
Interacting Effects of Nitrogen Addition and Mowing on Plant Diversity and Biomass of a Typical Grassland in Inner Mongolia
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Sampling
2.3. Data Analysis and Statistics
3. Results
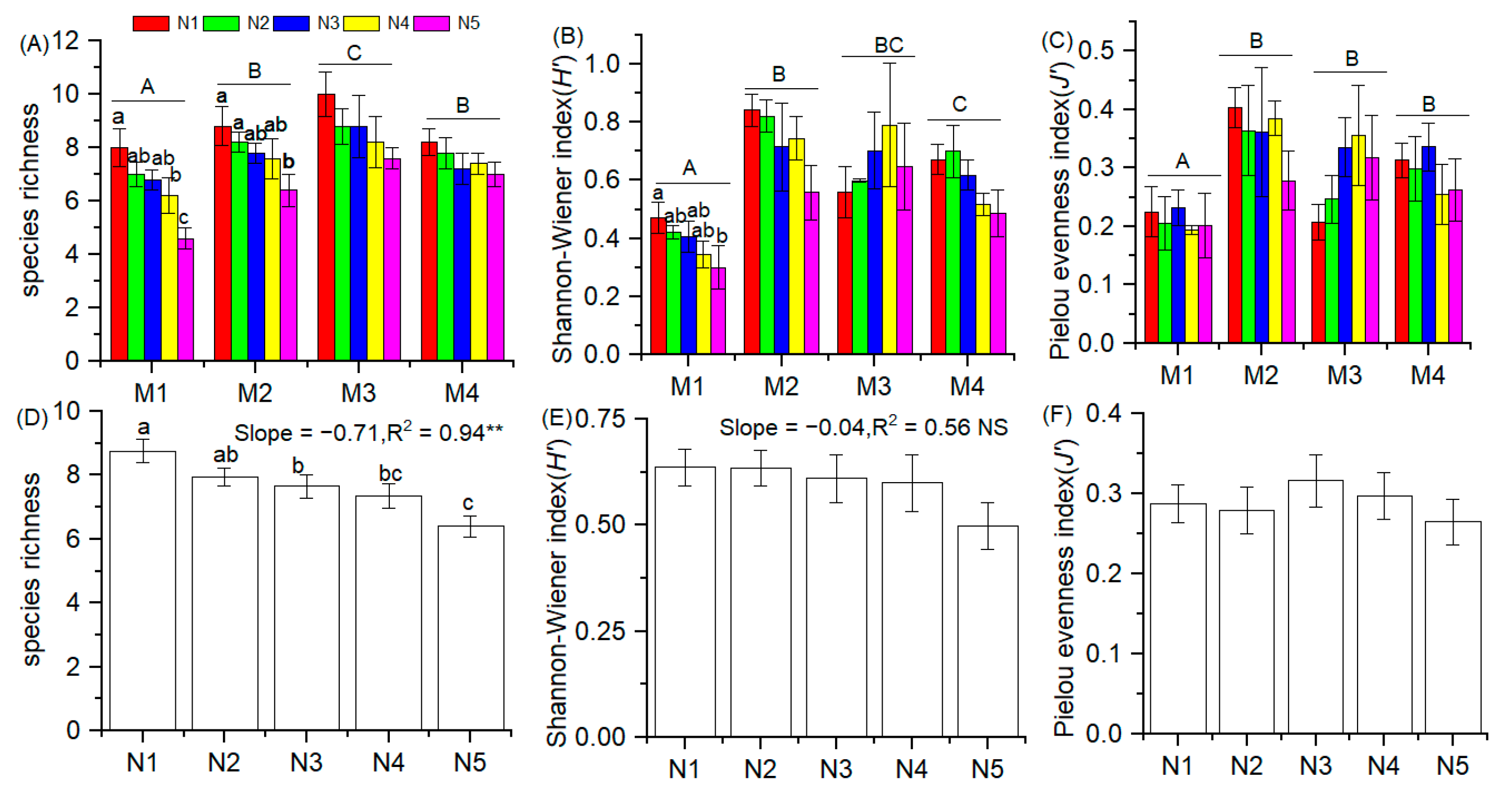
3.1. Plant Diversity
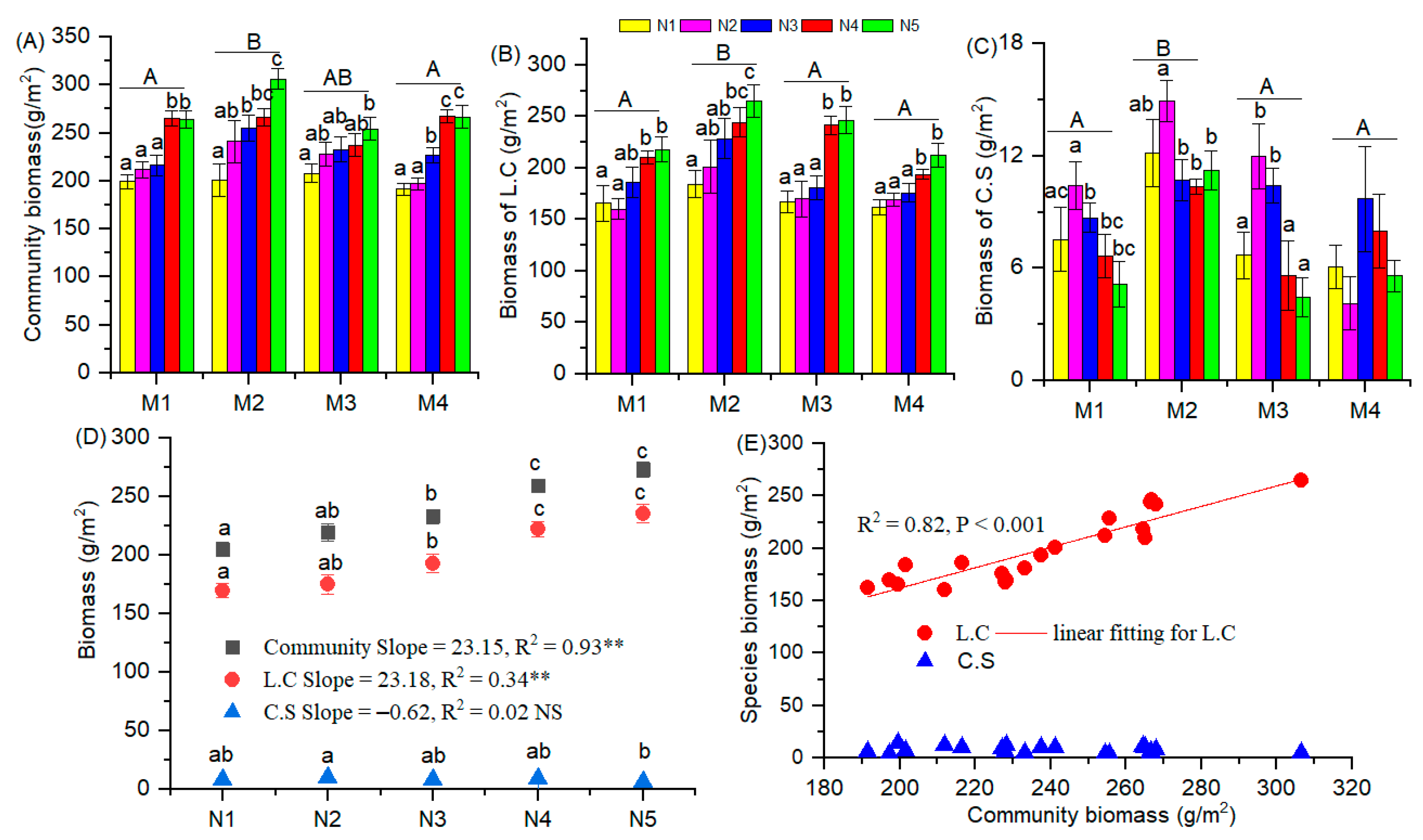
3.2. Biomass
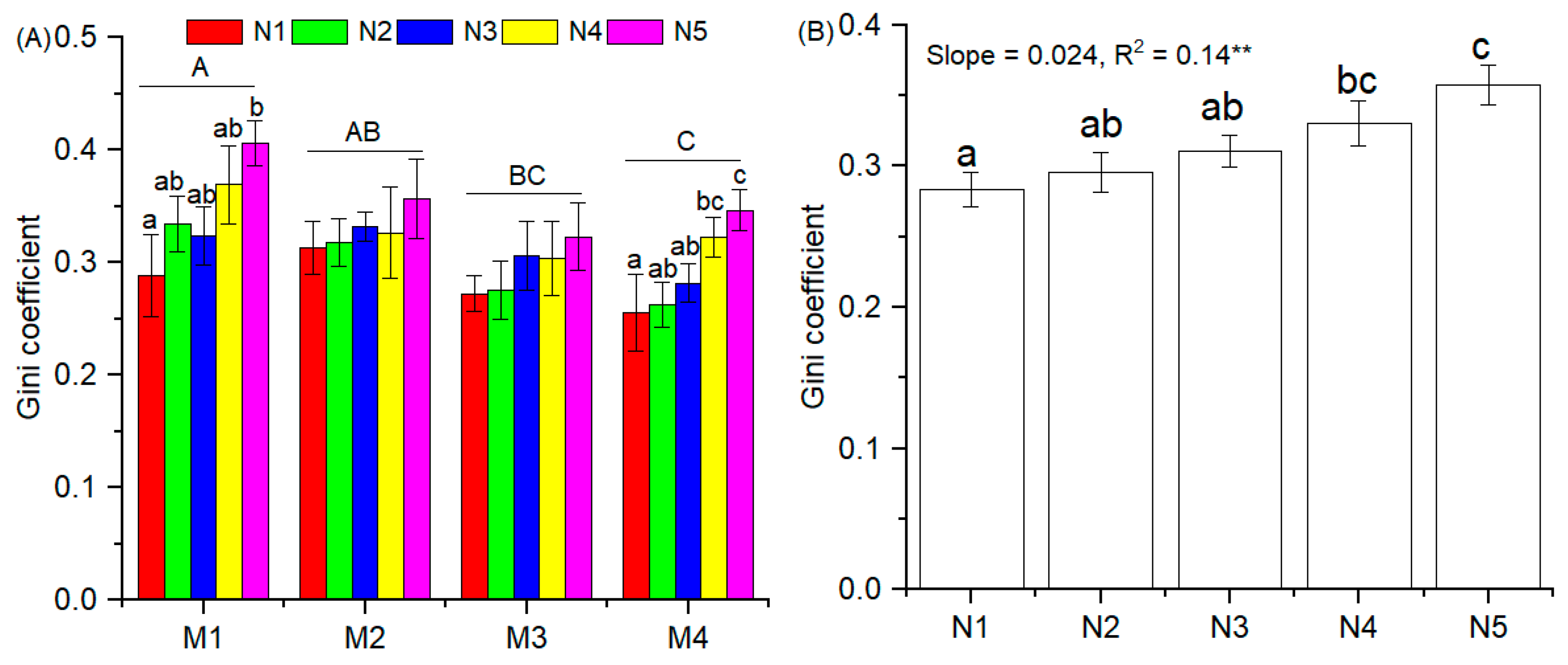
3.3. Gini Coefficient
3.4. Community Density and Litter
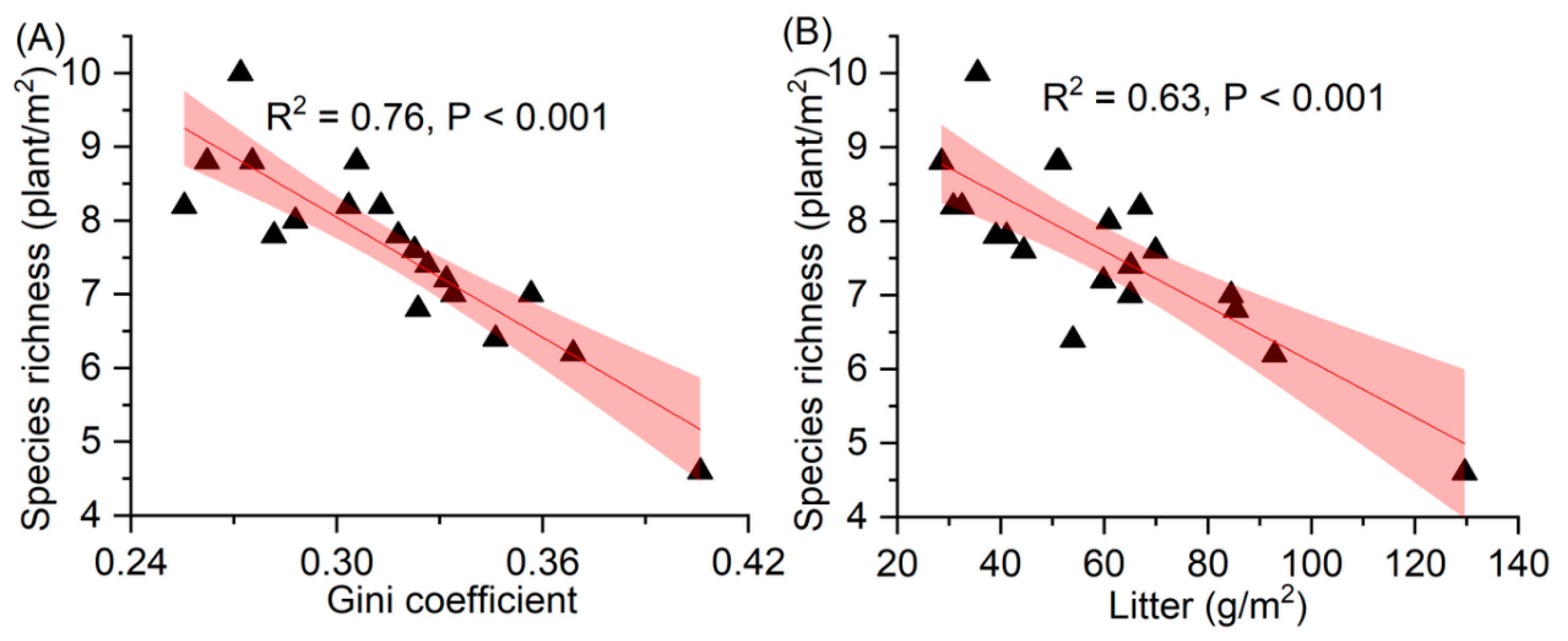
3.5. Relationship between Species Richness and the Gini Coefficient and Litter
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N.; Bleeker, A.; Erisman, J.W. The human creation and use of reactive nitrogen: A global and regional perspective. Annu. Rev. Environ. Resour. 2021, 46, 255–288. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitizinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Pierik, M.; van Ruijven, J.; Bezemer, T.M.; Geerts, R.H.E.M.; Berendse, F. Recovery of plant species richness during long-term fertilization of a species-rich grassland. Ecology 2011, 92, 1393–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scurlock, J.; Hall, D.O. The global carbon sink: A grassland perspective. Glob. Chang. Biol. 1998, 4, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Havstad, K.M.; Peters, D.P.C.; Skaggs, R.; Brown, J.; Bestelmeyer, B.; Fredrickson, E.; Herrick, J.; Wright, J. Ecological services to and from rangelands of the United States. Ecol. Econ. 2007, 64, 261–268. [Google Scholar] [CrossRef]
- Schirpke, U.; Kohler, M.; Leitinger, G.; Fontana, V.; Tasser, E.; Tappeiner, U. Future impacts of changing land-use and climate on ecosystem services of mountain grassland and their resilience. Ecosyst. Serv. 2017, 26, 79–94. [Google Scholar] [CrossRef]
