
Evaluation of Biochar and Inorganic Fertilizer on Soil Available Phosphorus and Bacterial Community Dynamics in Acidic Paddy Soils for Different Incubation Temperatures
 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Descriptions and Soil Sampling
2.2. Biochar Production and Characterization
2.3. Incubation Experiment
Treatments
2.4. Determination of Soil pH, Available P, and Phosphatase Enzyme Activity
2.5. Illumina MiSeq Sequencing
2.6. High-Throughput Sequencing Data Processing
2.7. Statistical Analysis
3. Results
3.1. Variations in the Soil pH, Available P Concentration, and Acid Phosphatase Activity
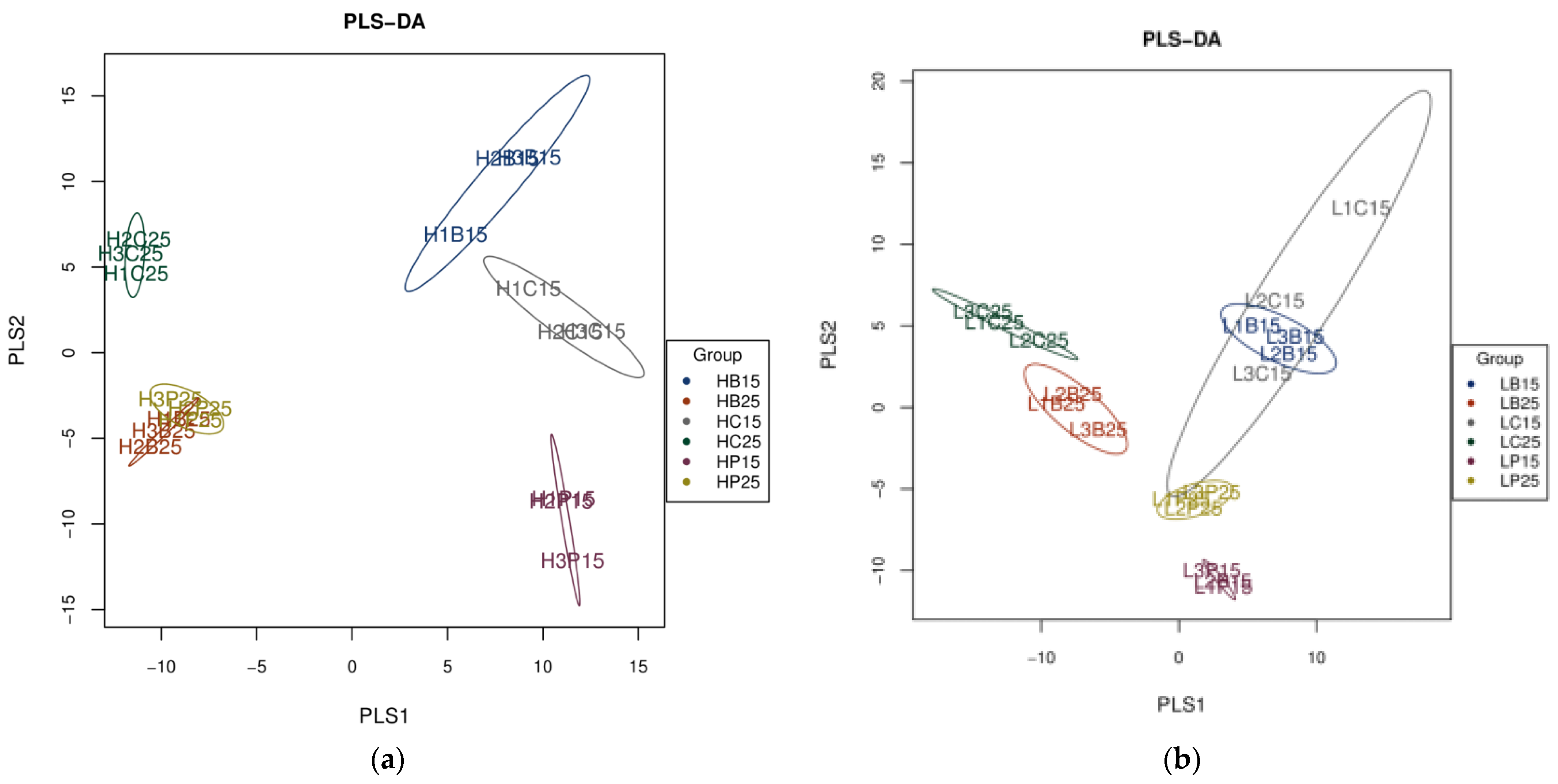
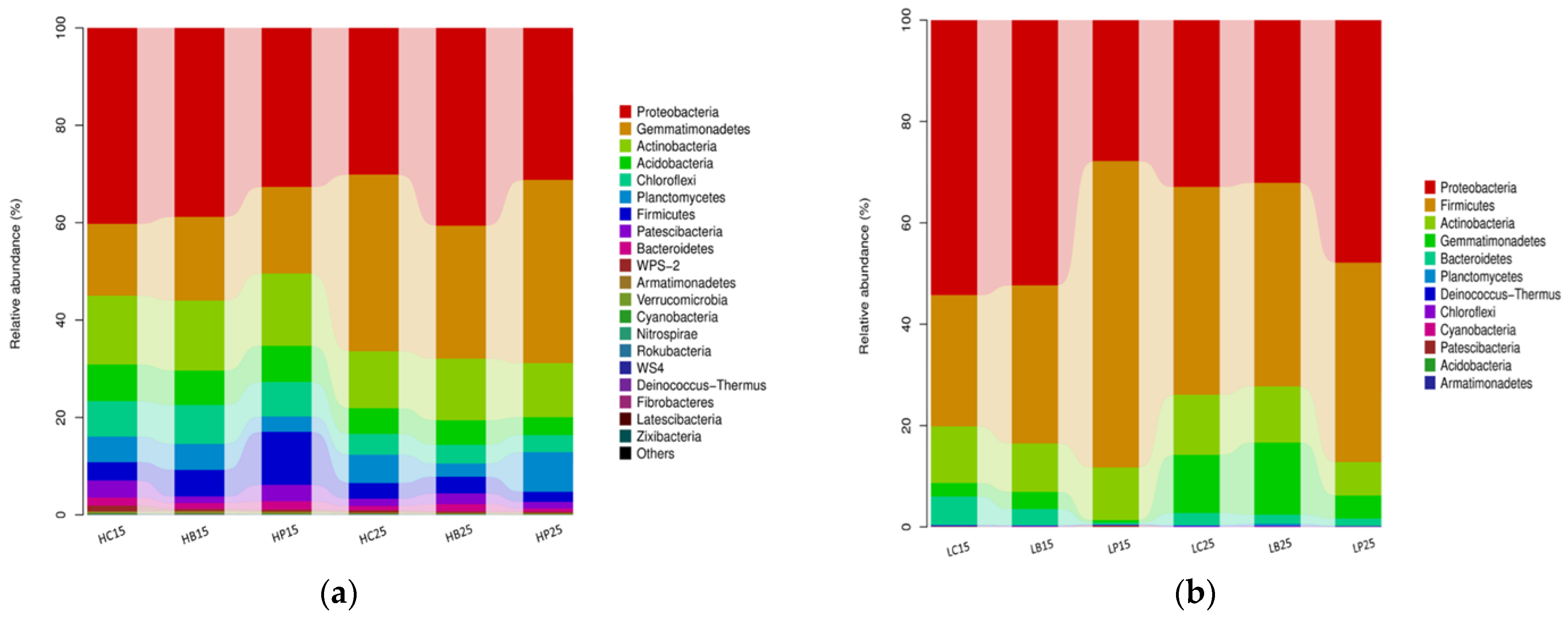
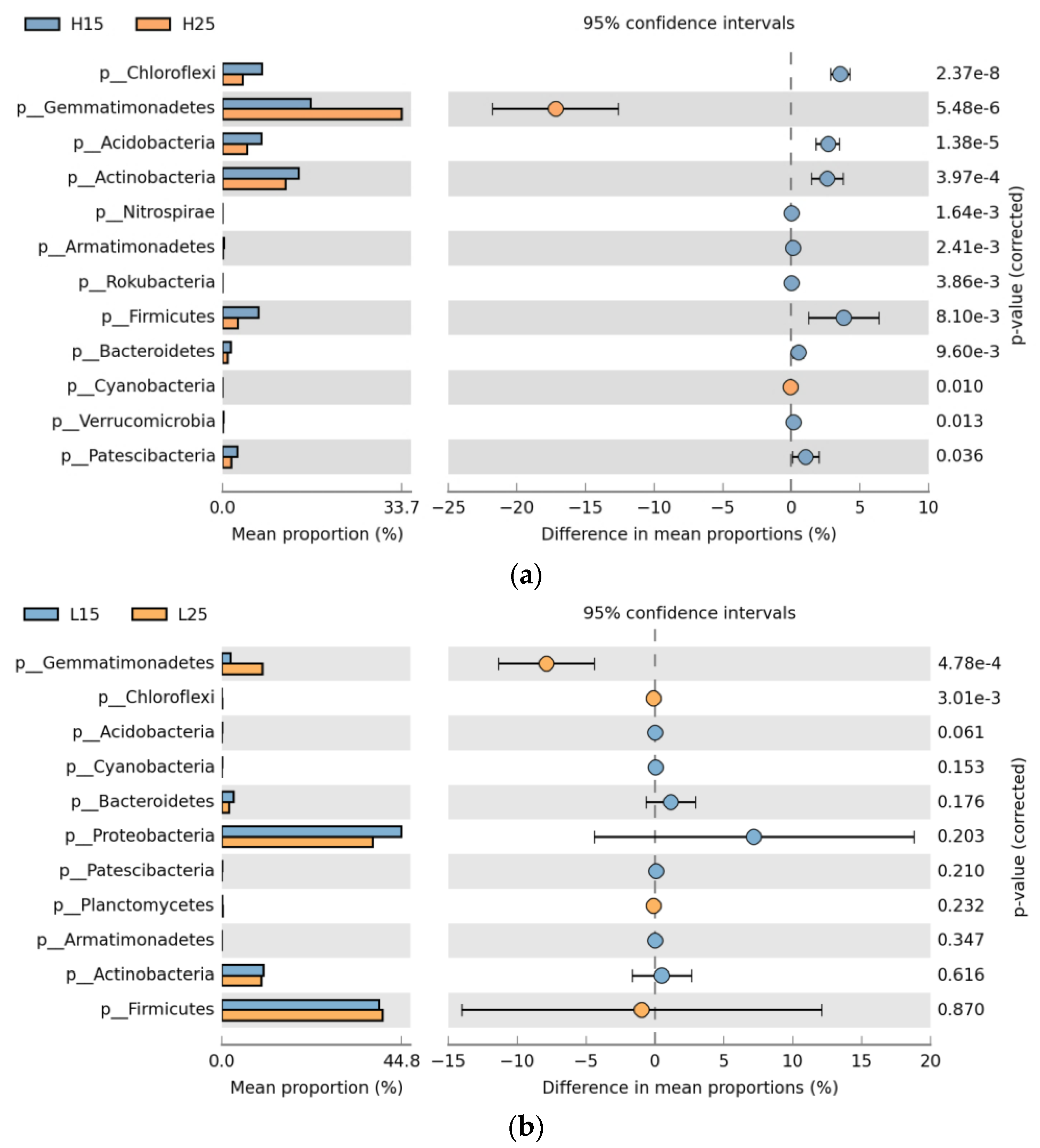
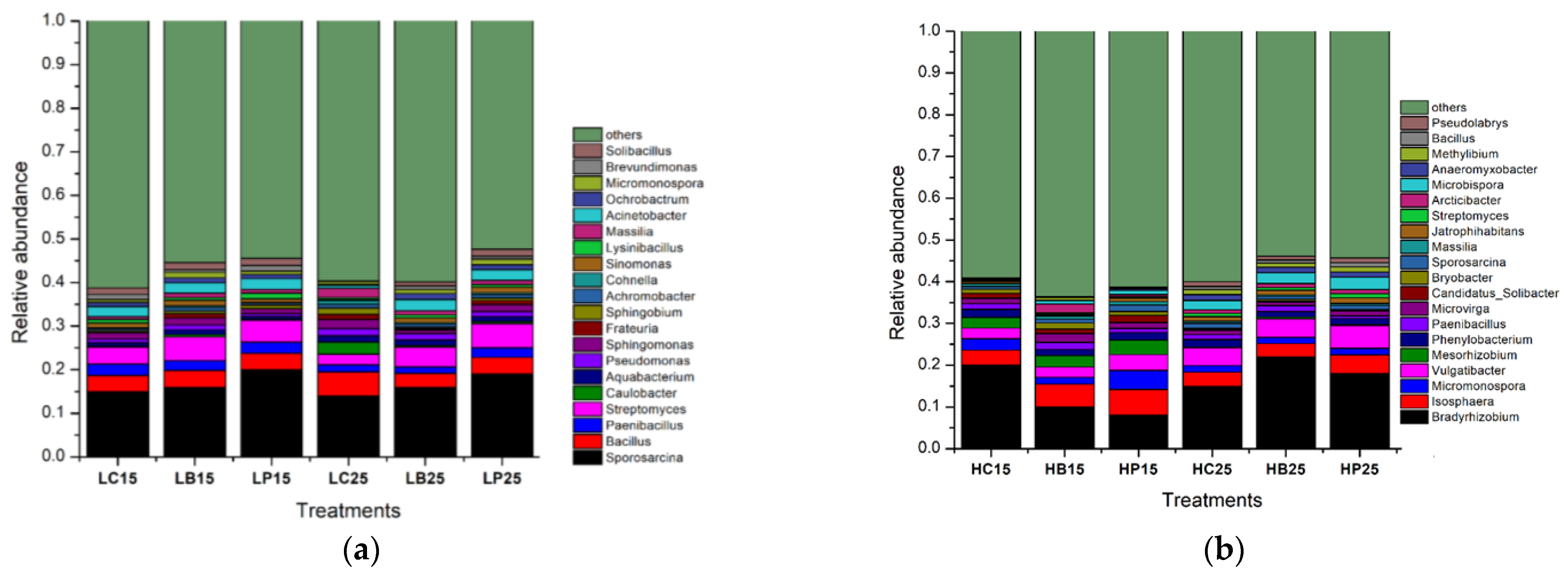
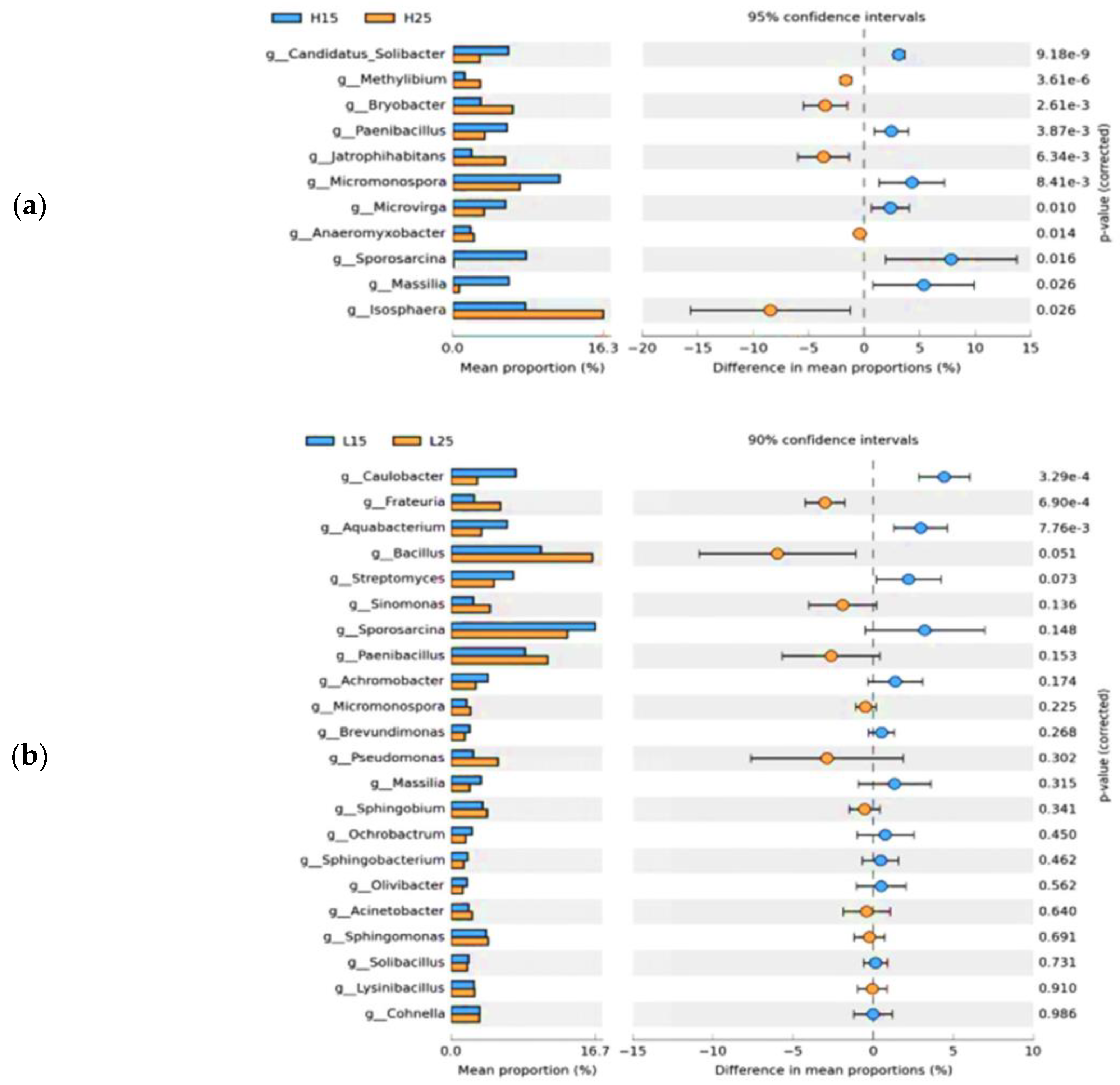
3.2. Dynamics of Soil Bacterial Community after Biochar Fertilizer Amendment
3.3. Relationship between Soil Bacterial Communities and Chemical Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, A.; Ahmad, M.; Mumtaz, M.Z.; Ali, S.; Sarfraz, R.; Naveed, M.; Jamil, M.; Damalas, C.A. Integrated Application of Organic Amendments with Alcaligenes sp. AZ9 Improves Nutrient Uptake and Yield of Maize (Zea mays). J. Plant Growth Regul. 2020, 39, 1277–1292. [Google Scholar] [CrossRef]
- Yang, W.; Li, P.; Rensing, C.; Ni, W.; Xing, S. Biomass, activity and structure of rhizosphere soil microbial community under different metallophytes in a mining site. Plant Soil 2019, 434, 245–262. [Google Scholar] [CrossRef]
- Sarfraz, R.; Shakoor, A.; Abdullah, M.; Arooj, A.; Hussain, A.; Xing, S. Impact of integrated application of biochar and nitrogen fertilizers on maize growth and nitrogen recovery in alkaline calcareous soil. Soil Sci. Plant Nutr. 2017, 63, 488–498. [Google Scholar] [CrossRef]
- Tarin, M.W.K. Effects of different biochars ammendments on physiochemical properties of soil and root morphological attributes of Fokenia Hodginsii (Fujian cypress). Appl. Ecol. Environ. Res. 2019, 17, 11107–11120. [Google Scholar] [CrossRef]
