wang, X.; Shou, C.-C.; Cai, S.-Q. Chemical constituents from the rhizomes of Smilax glabra and their antimicrobial activity. Molecules 2013, 18, 5265–5287. [Google Scholar] [CrossRef] [PubMed]
- McMurray, R.L.; Ball, M.E.E.; Tunney, M.M.; Corcionivoschi, N.; Situ, C. Antibacterial activity of four plant extracts extracted from traditional chinese medicinal plants against Listeria monocytogenes, Escherichia coli, and Salmonella enterica subsp. enterica serovar Enteritidis. Microorganisms 2020, 8, 962. [Google Scholar] [CrossRef]
- Cáceres, A.; Cruz, S.M.; Martínez, V.; Gaitán, I.; Santizo, A.; Gattuso, S.; Gattuso, M. Ethnobotanical, pharmacognostical, pharmacological and phytochemical studies on Smilax domingensis in Guatemala. Rev. Bras. Farmacogn. 2012, 22, 239–248. [Google Scholar] [CrossRef]
- Jimenez Madrid, A.M.; Ivey, M.L.L. An overview of streptomycin resistance in Erwinia amylovora from Ohio apple orchards. Plant Health Prog. 2023, 24, 56–61. [Google Scholar] [CrossRef]
- Förster, H.; McGhee, G.C.; Sundin, G.W.; Adaskaveg, J.E. Characterization of streptomycin resistance in isolates of Erwinia amylovora in California. Phytopathology 2015, 105, 1302–1310. [Google Scholar] [CrossRef]
- Cameron, A.; Sarojini, V. Pseudomonas syringae pv. actinidiae: Chemical control, resistance mechanisms and possible alternatives. Plant Pathol. 2013, 63, 1–11. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, J.-H.; Kim, G.-H.; Jung, J.-S.; Hur, J.-S.; Koh, Y.-J. Comparative analysis of Korean and Japanese strains of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit. Plant Pathol. J. 2005, 21, 119–126. [Google Scholar] [CrossRef]
- Chang, C.; Donaldson, R.; Crowley, M.; Pinnow, D. A new semiselective medium for the isolation of Xanthomonas campestris pv. campestris from crucifer seeds. Phytopathology 1991, 81, 449–453. [Google Scholar] [CrossRef]
- Yemata, G.; Fetene, M. In vitro antibacterial activity of some antibiotics against Xanthomonas campestris pv. musacearum: Ensete ventricosum (Welw.) Cheesman pathogen. J. Phytopathol. Pest Manag. 2017, 4, 48–57. [Google Scholar]
- Kim, J.Y.; Kim, J.Y.; Park, J.Y.; Kim, J.-S.; Seo, M.-K.; Shin, M.-K.; Kim, J.-H. Synergistic bactericidal effects of carvone and β-lactams against Xanthomonas campestris pv. vesicatoria. Appl. Biol. Chem. 2023, 66, 44. [Google Scholar] [CrossRef]
- Kavitha, K.S.; Satish, S. Evaluation of antimicrobial and antioxidant activities from Toona ciliata Roemer. J. Anal. Sci. Technol. 2013, 4, 23. [Google Scholar] [CrossRef]
- Wang, M.; Bai, Q.-X.; Zheng, X.-X.; Hu, W.-J.; Wang, S.; Tang, H.-P.; Yu, A.-Q.; Yang, B.-Y.; Kuang, H.-X. Smilax china L.: A review of its botany, ethnopharmacology, phytochemistry, pharmacological activities, actual and potential applications. J. Ethnopharmacol. 2024, 318, 116992. [Google Scholar] [CrossRef]
- Bastas, K.K. Effects of some plant extracts against Erwinia amylovora by the induced resistance on apple in vitro and in vivo conditions. In Proceedings of the 6th International Conference on Sustainable Agriculture and Environment (ICSAE-6), Konya, Turkey, 3–5 October 2019; pp. 504–509. [Google Scholar]
- Fontana, R.; Macchi, G.; Caproni, A.; Sicurella, M.; Buratto, M.; Salvatori, F.; Pappadà, M.; Manfredini, S.; Baldisserotto, A.; Marconi, P. Control of Erwinia amylovora growth by Moringa oleifera leaf extracts: In vitro and in planta effects. Plants 2022, 11, 957. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf | Fruit | Assignment |
|---|---|---|
| 3354 | 3378 | O–H stretching of alcohols (e.g., 2,3-butanediol) and phenols |
| 2916 | 2915 | C–H stretching of alkanes, methyl, and methylene groups |
| 2848 | 2849 | C–H stretching of aldehydes |
| 1732 | 1716 | C=O stretching of ketones (e.g., 1-hydroxy-2-propanone and 2-hydroxy-2-cyclopenten-1-one) and aldehydes |
| 1640 | 1640 | C=C stretching of alkenes and aromatic compounds |
| 1516 | 1521 | C=C stretching and C=C–H bending vibrations of the aromatic ring (e.g., in catechol) |
| 1472 | 1465 | C–H bending of alkanes, methyl and methylene groups, and aldehydes |
| 1377 | 1376 | C–H bending of alkanes and methyl groups |
| 1242 | 1246 | C–O stretching vibration in esters |
| 1161 | 1164 | C–O–C stretching mode |
| 1066 | 1066 | N–O stretching frequencies in oximes |
| 890 | C–H bending in alkenes; C–H out-of-plane deformation of substituted benzene rings |
| Pathogen | Streptomycin | Tetracycline | Smilax aspera Leaf Extract | Smilax aspera Fruit Extract |
|---|---|---|---|---|
| Erwinia amylovora | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.014 ± 0.002 | 0.018 ± 0.003 |
| Pseudomonas syringae pv. actinidiae | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | 0.003 ± 0.001 |
| Xanthomonas campestris pv. campestris | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.001 | 0.001 ± 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontana, R.; Sánchez-Hernández, E.; Martín-Ramos, P.; Martín-Gil, J.; Marconi, P. Smilax aspera L. Leaf and Fruit Extracts as Antibacterial Agents for Crop Protection. Agronomy 2024, 14, 383. https://doi.org/10.3390/agronomy14020383
Fontana R, Sánchez-Hernández E, Martín-Ramos P, Martín-Gil J, Marconi P. Smilax aspera L. Leaf and Fruit Extracts as Antibacterial Agents for Crop Protection. Agronomy. 2024; 14(2):383. https://doi.org/10.3390/agronomy14020383
Chicago/Turabian StyleFontana, Riccardo, Eva Sánchez-Hernández, Pablo Martín-Ramos, Jesús Martín-Gil, and Peggy Marconi. 2024. "Smilax aspera L. Leaf and Fruit Extracts as Antibacterial Agents for Crop Protection" Agronomy 14, no. 2: 383. https://doi.org/10.3390/agronomy14020383