- Chapman, S.K.; Devine, K.A.; Curran, C.; Jones, R.O.; Gilliam, F.S. Impacts of Soil Nitrogen and Carbon Additions on Forest Understory Communities with a Long Nitrogen Deposition History. Ecosystems 2016, 19, 142–154. [Google Scholar] [CrossRef]
- Stevens, C.J.; Lind, E.M.; Hautier, Y.; Harpole, W.S.; Borer, E.T.; Hobbie, S.; Seabloom, E.W.; Ladwig, L.; Bakker, J.D.; Chu, C.J.; et al. Anthropogenic nitrogen deposition predicts local grassland primary production worldwide. Ecology 2015, 96, 1459–1465. [Google Scholar] [CrossRef] [Green Version]
- Tognetti, P.M.; Prober, S.M.; Báez, S.; Chaneton, E.J.; Firn, J.; Risch, A.C.; Schuetz, M.; Simonsen, A.K.; Yahdjian, L.; Borer, E.T.; et al. Negative effects of nitrogen override positive effects of phosphorus on grassland legumes worldwide. Proc. Natl. Acad. Sci. USA 2021, 118, e2023718118. [Google Scholar] [CrossRef]
- Isbell, F.; Reich, P.B.; Tilman, D.; Hobbie, S.E.; Polasky, S.; Binder, S. Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc. Natl. Acad. Sci. USA 2013, 110, 11911–11916. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Yang, L.; Ma, P.; Zhou, H.; Liu, N.; Bai, W.; Wang, H.; Ren, L.; Lu, P.; Han, W.; et al. Below-ground-mediated and phase-dependent processes drive nitrogen-evoked community changes in grasslands. J. Ecol. 2020, 108, 1874–1887. [Google Scholar] [CrossRef]
- Bai, Y.; Wu, J.; Clark, C.M.; Naeem, S.; Pan, Q.; Huang, J.; Zhang, L.; Han, X. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of nitrogen deposition on the species richness of grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbert, J.Y.; Dwyer, J.M.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Chang. Biol. 2016, 22, 110–120. [Google Scholar] [CrossRef]
- Goldberg, D.E.; Miller, T.E. Effects of different resource additions on species diversity in an annual plant community. Ecology 1990, 71, 213–225. [Google Scholar] [CrossRef]