- Zheng, B.-X.; Ding, K.; Yang, X.-R.; Wadaan, M.A.; Hozzein, W.N.; Peñuelas, J.; Zhu, Y.-G. Straw biochar increases the abundance of inorganic phosphate solubilizing bacterial community for better rape (Brassica napus) growth and phosphate uptake. Sci. Total. Environ. 2019, 647, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, L.; Xu, J.; Xin, F.; Jiang, L. Applications of synthetic microbial consortia in biological control of mycotoxins and fungi. Curr. Opin. Food Sci. 2023, 53, 101074. [Google Scholar] [CrossRef]
- Tarin, M.W.K.; Fan, L.; Tayyab, M.; Sarfraz, R.; He, T.; Rong, J.; Chen, L.; Zheng, Y. Effects of bamboo biochar amendment on the growth and physiological characteristics of fokienia hodginsii. Appl. Ecol. Environ. Res. 2018, 16, 8055–8074. [Google Scholar] [CrossRef]
- Tayyab, M. Biochar: An efficient way to manage low water availability in plants. Appl. Ecol. Environ. Res. 2018, 16, 2565–2583. [Google Scholar] [CrossRef]
- Kumar, U.; Shahid, M.; Tripathi, R.; Mohanty, S.; Kumar, A.; Bhattacharyya, P.; Lal, B.; Gautam, P.; Raja, R.; Panda, B.B.; et al. Variation of functional diversity of soil microbial community in sub-humid tropical rice-rice cropping system under long-term organic and inorganic fertilization. Ecol. Indic. 2017, 73, 536–543. [Google Scholar] [CrossRef]
- Estrada-Bonilla, G.A.; Lopes, C.M.; Durrer, A.; Alves, P.R.; Passaglia, N.; Cardoso, E.J. Effect of phosphate-solubilizing bacteria on phosphorus dynamics and the bacterial community during composting of sugarcane industry waste Effect of phosphate-solubilizing bacteria on phosphorus dynamics and the bacterial community during composting of su. Syst. Appl. Microbiol. 2017, 40, 308–313. [Google Scholar] [CrossRef]
- Corneo, P.E.; Pellegrini, A.; Cappellin, L.; Gessler, C.; Pertot, I. Moderate warming in microcosm experiment does not affect microbial communities in temperate vineyard soils. Microb. Ecol. 2014, 67, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-C.; Fang, X.-M.; Wang, G.G.; Mao, R.; Lin, X.-F.; Wang, H.; Chen, F.-S. Effects of nutrient addition on foliar phosphorus fractions and their resorption in different-aged leaves of Chinese fir in subtropical China. Plant Soil 2019, 443, 41–54. [Google Scholar] [CrossRef]
