


Integrated Management of Clubroot in Zhejiang Province, China
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of the Pathotype of P. brassicae Using the ECD and Williams Systems
2.3. Chemical Treatment in Greenhouse Experiment
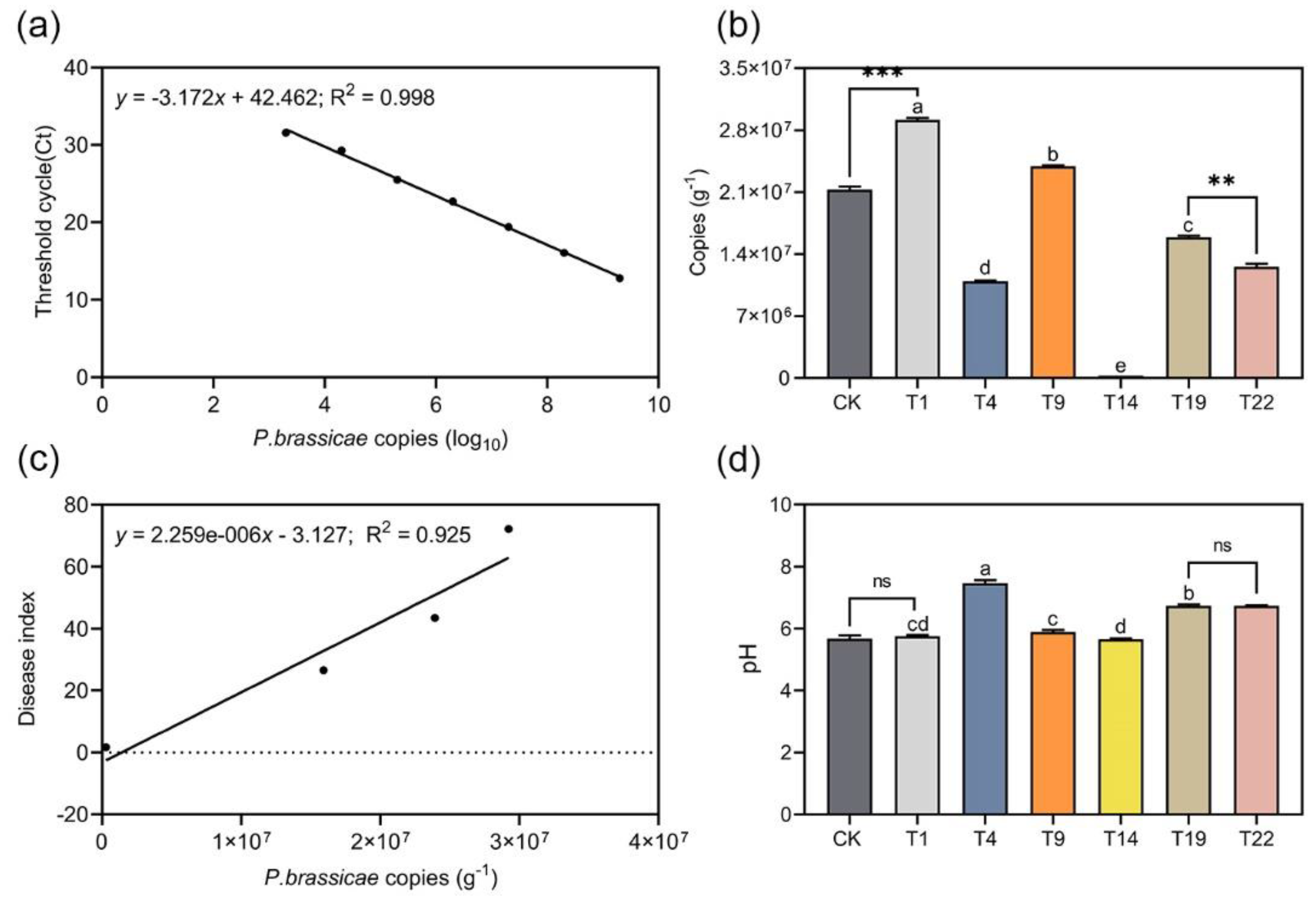
2.4. Quantitative Polymerase Chain Reaction (qPCR) Analysis of Infected Soil
2.5. Soil pH Determination
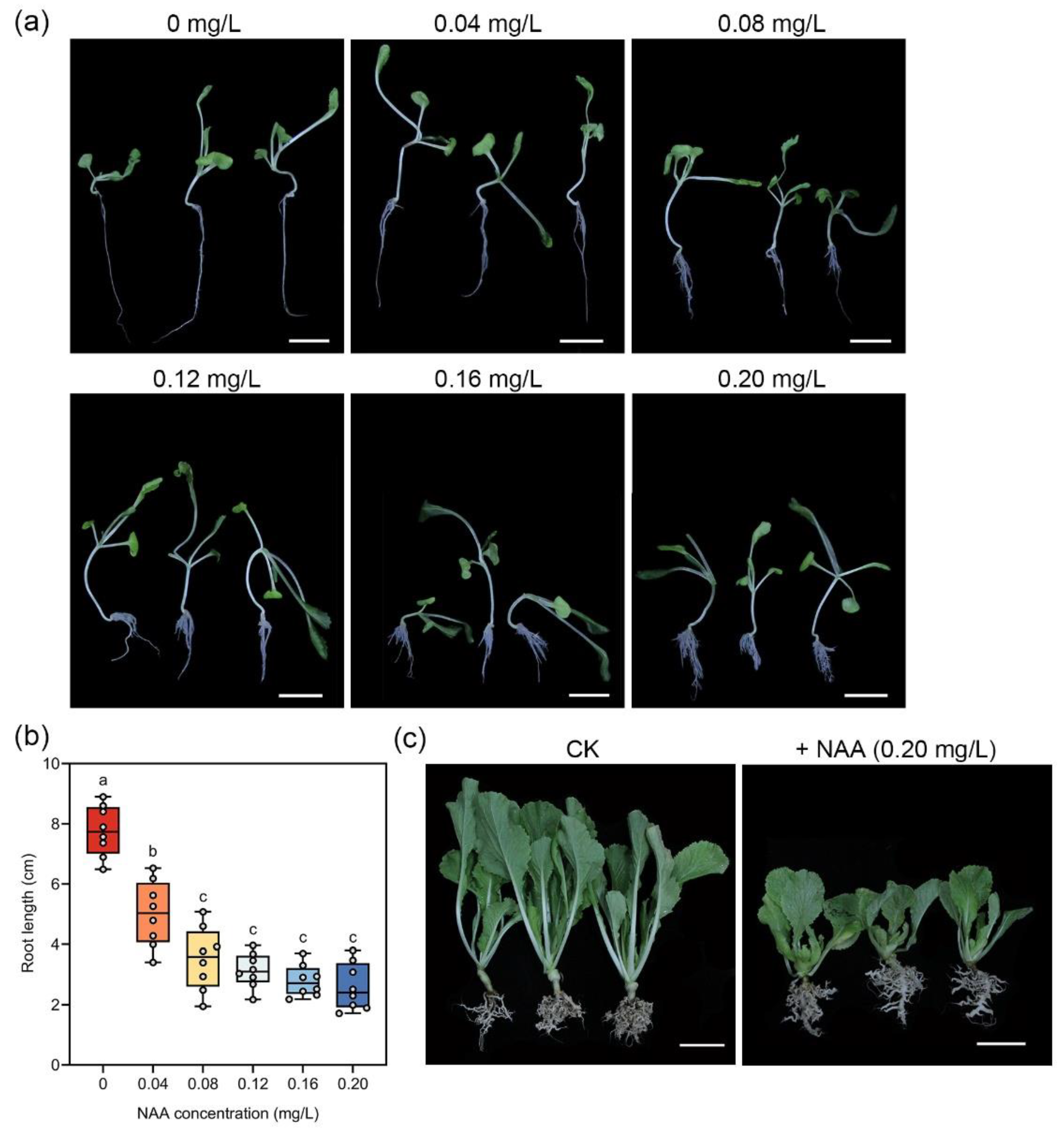
2.6. NAA Treatment
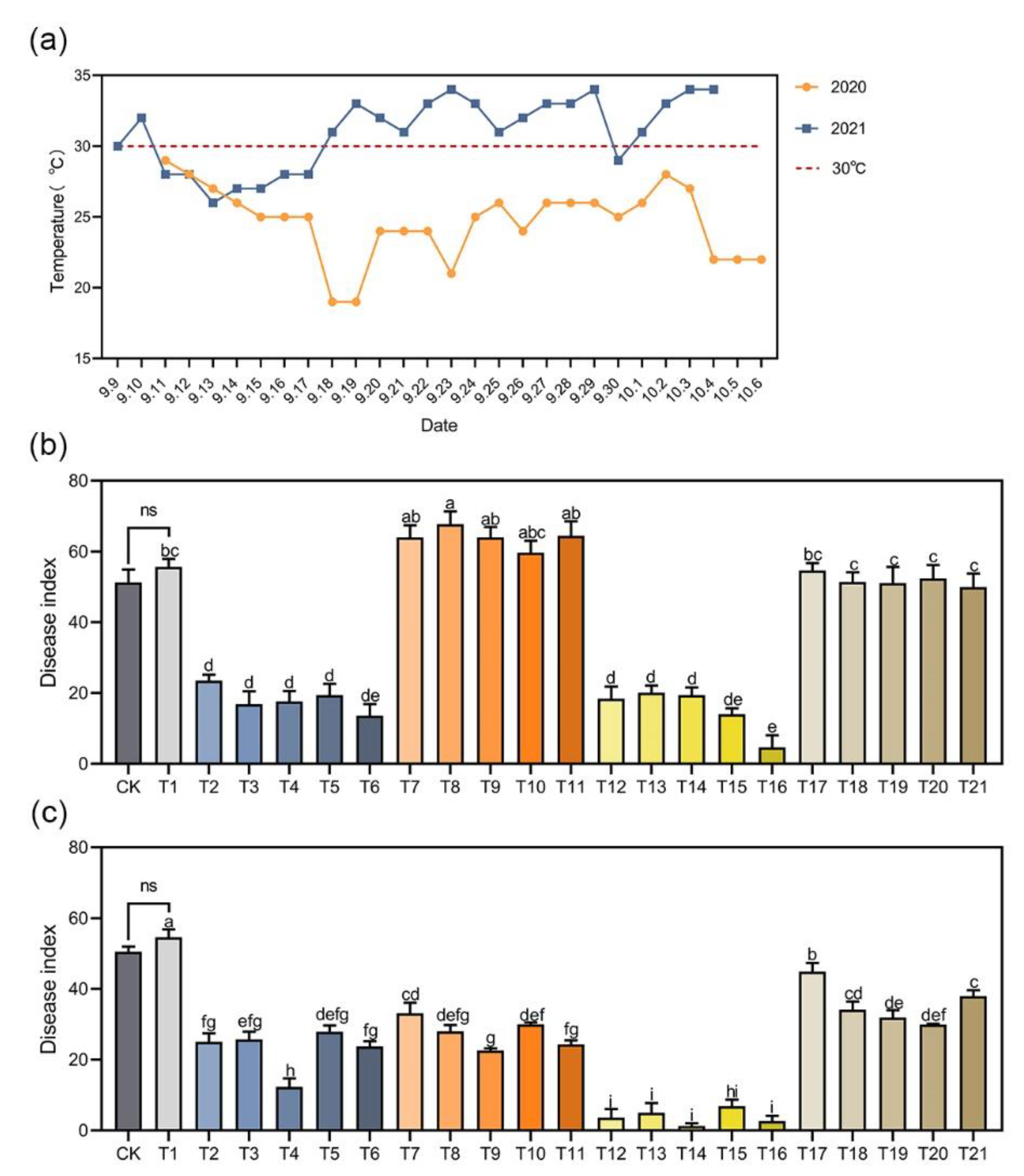
2.7. Field Trials
2.8. Statistical Analysis
3. Results
3.1. Identification of the Pathotypes of P. brassicae with the Williams and ECD Systems
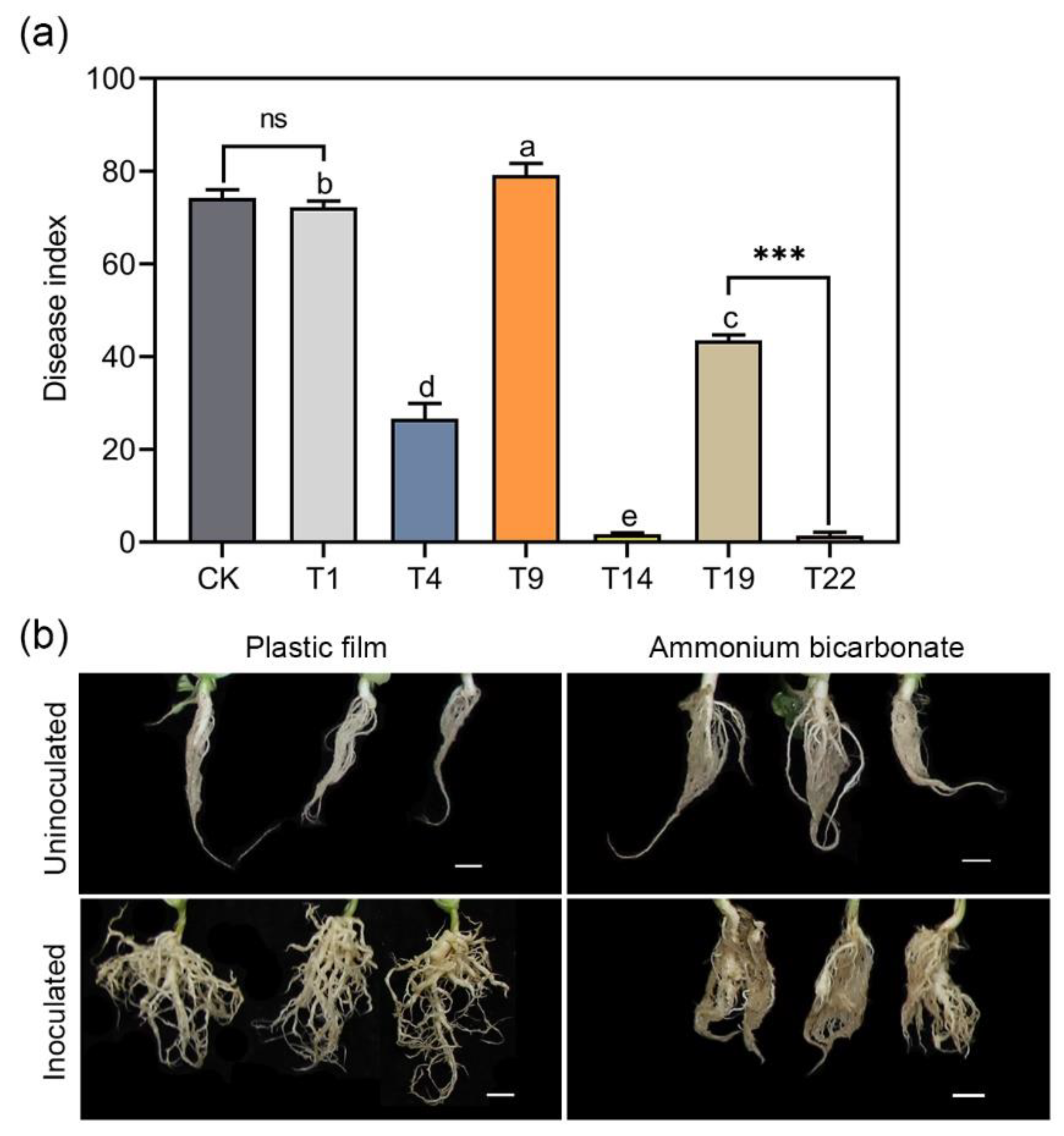
3.2. Effects of Different Chemical Treatments on Clubroot Control in the Greenhouse Experiments
3.3. The Resting Spore Density in the Soil and the Soil pH in Greenhouse Experiments
3.4. The Effect of NAA on ‘Granaat’ Root System and Clubroot Control
3.5. Effects of Different Chemical Treatments on Clubroot Control in the Field Trials
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dixon, G.R. Clubroot (Plasmodiophora brassicae Woronin)—An agricultural and biological challenge worldwide. Can. J. Plant Pathol. 2014, 36, 5–18. [Google Scholar] [CrossRef]
- Hasan, J.; Megha, S.; Rahman, H. Clubroot in Brassica: Recent advances in genomics, breeding, and disease management. Genome 2021, 64, 735–760. [Google Scholar] [CrossRef]
- Aist, J.R.; Williams, P.H. Cytology and kinetics of cabbage root hair penetration by Plasmodiophora-brassicae. Can. J. Bot. 1971, 49, 2023–2034. [Google Scholar] [CrossRef]