- Huberty, L.E.; Gross, K.L.; Miller, C.J. Effects of nitrogen addition on successional dynamics and species diversity in Michigan old-fields. J. Ecol. 1998, 86, 794–803. [Google Scholar] [CrossRef]
- Zheng, H.P.; Chen, Z.X.; Wang, S.R.; Niu, J.Y. Effects of fertilizer on plant diversity and productivity of desertified alpine grassland at Maqu, Gansu. Acta Pratacult. Sin. 2007, 16, 34–39, (In Chinese with English Abstract). [Google Scholar]
- Yang, Q.; Wang, W.; Zeng, H. Effects of nitrogen addition on the plant diversity and biomass of degraded grasslands of NeiMongol, China. Chin. J. Plant Ecol. 2018, 42, 430–441. [Google Scholar] [CrossRef]
- Hou, S.L.; Yin, J.X.; Sistla, S.; Yang, J.J.; Sun, Y.; Li, Y.Y.; Lü, X.T.; Han, X.G. Long-term mowing did not alter the impacts of nitrogen deposition on litter quality in a temperate steppe. Ecol. Eng. 2017, 102, 404–410. [Google Scholar] [CrossRef]
- Yang, Z.P.; Hugjiltu, M.; Baoyin, T.; Li, F.Y. Plant production decreases whereas nutrients concentration increases in response to the decrease of mowing stubble height. J. Environ. Manag. 2020, 253, 109745. [Google Scholar] [CrossRef]
- Kotas, P.; Choma, M.; Šantrůčková, H.; Lepš, J.; Tříska, J.; Kaštovská, E. Linking above- and belowground responses to 16 years of fertilization, mowing, and removal of the dominant species in a temperate grassland. Ecosystems 2017, 20, 354–367. [Google Scholar] [CrossRef]
- Blüthgen, N.; Dormann, C.F.; Prati, D.; Klaus, V.H.; Kleinebecker, T.; Hölzel, N.; Alt, F.; Boch, S.; Gockel, S.; Hemp, A.; et al. A quantitative index of land-use intensity in grasslands: Integrating mowing, grazing and fertilization. Basic Appl. Ecol. 2012, 13, 207–220. