

Turnip Yellows Virus Field Infection in Oilseed Rape: Does It Impact the Yield and Quality?
Abstract
:1. Introduction
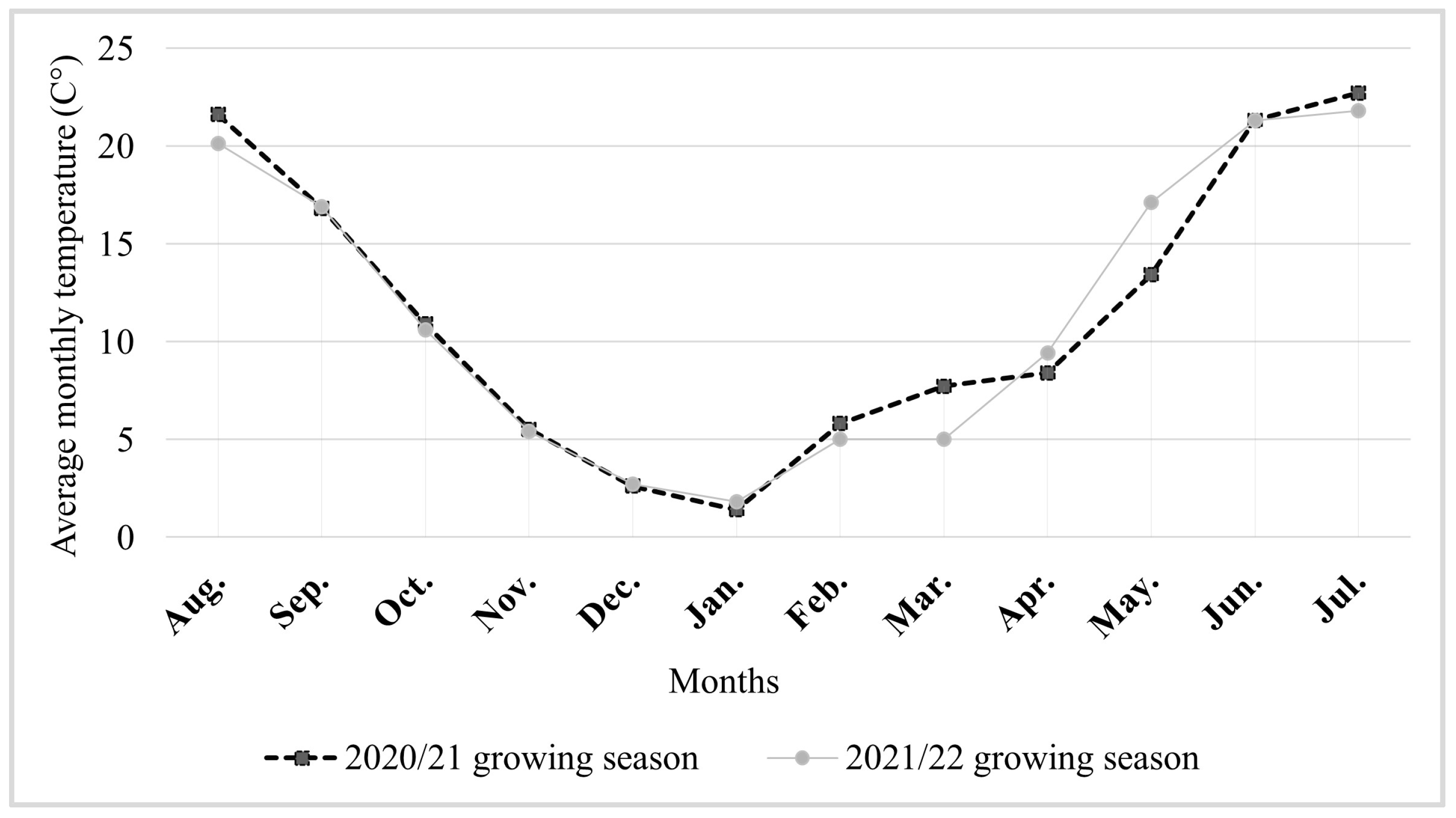
2. Materials and Methods
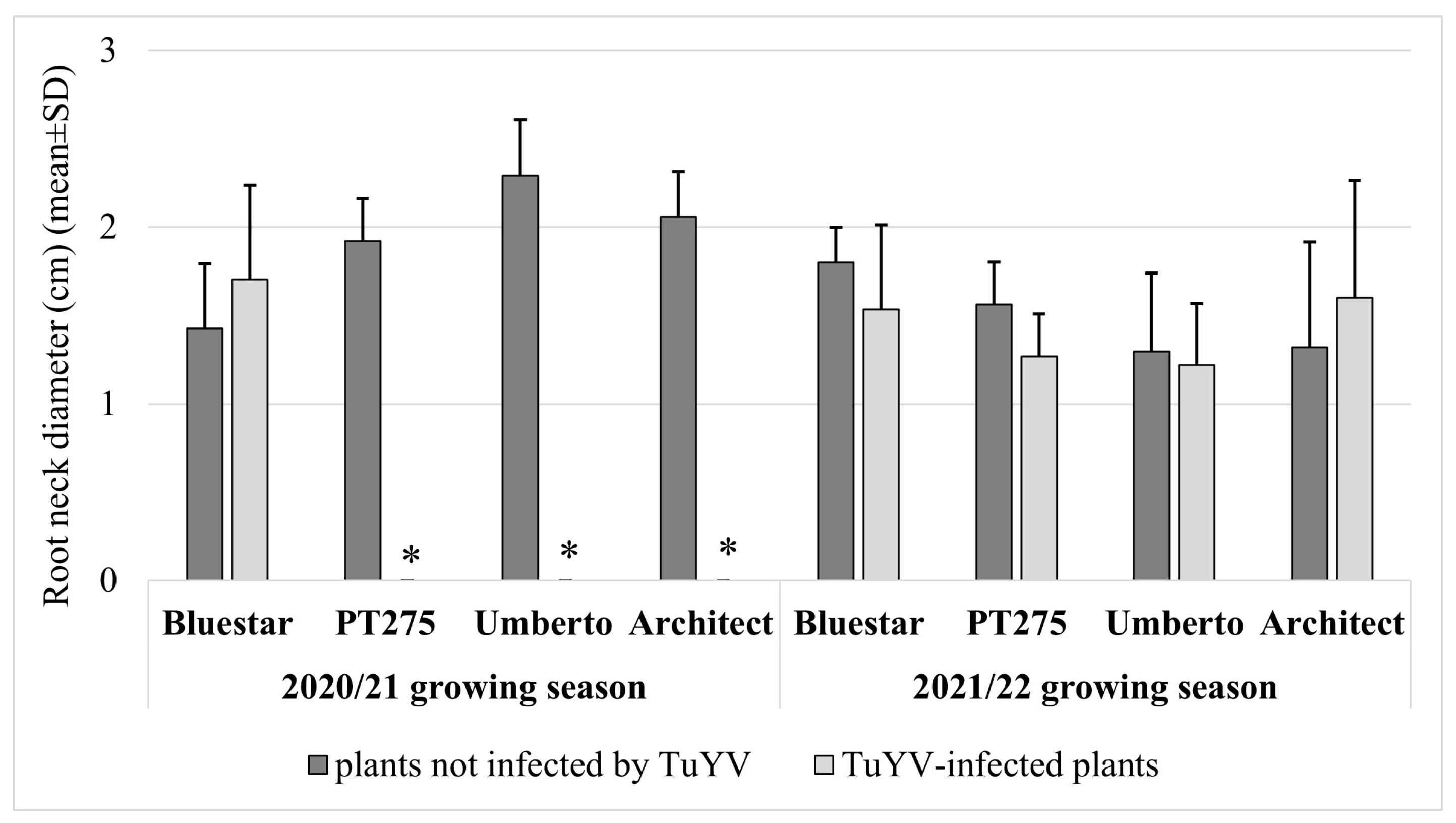