- Liu, L.J.; Qin, L.; Zhou, Z.Q.; Hendriks, W.; Liu, S.Y.; Wei, Y.D. Refining the life cycle of Plasmodiophora brassicae. Phytopathology 2020, 110, 1704–1712. [Google Scholar] [CrossRef]
- Zhu, B.; Liang, Z.L.; Zang, Y.X.; Zhu, Z.J.; Yang, J. Diversity of glucosinolates among common Brassicaceae vegetables in China. Hortic. Plant J. 2023, 9, 365–380. [Google Scholar] [CrossRef]
- Huang, Q.W.; Ouyang, L.; Wang, Y.J. Investigation and control of clubroot for cruciferious crops in Jiangxi. Plant Protec. Commun. 1955, 8, 142–153. [Google Scholar]
- Zhu, G.N. Clubroot of cruciferous vegetables. Sci. Agric. Sin. 1956, 11, 677–678. [Google Scholar]
- Chai, A.L.; Xie, X.W.; Shi, Y.X.; Li, B.J. Research status of clubroot (Plasmodiophora brassicae) on cruciferous crops in China. Can. J. Plant Pathol. 2014, 36, 142–153. [Google Scholar] [CrossRef]
- Macfarlane, I. Variation in Plasmodiophora brassicae woron. Ann. Appl. Biol. 1955, 43, 297–306. [Google Scholar] [CrossRef]
- Tso, H.H.; Galindo-González, L.; Strelkov, S.E. Current and future pathotyping platforms for Plasmodiophora brassicae in Canada. Plants-Basel 2021, 10, 1446. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.H. A system for determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga. Phytopathology 1966, 56, 624–626. [Google Scholar]
- Buczacki, S.T.; Toxopeus, H.; Mattusch, P.; Johnston, T.D.; Dixon, G.R.; Hobolth, L.A. Study of physiologic specialization in Plasmodiophora-brassicae—Proposals for attempted rationalization through an international approach. Trans. Br. Mycol. Soc. 1975, 65, 295–303. [Google Scholar] [CrossRef]