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Socher, S.A.; Prati, D.; Boch, S.; Müller, J.; Klaus, V.H.; Hölzel, N.; Fischer, M. Direct and productivity-mediated indirect effects of fertilization, mowing and grazing on grassland species richness. J. Ecol. 2012, 100, 1391–1399. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, A.P.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [Green Version]
- Foster, B.L.; Gross, K.L. Species richness in a successional grassland: Effects of nitrogen enrichment and plant litter. Ecology 1998, 79, 2593–2602. [Google Scholar] [CrossRef]
- Maron, J.L.; Jefferies, R.L. Restoring enriched grasslands: Effects of mowing on species richness, productivity, and nitrogen retention. Ecol. Appl. 2001, 11, 1088–1100. [Google Scholar] [CrossRef]
- Hiernaux, P.; Turner, M.D. The effect of clipping on growth and nutrient uptake of Sahelian annual rangelands. J. Appl. Ecol. 1996, 33, 387–399. [Google Scholar] [CrossRef]
- Barker, C.G.; Power, S.A.; Bell, J.N.B.; Orme, C.D.L. Effects of habitat management on heathland response to atmospheric nitrogen deposition. Biol. Conserv. 2004, 120, 41–52. [Google Scholar] [CrossRef]
- Li, Z.Q.; Liu, Z.G.; Chen, Z.Z.; Yang, Z.G. The effects of climate changes on the productivity in the Inner Mongolia steppe of China. Acta Pratacult. Sin. 2003, 12, 4–10, (In Chinese with English Abstract). [Google Scholar]
- Pan, Q.M.; Xue, J.G.; Tao, J.; Xu, M.Y.; Zhang, W.H. Current status of grassland degradation and measures for grassland restoration in northern China. Chin. Sci. Bull. 2018, 63, 1642–1650. [Google Scholar] [CrossRef]
- Lü, C.Q.; Tian, H.Q. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. 2007, 112, 10–15. [Google Scholar] [CrossRef]