- Li, Y.; Hu, S.; Chen, J.; Mueller, K.; Li, Y.; Fu, W.; Lin, Z.; Wang, H. Effects of biochar application in forest ecosystems on soil properties and greenhouse gas emissions: A review. J. Soils Sediments 2018, 18, 546–563. [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Arafat, Y.; Pang, Z.; Zhang, C.; Lin, Y.; Waqas, M.; Lin, S.; Lin, W.; Zhang, H. Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities. Sustainability 2018, 10, 2361. [Google Scholar] [CrossRef]
- McLean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties; American Society of Agronomy, Inc.: Madison, WI, USA, 1982. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Ng, C.W.W.; Wang, Y.C.; Ni, J.J.; So, P.S. Effects of phosphorus-modified biochar as a soil amendment on the growth and quality of Pseudostellaria heterophylla. Sci. Rep. 2022, 12, 7268. [Google Scholar] [CrossRef]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Fekih, I.B.; Ditta, A.; Xing, S. Role of biochar and plant growth promoting rhizobacteria to enhance soil carbon sequestration—a review. Environ. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef]
- Lu, W.; Ding, W.; Zhang, J.; Li, Y.; Luo, J.; Bolan, N.; Xie, Z. Biochar suppressed the decomposition of organic carbon in a cultivated sandy loam soil: A negative priming effect. Soil Biol. Biochem. 2014, 76, 12–21. [Google Scholar] [CrossRef]
- Onwuka, B.M. Effects of Soil Temperature on Some Soil Properties and Plant Growth. Adv. Plants Agric. Res. 2018, 8, 34–37. [Google Scholar] [CrossRef]
- Sarfraz, R.; Yang, W.; Wang, S.; Zhou, B.; Xing, S. Short term effects of biochar with different particle sizes on phosphorous availability and microbial communities. Chemosphere 2020, 256, 126862. [Google Scholar] [CrossRef]
- Xu, G.; Sun, J.; Shao, H.; Chang, S.X. Biochar had effects on phosphorus sorption and desorption in three soils with differing acidity. Ecol. Eng. 2014, 62, 54–60. [Google Scholar] [CrossRef]
- Luo, D.; Wang, L.; Nan, H.; Cao, Y.; Wang, H.; Kumar, T.V.; Wang, C. Phosphorus adsorption by functionalized biochar: A review. Environ. Chem. Lett. 2023, 21, 497–524. [Google Scholar] [CrossRef]
- Gao, S.; DeLuca, T.H. Wood biochar impacts soil phosphorus dynamics and microbial communities in organically-managed croplands. Soil Biol. Biochem. 2018, 126, 144–150. [Google Scholar] [CrossRef]