- Pang, W.X.; Liang, Y.; Zhan, Z.X.; Li, X.N.; Piao, Z.Y. Development of a sinitic clubroot differential set for the pathotype classification of Plasmodiophora brassicae. Front. Plant. Sci. 2020, 11, 568771. [Google Scholar] [CrossRef] [PubMed]
- Schwelm, A.; Ludwig-Müller, J. Molecular pathotyping of Plasmodiophora brassicae-genomes, marker genes, and obstacles. Pathogens 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.R.; Gossen, B.D.; Robson, J.; Al-Daoud, F. Interaction of solarization, fumigation, and totally impermeable film for the management of clubroot (Plasmodiophora brassicae) on brassica crops. In Proceedings of the 9th International Symposium on Soil and Substrate Disinfestation, Heraklion, Greece, 9–13 September 2018; pp. 153–159. [Google Scholar]
- Diederichsen, E.; Frauen, M.; Ludwig-Müller, J. Special Issue: Clubroot disease management challenges from a German perspective. Can. J. Plant Pathol. 2014, 36, 85–98. [Google Scholar] [CrossRef]
- Fredua-Agyeman, R.; Rahman, H. Mapping of the clubroot disease resistance in spring Brassica napus canola introgressed from European winter canola cv. ‘Mendel’. Euphytica 2016, 211, 201–213. [Google Scholar] [CrossRef]
- Fredua-Agyeman, R.; Hwang, S.F.; Strelkov, S.E.; Zhou, Q.; Feindel, D. Potential loss of clubroot resistance genes from donor parent Brassica rapa subsp. rapifera (ECD 04) during doubled haploid production. Plant Pathol. 2018, 67, 892–901. [Google Scholar] [CrossRef]