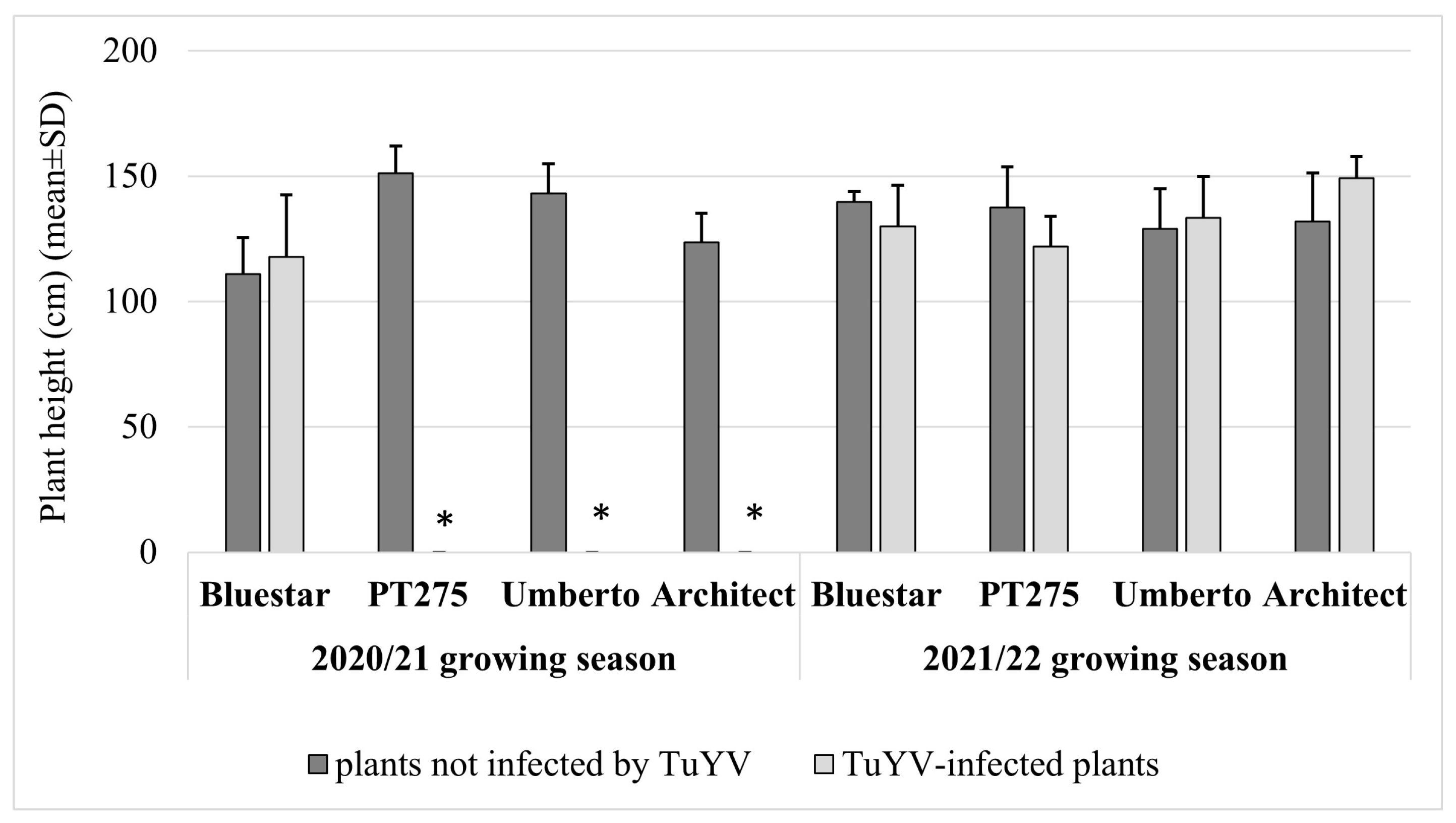
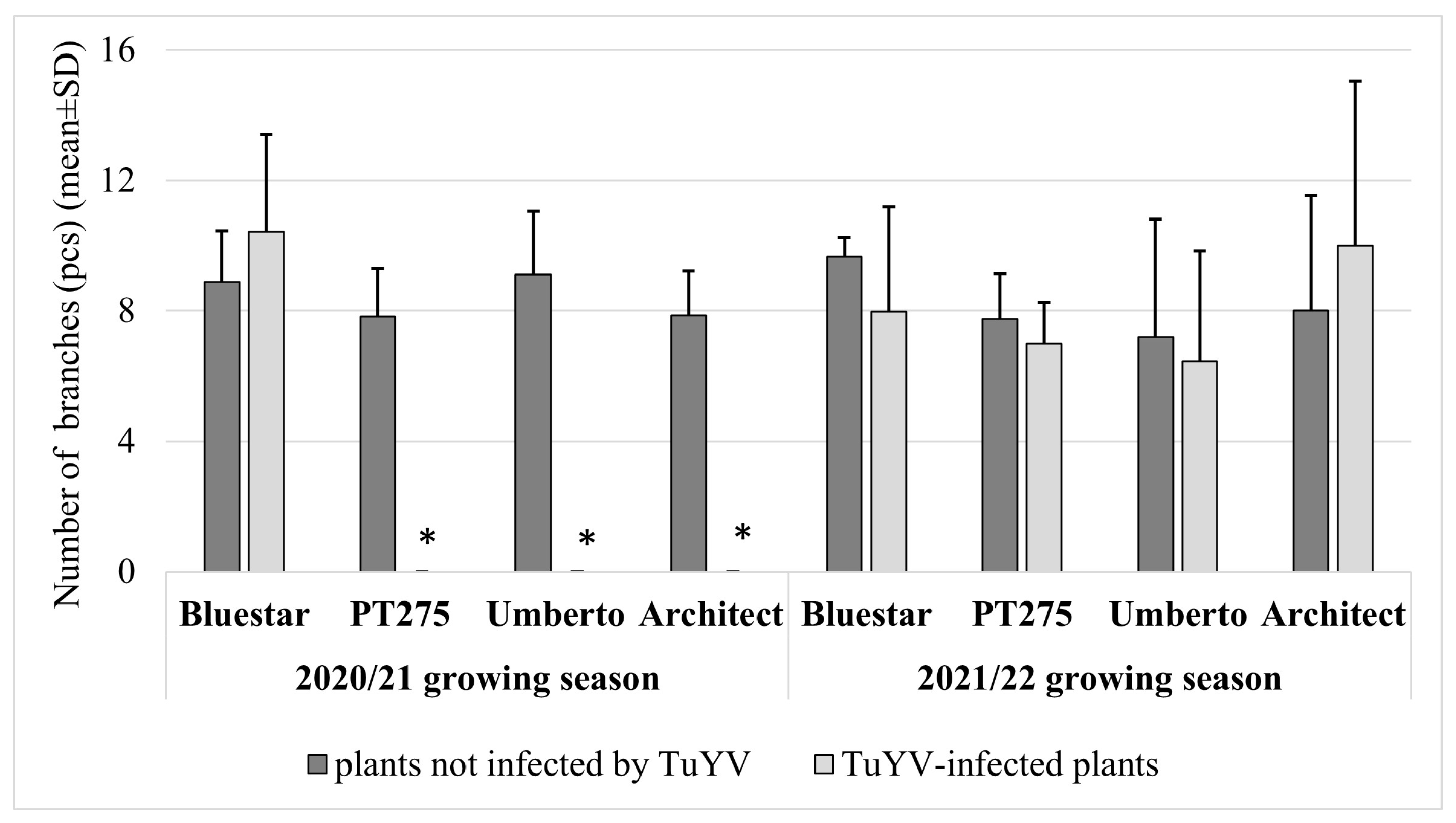
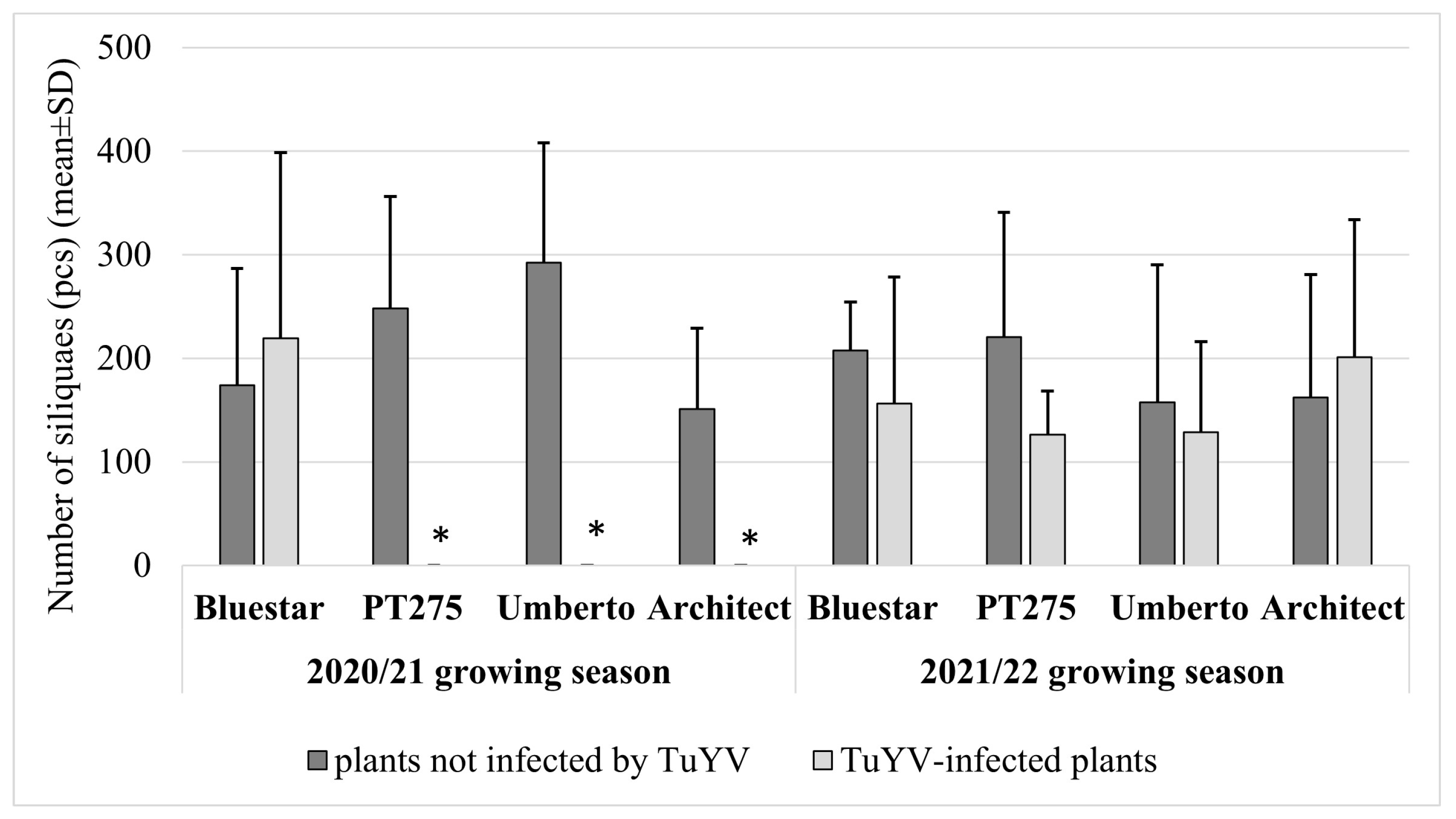