- Ahmed, W.; Jing, H.; Kaillou, L.; Qaswar, M.; Khan, M.N.; Jin, C.; Geng, S.; Qinghai, H.; Yiren, L.; Guangrong, L.; et al. Changes in phosphorus fractions associated with soil chemical properties under long-term organic and inorganic fertilization in paddy soils of southern China. PLoS ONE 2019, 14, e0216881. [Google Scholar] [CrossRef] [PubMed]
- Kahura, M.W.; Min, H.; Kim, M.S.; Kim, J.G. Assessing Phosphorus Availability in a High pH, Biochar Amended Soil under Inorganic and Organic Fertilization. Ecol. Resilient Infrastruct. 2018, 5, 11–18. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, Z.; Zhu, Y. Changes in abiotic dissipation rates and bound fractions of antibiotics in biochar-amended soil. J. Clean. Prod. 2020, 256, 120314. [Google Scholar] [CrossRef]
- Fan, J.; Wang, J.-Y.; Hu, X.-F.; Chen, F.-S. Seasonal dynamics of soil nitrogen availability and phosphorus fractions under urban forest remnants of different vegetation communities in Southern China. Urban For. Urban Green. 2014, 13, 576–585. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 54, 655–670. [Google Scholar] [CrossRef]
- Saha, S.; Prakash, V.; Kundu, S.; Kumar, N.; Lal Mina, B. Soil enzymatic activity as affected by long term application of farm yard manure and mineral fertilizer under a rainfed soybean-wheat system in N-W Himalaya. Eur. J. Soil Biol. 2008, 44, 309–315. [Google Scholar] [CrossRef]
- Kader, M.; Yeasmin, S.; Solaiman, Z.; De Neve, S.; Sleutel, S. Response of hydrolytic enzyme activities and nitrogen mineralization to fertilizer and organic matter application in subtropical paddy soils. Eur. J. Soil Biol. 2017, 80, 27–34. [Google Scholar] [CrossRef]
- Zhang, T.; Li, H.; Yan, T.; Shaheen, S.M.; Niu, Y.; Xie, S.; Zhang, Y.; Abdelrahman, H.; Ali, E.F.; Bolan, N.S.; et al. Organic matter stabilization and phosphorus activation during vegetable waste composting: Multivariate and multiscale investigation. Sci. Total. Environ. 2023, 891, 164608. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yu, X.; Song, C.; Pang, X.; Huang, J.; Li, Y. Effect of pyrolysis temperature on the chemical oxidation stability of bamboo biochar. Bioresour. Technol. 2016, 218, 1303–1306. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, W.; Hao, X.; Zhou, D. Transport of Biochar Particles in Saturated Granular Media: E ff ects of Pyrolysis Temperature and Particle Size. Environ. Sci. Technol. 