- Peng, G.; McGregor, L.; Lahlali, R.; Gossen, B.D.; Hwang, S.F.; Adhikari, K.K.; Strelkov, S.E.; McDonald, M.R. Potential biological control of clubroot on canola and crucifer vegetable crops. Plant Pathol. 2011, 60, 566–574. [Google Scholar] [CrossRef]
- Liu, C.M.; Yang, Z.F.; He, P.F.; Munir, S.; He, P.B.; Wu, Y.X.; Ho, H.H.; He, Y.Q. Fluazinam positively affected the microbial communities in clubroot cabbage rhizosphere. Sci. Hortic. 2019, 256, 108519. [Google Scholar] [CrossRef]
- Naiki, T.; Dixon, G.R. The effects of chemicals on developmental stages of Plasmodiophora-brassicae (clubroot). Plant Pathol. 1987, 36, 316–327. [Google Scholar] [CrossRef]
- Wang, Q.X.; Yan, D.D.; Wang, X.L.; Lu, P.X.; Li, X.Y.; Cao, A.C. Research advances in soil fumigants. Acta Phytophy. Sin. 2017, 44, 529–543. [Google Scholar]
- Hwang, S.F.; Ahmed, H.U.; Strelkov, S.E.; Zhou, Q.; Gossen, B.D.; McDonald, M.R.; Peng, G.; Turnbull, G.D. Suppression of clubroot by dazomet fumigant. Can. J. Plant Sci. 2018, 98, 172–182. [Google Scholar] [CrossRef]
- Arslan, U.; Ilhan, K.; Karabulut, O.A. Evaluation of the use of ammonium bicarbonate and oregano (Origanum vulgare ssp. hirtum) extract on the control of apple scab. J. Phytopathol. 2013, 161, 382–388. [Google Scholar] [CrossRef]
- Wang, J.Z. A study on the soil treatment efficacy of 50% soluble powder of chloroisobromine cyanuric acid in controlling soil-borne crop diseases. Mod. Agric. Sci. Tech. 2019, 09, 86–87. [Google Scholar]