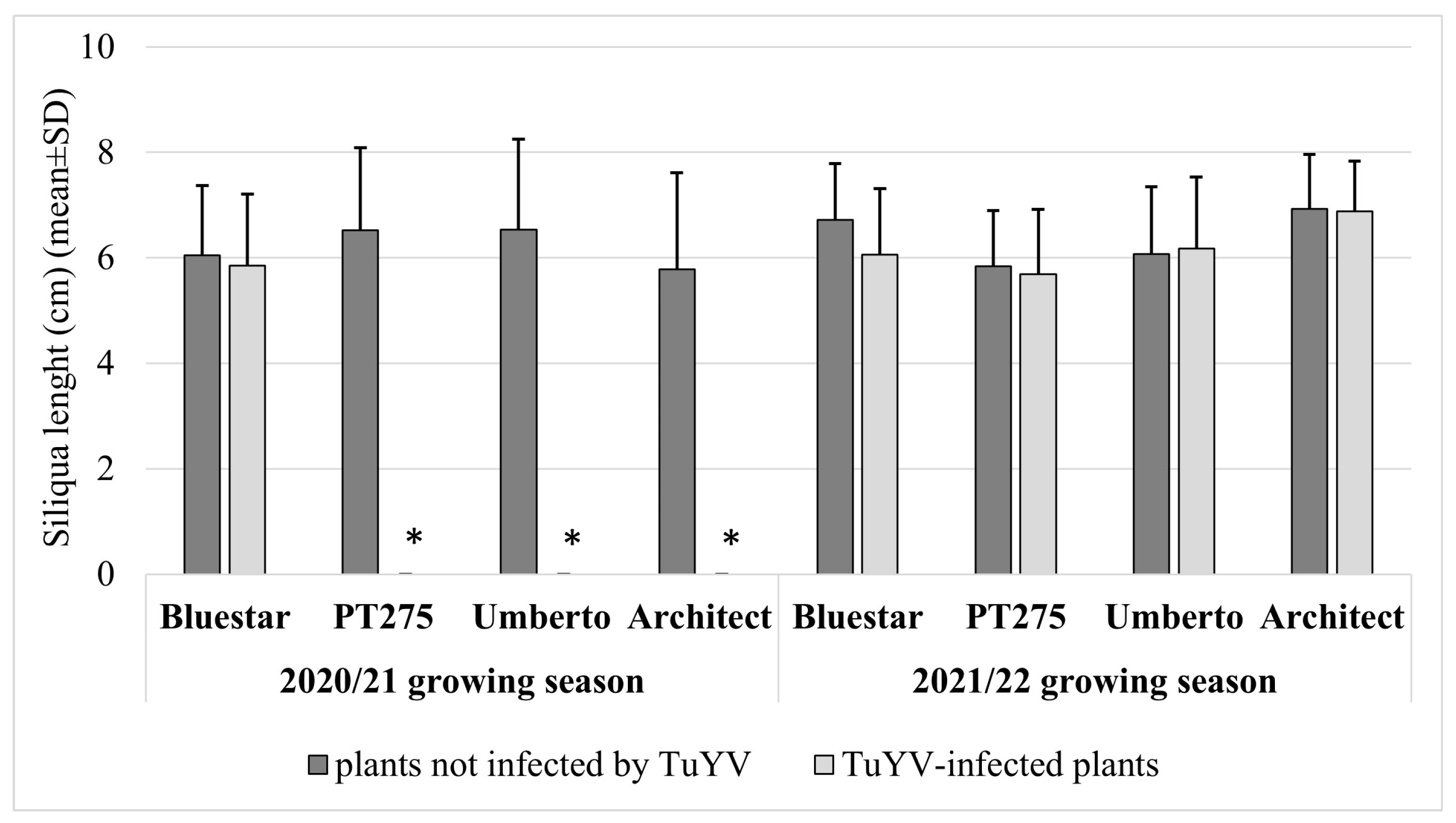
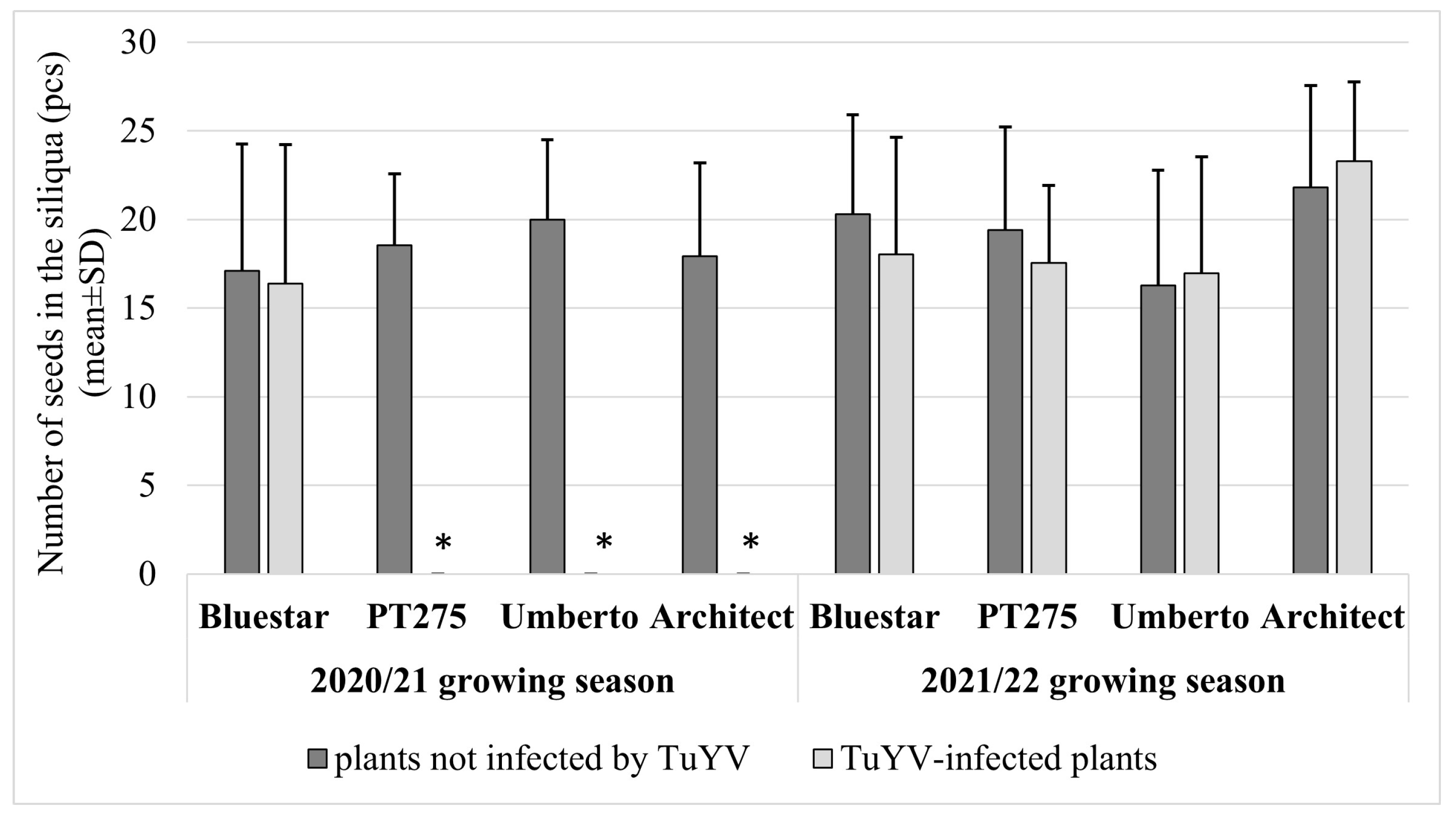