2013, 47, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Dugan, B.; Masiello, C.A.; Gonnermann, H.M. Biochar particle size, shape, and porosity act together to influence soil water properties. PLoS ONE 2017, 12, e0179079. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Islam, F.; Ali, B.; Pei, Z.; Li, J.; Ghani, M.A.; Zhou, W. Silicon and water-deficit stress differentially modulate physiology and ultrastructure in wheat (Triticum aestivum L.). 3 Biotech 2017, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.-L.; Mortimer, P.E.; Ferry Slik, J.W.; Zou, X.-M.; Xu, J.; Feng, W.-T.; Qiao, L. Variation in forest soil fungal diversity along a latitudinal gradient. Fungal Divers. 2014, 64, 305–315 . [Google Scholar] [CrossRef]
- Liu, M.; Liu, J.; Chen, X.; Jiang, C.; Wu, M.; Li, Z. Shifts in bacterial and fungal diversity in a paddy soil faced with phosphorus surplus. Biol. Fertil. Soils 2018, 54, 259–267. [Google Scholar] [CrossRef]
- Deng, J.; Yin, Y.; Zhu, W.; Zhou, Y. Variations in soil bacterial community diversity and structures among different revegetation types in the Baishilazi nature reserve. Front. Microbiol. 2018, 9, 2874. [Google Scholar] [CrossRef]
- Han, G.; Lan, J.; Chen, Q.; Yu, C.; Bie, S. Response of soil microbial community to application of biochar in cotton soils with different continuous cropping years. Sci. Rep. 2017, 7, 10184. [Google Scholar] [CrossRef]
- Yu, H.; Yan, X.; Zheng, X.; Xu, K.; Zhong, Q.; Yang, T.; Liu, F.; Wang, C.; Shu, L.; He, Z.; et al. Differential distribution of and similar biochemical responses to different species of arsenic and antimony in Vetiveria zizanioides. Environ. Geochem. Heal. 2020, 42, 3995–4010. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Anwar, Y.; Hasan, M.M.; Iqbal, A.; Ali, M.; Alharby, H.F.; Hakeem, K.R.; Hasanuzzaman, M. Attenuation of Drought Stress in Brassica Seedlings with Exogenous Application of Ca2+ and H2O2. Plants 2017, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, Y.; Dijkstra, F.A.; Li, Z.; Zhang, Y.; Zhang, T.; Lu, Y.; Shi, J.; Yang, L. Effects of amendments on phosphorous status in soils with different phosphorous levels. Catena 2019, 172, 97–103. [Google Scholar] [CrossRef]