- Li, D. The research on the efficacy of 50% chloroisobromine cyanuric acid in controlling basal stem rot, root rot, and wilt disease in tomatoes. Jiangxi Agric. 2019, 22, 35. [Google Scholar] [CrossRef]
- Meier, M.; Liu, Y.; Lay-Pruitt, K.S.; Takahashi, H.; Von Wirén, N. Auxin-mediated root branching is determined by the form of available nitrogen. Nat. Plants 2020, 6, 1136–1145. [Google Scholar] [CrossRef]
- Yang, Y.F. Effects of Naphthalene Acetic Acid on the Growth and Development of Arabidopsis thaliana. Master’s Thesis, Zhengzhou University, Zhengzhou, China, 2017. [Google Scholar]
- Liu, X.T.; Strelkov, S.E.; Sun, R.F.; Hwang, S.F.; Fredua-Agyeman, R.; Li, F.; Zhang, S.F.; Li, G.L.; Zhang, S.J.; Zhang, H. Histopathology of the Plasmodiophora brassicae-Chinese cabbage interaction in hosts carrying different sources of resistance. Front. Plant. Sci. 2022, 12, 783550. [Google Scholar] [CrossRef] [PubMed]
- Kuginuki, Y.; Yoshikawa, H.; Hirai, M. Variation in virulence of Plasmodiophora brassicae in Japan tested with clubroot-resistant cultivars of Chinese cabbage (Brassica rapa L. ssp pekinensis). Eur. J. Plant Pathol. 1999, 105, 327–332. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Tewari, J.P.; Smith-Degenhardt, E. Characterization of Plasmodiophora brassicae populations from Alberta, Canada. Can. J. Plant Pathol. 2006, 28, 467–474. [Google Scholar] [CrossRef]