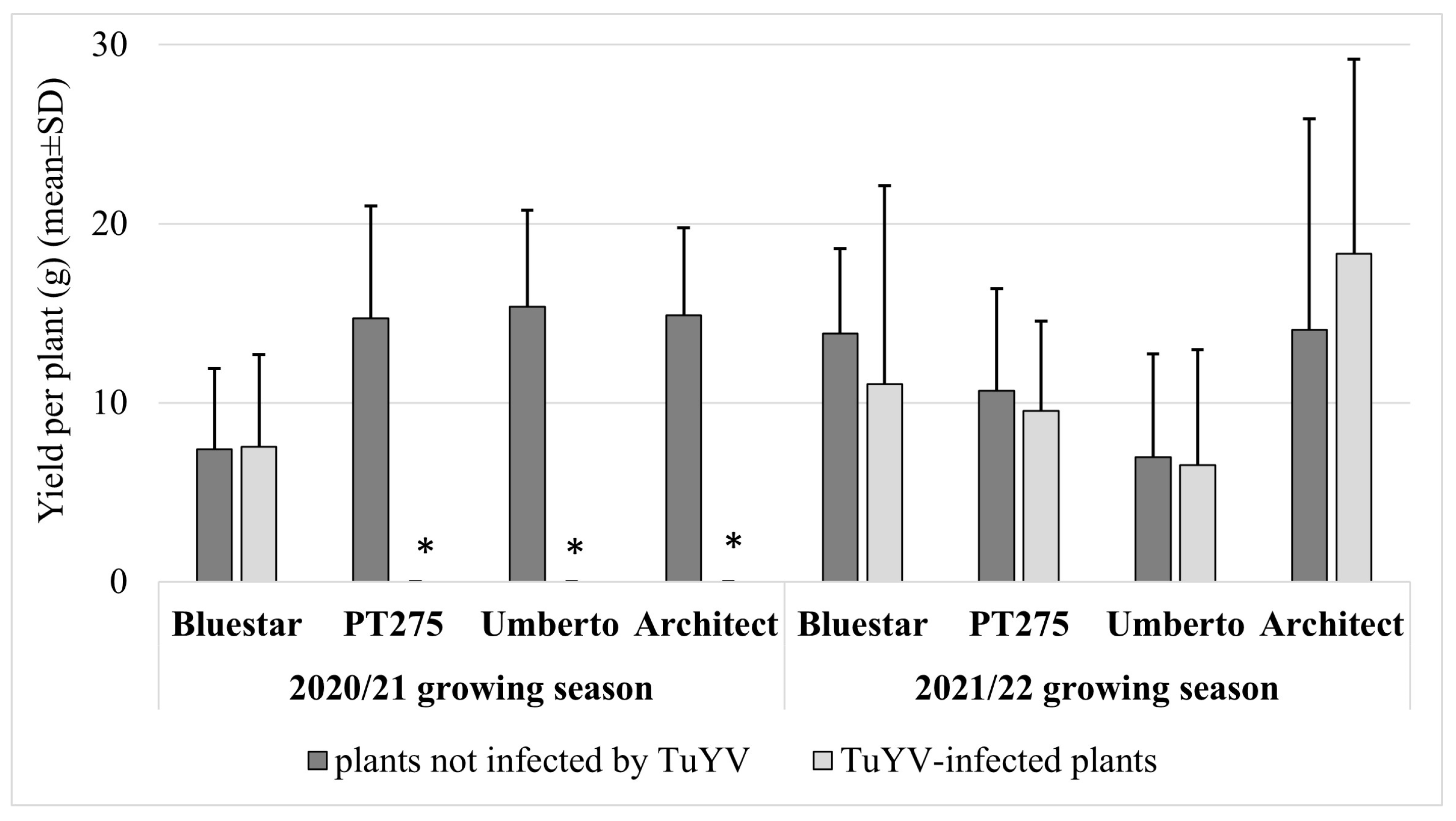
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eurostat. 2021. Available online: https://agridata.ec.europa.eu/extensions/DashboardCereals/OilseedProduction.html# (accessed on 2 June 2023).