- Wang, H.H.; Chu, H.L.; Dou, Q.; Xie, Q.Z.; Tang, M.; Sung, C.K.; Wang, C.Y. Phosphorus and nitrogen drive the seasonal dynamics of bacterial communities in Pinus forest rhizospheric soil of the Qinling Mountains. Front. Microbiol. 2018, 9, 1930. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Paddy Soil (H) | Red Soil (L) |
|---|---|---|
| pH | 5.29 | 5.12 |
| Total P (g kg−1) | 2.54 | 0.56 |
| available P (mg kg−1) | 11.2 | 3.48 |
| Factors | pH | Available P | Acid Phosphatase Activity | ||||
|---|---|---|---|---|---|---|---|
| Soil Types | F Value | p-Value | F Value | p-Value | F Value | p-Value | |
| Treatments (Trt.) | Red soil | 280.93 | <0.001 * | 10,262.66 | 0.002 * | 3509.14 | 0.003 * |
| Paddy soil | 524.70 | <0.001 * | 10,234.78 | <0.001 * | 5094.32 | <0.001 * | |
| DOI | Red soil | 4.51 | <0.001 * | 699.56 | <0.001 * | 1.94 | 0.10 |
| Paddy soil | 8.62 | <0.001 * | 694.60 | <0.001 * | 1.57 | 0.18 | |
| T | Red soil | 0.03 | 0.85 | 0.67 | 0.41 | 0.05 | 0.83 |
| Paddy soil | 3.03 | 0.09 | 1.98 | 0.16 | 0.09 | 0.77 | |
| DOI × Trt. | Red soil | 1.02 | 0.44 | 178.53 | <0.001 * | 1.91 | 0.06 |
| Paddy soil | 1.97 | 0.05 | 177.48 | <0.001 * | 2.91 | <0.001 * | |
| DOI × T | Red soil | 3.98 | <0.001 * | 8.42 | <0.001 * | 0.16 | 0.97 |
| Paddy soil | 3.75 | <0.001 * | 10.60 | <0.001 * | 0.40 | 0.85 | |
| T × Trt. | Red soil | 0.33 | 0.72 | 1.07 | 0.35 | 0.23 | 0.80 |
| Paddy soil | 0.70 | 0.50 | 0.72 | 0.49 | 0.99 | 0.38 | |
| DOI × T × Trt. | Red soil | 1.24 | 0.28 | 4.82 | <0.001 * | 2.62 | 0.01 * |
| Paddy soil | 1.45 | 0.18 | 6.47 | <0.001 * | 4.21 | <0.001 * | |
| Properties | Soil Types | Trt | T | Soil * Trt. | Trt. * T | Soil * T | Soil * Trt. * T | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | sig. | F | sig. | F | sig. | F | sig. | F | sig. | F | sig. | F | sig. | |
| pH | 0.00 | 0.97 | 0.00 | 1.00 | 0.02 | 0.90 | 0.01 | 1.00 | 0.00 | 0.98 | 0.00 | 1.00 | 0.00 | 1.00 |
| Available P | 0.23 | 0.64 | 0.00 | 1.00 | 0.04 | 0.84 | 0.00 | 1.00 | 0.00 | 1.00 | 0.00 | 1.00 | 0.00 | 1.00 |
| Acid phosphatase | 0.00 | 0.95 | 0.03 | 0.97 | 0.00 | 0.98 | 0.00 | 1.00 | 0.00 | 0.98 | 0.00 | 1.00 | 0.00 | 1.00 |
| Proteobacteria | 2.84 | 0.11 | 0.12 | 0.89 | 2.47 | 0.13 | 0.04 | 0.97 | 0.35 | 0.56 | 0.11 | 0.90 | 0.09 | 0.91 |