- Czubatka-Bienkowska, A.; Kaczmarek, J.; Marzec-Schmidt, K.; Nierobca, A.; Czajka, A.; Jedryczka, M. Country-wide qPCR based assessment of Plasmodiophora brassicae spread in agricultural soils and recommendations for the cultivation of Brassicaceae crops in Poland. Pathogens 2020, 9, 1070. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.D.; He, Q.W.; Mu, J.H.; Kang, J.; Wang, P.W. Study on the tissue culture and plant regeneration of Chinese cabbage. J. Jilin Agric. Univ. 2005, 27, 391–395. [Google Scholar]
- Dixon, G.R. Managing clubroot disease (caused by Plasmodiophora brassicae Wor.) by exploiting the interactions between calcium cyanamide fertilizer and soil microorganisms. J. Agric. Sci. 2017, 155, 527–543. [Google Scholar] [CrossRef]
- Niwa, R.; Nomura, Y.; Osaki, M.; Ezawa, T. Suppression of clubroot disease under neutral pH caused by inhibition of spore germination of Plasmodiophora brassicae in the rhizosphere. Plant Pathol. 2008, 57, 445–452. [Google Scholar] [CrossRef]
- Myers, D.F.; Campbell, R.N. Lime and the control of clubroot of crucifers—Effects of pH, calcium, magnesium, and their interactions. Phytopathology 1985, 75, 670–673. [Google Scholar] [CrossRef]
- Fletcher, J.T.; Hims, M.J.; Archer, F.C.; Brown, A. Effects of adding calcium and sodium salts to field soils on the incidence of clubroot. Ann. Appl. Biol. 1982, 100, 245–251. [Google Scholar] [CrossRef]
- Hu, H.T.; Zhu, Z.G.; Jiao, Z.J.; Min, Y.; Cao, C.X.; Yan, D.D.; Cao, A.C. Effects of soil disinfection by dazomet on control efficacy of cabbage clubroot disease and soil fungal communities in high mountain area. J. Huazhong Agric. Univ. 2019, 38, 25–31. [Google Scholar]