- Choi, B.; Lim, J.E.; Sung, J.K.; Jeon, W.T.; Lee, S.S.; Oh, S.E.; Yang, J.E.; Ok, Y.S. Effect of rapeseed green manure amendment on soil properties and rice productivity. Commun. Soil Sci. Plant Anal. 2014, 45, 751–764. [Google Scholar] [CrossRef]
- Fitt, B.D.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Sustainable strategies for managing Brassica napus (oilseed rape) resistance to Leptosphaeria maculans (phoma stem canker). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Zală, C.R.; Cristea, S.; Radu, E.; Manole, M.S.; Bălaşu, V.A.; Clinciu, E.C. Research regarding the response of rapeseed hybrids to pathogens. Sci. Papers. Ser. A. Agron. 2012, 55, 265–268. [Google Scholar]
- Nooh, S. An overview of oilseed rape (canola) virus diseases in Iran. Int. Res. J. Microbiol. 2012, 3, 24–28. [Google Scholar]
- Graichen, K.; Rabenstein, F.; Kurtz, E. The occurrence of Turnip yellows virus in winter oilseed rape in Austria. Pflanzenschutzberichte 2000, 59, 35–46. [Google Scholar]
- Milošević, D.; Marjanović-Jeromela, A.; Ignjatov, M.; Jovičić, D.; Stanković, I.; Bulajić, A.; Krstić, B. First report of Turnip yellows virus on oilseed rape in Serbia. Plant Dis. 2015, 99, 1869. [Google Scholar] [CrossRef]
- Newbert, M.J. The Genetic Diversity of Turnip Yellows Virus in Oilseed Rape (Brassica napus) in Europe, Pathogenic Determinants, New Sources of Resistance and Host Range. Doctoral Dissertation, University of Warwick, Coventry, UK, 2016. [Google Scholar]
- Konradyová, V.; Zusková, E.; Grimová, L.; Ryšánek, P.; Kazda, J. Virus diseases of oilseed rape (Brassica napus subs. napus) in the Czech Republic. In Proceedings of the OBC-WPRS Working Group “Integrated Control in Oilseed Crops”, Zagreb, Croatia, 18–20 September 2018. [Google Scholar]
- Orfanidou, C.G.; Lotos, L.; Tsiolakis, G.; Stefanidis, S.K.; Tsialtas, J.T.; Katis, N.I.; Maliogka, V.I. Molecular characterization of poleroviruses isolated from oilseed rape in Greece. Virus Genes 2021, 57, 289–292. [Google Scholar] [CrossRef]
- Sharma, P.; Verma, R.K.; Mishra, R.; Choudhary, D.K.; Gaur, R.K. First report of Turnip yellow virus (TuYV) in Brassica juncea (Indian mustard) in India. New Dis. Rep. 2013, 27, 21. [Google Scholar] [CrossRef]
- Wang, F.; Wu, Q.F.; Zhou, B.G.; Gao, Z.L.; Xu, D.F. First Report of Turnip yellows virus in Tobacco in China. Plant Dis. 2015, 99, 1870. [Google Scholar] [CrossRef]
- New, S.A.; Van Heerden, S.W.; Pietersen, G.; Esterhuizen, L.L. First report of a Turnip yellows virus in association with the brassica stunting disorder in South Africa. Plant Dis. 2016, 100, 2341. [Google Scholar] [CrossRef]
- Kehoe, M.A.; Coutts, B.A. Turnip yellows virus and Soybean dwarf virus in Western Australia. Australas. Plant Pathol. 2019, 48, 323–329. [Google Scholar] [CrossRef]
- Stevens, M.; McGrann, G.; Clark, B. Turnip Yellows Virus (Syn Beet Western Yellows Virus): An Emerging Threat to European Oilseed Rape Production; HGCA: Cape Town, South Africa, 2008. [Google Scholar]
- Slavíková, L.; Ibrahim, E.; Alquicer, G.; Tomašechová, J.; Šoltys, K.; Glasa, M.; Kundu, J.K. Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop. Viruses 2022, 14, 2511. [Google Scholar] [CrossRef] [PubMed]
- Jay, C.N.; Rossall, S.; Smith, H.G. Effects of beet western yellows virus on growth and yield of oilseed rape (Brassica napus). J. Agric. Sci. 1999, 133, 131–139. [Google Scholar] [CrossRef]
- Schliephake, E.; Graichen, K.; Rabenstein, F. Investigations on the vector transmission of the beet mild yellowing virus (BMYV) and the turnip yellows virus (TuYV). J. Plant Dis. Prot. 2000, 107, 81–87. [Google Scholar]
- Kátai, Z. Interactive evaluation of the main agrotechnical factors in rape production. Acta Agrar. Debreceniensis 2009, 36, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Asare-Bediako, E.; Jones, J.E.; Hambidge, A.J.; Stevens, M.; Mead, A.; Jenner, C.E.; Walsh, J.A. The incidence of turnip yellows virus in oilseed rape crops (Brassica napus L.) in three different regions of England over three consecutive growing seasons and the relationship with the abundance of flying Myzus persicae. Ann. Appl. Biol. 2020, 176, 130–137. [Google Scholar] [CrossRef]
- Graichen, K.; Schliephake, E. Occurrence, symptoms and vectors of turnip yellows virus (syn. beet western yellows virus) on winter oilseed rape. Nachrichtenblatt Des Dtsch. Pflanzenschutzdienstes 1996, 48, 186–191. [Google Scholar]
- Graichen, K.; Rabenstein, F. European isolates of beet western yellows virus (BWYV) from oilseed rape (Brassica napus L. ssp. napus) are non-pathogenic on sugar beet (Beta vulgaris L var. altissima) but represent isolates of turnip yellows virus (TuYV). J. Plant Dis. Prot. 1996, 103, 233–245. [Google Scholar]
- D’Arcy, C.J.; Torrance, L.; Martin, R.R. Discrimination among luteoviruses and their strains by monoclonal antibodies and identification of common epitopes. Phytopathology 1989, 79, 869–873. [Google Scholar] [CrossRef]