| Gemmatimonadetes | 167.87 | <0.001 * | 0.09 | 0.91 | 72.85 | <0.001 * | 0.12 | 0.89 | 10.07 | <0.001 * | 0.15 | 0.86 | 0.26 | 0.78 |
| Actinobacteria | 26.39 | <0.001 * | 0.47 | 0.63 | 7.08 | 0.01 * | 0.08 | 0.92 | 3.27 | 0.08 | 0.99 | 0.39 | 0.94 | 0.40 |
| Chloroflexi | 946.53 | <0.001 * | 0.38 | 0.69 | 91.97 | <0.001 * | 0.42 | 0.66 | 102.09 | <0.001 * | 0.18 | 0.84 | 0.14 | 0.87 |
| Acidobacteria | 781.69 | <0.001 * | 0.90 | 0.42 | 40.03 | <0.001 | 0.76 | 0.48 | 38.44 | <0.001 * | 0.03 | 0.97 | 0.03 | 0.97 |
| Planctomycetes | 58.65 | <0.001 * | 0.90 | 0.42 | 0.72 | 0.41 | 0.80 | 0.46 | 0.49 | 0.49 | 0.51 | 0.61 | 0.37 | 0.69 |
| Firmicutes | 110.53 | <0.001 * | 0.05 | 0.95 | 0.18 | 0.67 | 0.08 | 0.93 | 0.52 | 0.48 | 0.09 | 0.92 | 0.10 | 0.91 |
| Patescibacteria | 82.47 | <0.001 * | 1.23 | 0.31 | 5.71 | 0.03 * | 0.67 | 0.52 | 4.07 | 0.06 | 0.28 | 0.76 | 0.14 | 0.87 |
| Bacteroidetes | 6.68 | 0.02 * | 0.11 | 0.90 | 3.37 | 0.08 | 0.03 | 0.97 | 0.48 | 0.50 | 0.03 | 0.97 | 0.15 | 0.87 |
| Paddy Soil | Red Soil | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phylum | Genus | pH | Available P | Acid Phosphatase | Phylum | Genus | pH | Available P | Acid Phosphatase |
| Proteobacteria | Bradyrhizobium | −0.43 | −0.51 * | 0.50 * | Firmicutes | Sporosarcina | −0.11 | −0.27 | 0.58 * |
| Planctomycetes | Isosphaera | −0.14 | −0.16 | 0.11 | Firmicutes | Bacillus | −0.58 * | −0.58 * | 0.59 ** |
| Actinobacteria | Micromonospora | −0.12 | −0.05 | 0.07 | Firmicutes | Paenibacillus | −0.38 | −0.47 * | 0.22 |
| Proteobacteria | Vulgatibacter | 0.28 | 0.33 | −0.32 | Actinobacteria | Streptomyces | 0.13 | −0.19 | 0.25 |
| Proteobacteria | Mesorhizobium | −0.53 * | −0.57 * | 0.56 * | Proteobacteria | Caulobacter | 0.50 * | 0.38 | −0.07 |
| Proteobacteria | Phenylobacterium | 0.33 | 0.25 | −0.22 | Proteobacteria | Aquabacterium | 0.49 * | 0.44 | −0.08 |
| Firmicutes | Paenibacillus | 0.16 | 0.26 | −0.23 | Proteobacteria | Pseudomonas | 0.08 | 0.11 | 0.34 |
| Proteobacteria | Microvirga | −0.24 | −0.27 | 0.26 | Proteobacteria | Sphingomonas | −0.16 | −0.21 | 0.20 |
| Acidobacteria | Candidatus_Solibacter | −0.09 | −0.04 | 0.07 | Proteobacteria | Frateuria | 0.03 | −0.06 | −0.21 |
| Acidobacteria | Bryobacter | −0.29 | −0.29 | 0.33 | Proteobacteria | Sphingobium | −0.58 * | −0.45 | 0.73 ** |