- Donald, E.C.; Porter, I.J.; Faggian, R.; Lancaster, R.A. An integrated approach to the control of clubroot in vegetable Brassica crops. In Proceedings of the Joint Meeting of the 14th Crucifer Genetics Workshop/4th Symposium on Brassicas, Chungnam National University, Daejeon, Republic of Korea, 24–28 October 2004; pp. 283–300. [Google Scholar]
- Belec, C.; Tremblay, N.; Coulombe, J. Liming and calcium cyanamid for clubroot control in cauliflower. In Proceedings of the 26th International Horticultural Congress, Toronto, Canada, 11–17 August 2002; pp. 41–46. [Google Scholar]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Fang, W.S.; Wang, Q.X.; Yan, D.D.; Li, Y.; Cao, B.W.; Xu, J.; Jin, X.; Cao, A.C. Research progresses and future development trends of soil fumigant dazomet in control of soil-borne diseases. Acta Phytophy. Sin. 2023, 50, 40–49. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment 1 | Common Name | Formulation 2 | Rate (a.i. m−2) 3 | Registrant Information |
|---|---|---|---|---|
| CK | Without treatment | - | - | - |
| T1 | Black plastic film | - | - | Jiangsu Taika Greenhouse Co., Ltd. (Suqian City, Jiangsu Province, China) |
| T2 | Calcium cyanamide (chemical formula: CaCN2) | GR | 60 g | Ningxia Jiafeng Chemicals Co., Ltd. (Shizuishan City, Ningxia Hui Autonomous Region, China) |
| T3 | N ≥ 21% | 75 g | ||
| T4 | CaCN2 ≥ 55% | 90 g | ||
| T5 | 105 g | |||
| T6 | 120 g | |||
| T7 | Chloroisobromine cyanuric acid (chemical formula: C3HO3N3ClBr) | SP | 1.5 g | Hebei Shangrui Chemicals Co., Ltd. (Shijiazhuang City, Hebei Province, China) |