- Lemaire, O.; Herrbach, E.; Stevens, M.; Bouchery, Y.; Smith, H.G. Detection of sugar-beet-infecting beet mild yellowing virus isolates with a specific RMA probe. Phytopathology 1995, 85, 1513–1518. [Google Scholar] [CrossRef]
- Jones, T.D.; Buck, K.W.; Plumb, R.T. The detection of beet western yellows virus and beet mild yellowing virus in crop plants using the polymerase chain reaction. J. Virol. Methods 1991, 35, 287–296. [Google Scholar] [CrossRef]
- Graichen, K.; Schliephake, E. Infestation of winter oilseed rape by turnip yellows luteovirus and its effect on yield in Germany. In Proceedings of the 10th International Rapeseed Congress—New horizons for an old crop’, Canberra, Australia, 26–29 September 1999. [Google Scholar]
- Jones, R.A.C.; Coutts, B.A.; Hawkes, J. Yield-limiting potential of Beet western yellows virus in Brassica napus. Aust. J. Agric. Res. 2007, 58, 788–801. [Google Scholar] [CrossRef]
- Coutts, B.A.; Jones, R.A.C.; Umina, P.A.; Davidson, J.; Baker, G.; Aftab, M. Beet western yellows virus (synonym: Turnip yellows virus) and green peach aphid in canola. In Proceedings of the 2015 Grains Research and Development Updates, Adelaide, Australia, 10 February 2015. [Google Scholar]
- Barzman, M.; Barberi, P.; Birch, N.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of Integrated Pest Management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Broadbent, L. Factors affecting the activity of alatae of the aphids Myzus persicae (Sulzer) and Brevicoryne brassicae (L.). Ann. Appl. Biol. 1949, 36, 40–62. [Google Scholar] [CrossRef]
- Patel, S.R.; Awasthi, A.K.; Tomar, R.K.S. Assessment of yield losses in mustard (Brassica juncea L.) due to mustard aphid (Lipaphis erysimi Kalt.) under different thermal environments in Eastern Central India. Appl. Ecol. Environ. Res. 2004, 2, 1–15. [Google Scholar] [CrossRef]
- Sharma, A.R.; Singh, V.P. Integrated weed management in conservation agriculture systems. Indian J. Weed Sci. 2014, 46, 23–30. [Google Scholar]
- Jansson, R.K.; Smilowitz, Z. Influence of nitrogen on population parameters of potato insects: Abundance, population growth, and within-plant distribution of the green peach aphid, Myzus persicae (Homoptera: Aphididae). Environ. Entomol. 1986, 15, 49–55. [Google Scholar] [CrossRef]
- Juergens, M.; Paetsch, C.; Krämer, I.; Zahn, M.; Rabenstein, F.; Schondelmaier, J.; Schliepkhake, E.; Snowdon, R.; Friedt, W.; Ordon, F. Genetic analyses of the host-pathogen system Turnip yellows virus (TuYV)—Rapeseed (Brassica napus L.) and development of molecular markers for TuYV-resistance. Theor. Appl. Genet. 2010, 120, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Greer, S.F.; Hackenberg, D.; Gegas, V.; Mitrousia, G.; Edwards, D.; Batley, J.; Teakle, G.R.; Barker, G.C.; Walsh, J.A. Quantitative Trait Locus Mapping of Resistance to Turnip Yellows Virus in Brassica rapa and Brassica oleracea and Introgression of These Resistances by Resynthesis Into Allotetraploid Plants for Deployment in Brassica napus. Front. Plant Sci. 2021, 12, 781385. [Google Scholar] [CrossRef]
- Adam, N.M.; Hopkinson, D. Crop establishment improvements and foliar pest control in winter oilseed rape using a new clothianidin based seed treatment. In Proceedings of the Dundee Conference, Dundee, UK, 26–27 February 2008. [Google Scholar]
- Éghajlati Adatok, Szombathely. Available online: https://www.metnet.hu/napi-adatok?sub=4&pid=15260&date=2021-05-21 (accessed on 12 June 2023).
- Baráth, C.; Ittzés, A.; Ugrósdy, G. Biometria Módszertani Alapok és a Minitab Programcsomag Alkalmazása; Mezőgazda Kiadó: Budapest, Hungary, 1996. [Google Scholar]
- Congdon, B.S.; Baulch, J.R.; Coutts, B.A. Impact of turnip yellows virus infection on seed yield of an open-pollinated and hybrid canola cultivar when inoculated at different growth stages. Virus Res. 2020, 277, 197847. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Hybrid | Infection Based on Symptoms (%) | Infection Based on ELISA Test (%) | Symptom Appearance of Infected Plants Confirmed by ELISA Test (%) |
|---|---|---|---|---|
| 2020/21 | Bluestar | 52.00% | 32.00% | 71.43% |
| PT275 | 24.00% | 0.00% | 0.00% | |
| Umberto | 56.00% | 0.00% | 0.00% | |
| Architect | 24.00% | 0.00% | 0.00% | |
| 2021/22 | Bluestar | 23.33% | 90.00% | 25.93% |
| PT275 | 0.00% | 70.00% | 0.00% | |
| Umberto | 10.00% | 36.67% | 9.09% | |
| Architect | 0.00% | 50.00% | 0.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizi, R.; Kiss, J.; Turóczi, G.; Dobra, N.; Pálinkás, Z. Turnip Yellows Virus Field Infection in Oilseed Rape: Does It Impact the Yield and Quality? Agronomy 2023, 13, 2404. https://doi.org/10.3390/agronomy13092404
Vizi R, Kiss J, Turóczi G, Dobra N, Pálinkás Z. Turnip Yellows Virus Field Infection in Oilseed Rape: Does It Impact the Yield and Quality? Agronomy. 2023; 13(9):2404. https://doi.org/10.3390/agronomy13092404
Chicago/Turabian StyleVizi, Ramóna, József Kiss, György Turóczi, Nóra Dobra, and Zoltán Pálinkás. 2023. "Turnip Yellows Virus Field Infection in Oilseed Rape: Does It Impact the Yield and Quality?" Agronomy 13, no. 9: 2404. https://doi.org/10.3390/agronomy13092404