| Firmicutes | Sporosarcina | 0.31 | 0.44 | −0.40 | Proteobacteria | Achromobacter | 0.59 ** | 0.724 ** | −0.59 ** |
| Proteobacteria | Massilia | 0.24 | 0.22 | −0.21 | Firmicutes | Cohnella | −0.22 | −0.42 | 0.34 |
| Actinobacteria | Jatrophihabitans | 0.25 | 0.19 | −0.26 | Actinobacteria | Sinomonas | −0.03 | 0.22 | −0.69 ** |
| Actinobacteria | Streptomyces | −0.49 * | −0.44 | 0.43 | Firmicutes | Lysinibacillus | 0.15 | −0.11 | 0.44 |
| Bacteroidetes | Arcticibacter | −0.31 | −0.40 | 0.43 | Proteobacteria | Massilia | 0.62 ** | 0.17 | −0.65 ** |
| Actinobacteria | Microbispora | −0.39 | −0.3 | 0.35 | Proteobacteria | Acinetobacter | −0.36 | −0.34 | 0.57 * |
| Actinobacteria | Anaeromyxobacter | −0.09 | 0.00 | 0.04 | Proteobacteria | Ochrobactrum | 0.03 | 0.00 | 0.14 |
| Proteobacteria | Methylibium | −0.19 | −0.20 | 0.16 | Actinobacteria | Micromonospora | −0.23 | −0.09 | 0.06 |
| Proteobacteria | Bacillus | 0.34 | 0.26 | −0.25 | Proteobacteria | Brevundimonas | 0.52 * | −0.04 | 0.16 |
| Proteobacteria | Pseudolabrys | −0.44 | −0.44 | 0.44 | Firmicutes | Solibacillus | 0.13 | 0.08 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarfraz, R.; Nadeem, F.; Yang, W.; Tayyab, M.; Khan, M.I.; Mahmood, R.; Guo, X.; Xing, S.; Kim, G.W. Evaluation of Biochar and Inorganic Fertilizer on Soil Available Phosphorus and Bacterial Community Dynamics in Acidic Paddy Soils for Different Incubation Temperatures. Agronomy 2024, 14, 26. https://doi.org/10.3390/agronomy14010026
Sarfraz R, Nadeem F, Yang W, Tayyab M, Khan MI, Mahmood R, Guo X, Xing S, Kim GW. Evaluation of Biochar and Inorganic Fertilizer on Soil Available Phosphorus and Bacterial Community Dynamics in Acidic Paddy Soils for Different Incubation Temperatures. Agronomy. 2024; 14(1):26. https://doi.org/10.3390/agronomy14010026
Chicago/Turabian StyleSarfraz, Rubab, Faisal Nadeem, Wenhao Yang, Muhammad Tayyab, Muhammad Israr Khan, Rashid Mahmood, Xingjie Guo, Shihe Xing, and Gil Won Kim. 2024. "Evaluation of Biochar and Inorganic Fertilizer on Soil Available Phosphorus and Bacterial Community Dynamics in Acidic Paddy Soils for Different Incubation Temperatures" Agronomy 14, no. 1: 26. https://doi.org/10.3390/agronomy14010026