| T8 | C3HO3N3ClBr = 50% | 3.0 g | ||
| T9 | 4.5 g | |||
| T10 | 6.0 g | |||
| T11 | 7.5 g | |||
| T12 | Dazomet (chemical formula: C5H10N2S2) | MG | 27.5 g | Zhejiang Haizheng Chemicals Co., Ltd. (Taizhou City, Zhejiang Province, China) |
| T13 | C5H10N2S2 = 98% | 32.5 g | ||
| T14 | 37.5 g | |||
| T15 | 42.5 g | |||
| T16 | 47.5 g | |||
| T17 | Ammonium bicarbonate (chemical formula: NH4HCO3) | - | 120 g | Weihui Yubei Chemicals Co., Ltd. (Xinxiang City, Henan Province, China) |
| T18 | N ≥ 17.1% | 135 g | ||
| T19 | H2O ≤ 3.5% | 150 g | ||
| T20 | 165 g | |||
| T21 | 180 g | |||
| T22 | 150 g 4 |
| Sampling Location | Host Origin | Pathotype | |
|---|---|---|---|
| Williams System | ECD System | ||
| Hangzhou, Yuhang | Purple cai-tai | P1 | ECD20/31/12 |
| Quzhou, Kaihua | Canola | P3 | ECD20/15/4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yu, H.; Zhao, T.; Hussain, I.; Ma, X.; Wang, Y.; Liu, K.; Sun, N.; Yu, X. Integrated Management of Clubroot in Zhejiang Province, China. Agronomy 2024, 14, 377. https://doi.org/10.3390/agronomy14020377
Zhang Y, Yu H, Zhao T, Hussain I, Ma X, Wang Y, Liu K, Sun N, Yu X. Integrated Management of Clubroot in Zhejiang Province, China. Agronomy. 2024; 14(2):377. https://doi.org/10.3390/agronomy14020377
Chicago/Turabian StyleZhang, Yi, Hongrui Yu, Tong Zhao, Iqbal Hussain, Xinyan Ma, Yuqi Wang, Kaiwen Liu, Nairan Sun, and Xiaolin Yu. 2024. "Integrated Management of Clubroot in Zhejiang Province, China" Agronomy 14, no. 2: 377. https://doi.org/10.3390/agronomy14020377