- Ren, Z.W.; Li, Q.; Chu, C.J.; Zhao, L.Q.; Zhang, J.Q.; Ai, D.X.C.; Yang, Y.B.; Wang, G. Effects of resource additions on species richness and ANPP in an alpine meadow community. J. Plant Ecol. 2010, 3, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Niu, D.C.; Yuan, X.B.; Cease, A.J.; Wen, H.Y.; Zhang, C.P.; Fu, H.; Elser, J.J. The impact of nitrogen enrichment on grassland ecosystem stability depends on nitrogen addition level. Sci. Total Environ. 2017, 618, 1529–1538. [Google Scholar] [CrossRef]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Lü, X.T.; Isbell, F.; Stevens, C.; Han, X.; He, N.P.; Zhang, G.M.; Yu, Q.; Huang, J.H.; Han, X.G. Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Glob. Chang. Biol. 2014, 20, 3520–3529. [Google Scholar] [CrossRef]
- Newman, E.I. Competition and diversity in herbaceous vegetation. Nature 1973, 244, 310. [Google Scholar] [CrossRef]
- Henry, M.; Stevens, H.; Bunker, D.E.; Schnitzer, S.A.; Carson, W.P. Establishment limitation reduces species recruitment and species richness as soil resources rise. J. Ecol. 2004, 92, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. Species richness of experimental productivity gradients: How important is colonization limitation? Ecology 1993, 74, 2179–2191. [Google Scholar] [CrossRef]
- Tian, D.S.; Niu, S.L. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 024019. [Google Scholar] [CrossRef]
- Dickson, T.L.; Foster, B.L. Fertilization decreases plant biodiversity even when light is not limiting. Ecol. Lett. 2011, 14, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.P.; Li, Y.L.; Wang, Y.D.; Cheng, J.W.; Li, F.Y.H. Preferences for different nitrogen forms in three dominant plants in a semi-arid grassland under different grazing intensities. Agric. Ecosyst. Environ. 2022, 333, 107959. [Google Scholar] [CrossRef]
- Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Wilcox, K.R.; Wilson, G.W.T.; Smith, M.D.; Collins, S.L. Changes in plant community composition, not diversity, during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrass prairie. J. Ecol. 2014, 102, 1649–1660. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, L.J.L.; Vergeer, P.; Tim, C.G.R.; Smart, S.M.; Guest, D.; Ashmore, M.R. Direct and indirect effects of nitrogen deposition on species composition change in calcareous grasslands. Glob. Chang. Biol. 2011, 17, 1871–1883. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.L.; Barrett, R.L.; Milner, R.N.C. Annual mowing maintains plant diversity in threatened temperate grasslands. Appl. Veg. Sci. 2017, 12, 3218–3221. [Google Scholar] [CrossRef]
- Socher, S.A.; Prati, D.; Boch, S.; Müller, J.; Baumbach, H.; Gockel, S.; Hemp, A.; Schöning, I.; Wells, K.; Kalko, E.K.V.; et al. Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions. Basic Appl. Ecol. 2013, 14, 126–136. [Google Scholar] [CrossRef]
- Jones, L.; Stevens, C.; Rowe, E.C.; Payne, R.; Caporn, S.J.M.; Evans, C.D.; Field, C.; Dale, S. Can on-site management mitigate nitrogen deposition impacts in non-wooded habitats? Biol. Conserv. 2016, 212, 464–475. [Google Scholar] [CrossRef] [Green Version]
- Lebauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Classen, A.T.; Dukes, J.S.; Kardol, P.; Liu, L.; Luo, Y.; Rustad, L.; Sun, J.; Tang, J.; Templer, P.H.; et al. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol. Lett. 2016, 19, 697–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Chen, S.P.; Lin, G.H. Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecol. 2008, 196, 85–99. [Google Scholar] [CrossRef]
- Liu, Y.; Pan, Q.; Liu, H.; Bai, Y.; Simmons, M.; Dittert, K.; Han, X. Plant responses following grazing removal at different stocking rates in an Inner Mongolia grassland ecosystem. Plant Soil. 2011, 340, 199–213. [Google Scholar] [CrossRef]
- Keddy, P.A.; Twolan-Sstrutt, L.; Shipley, B. Experimental evidence that interspecific competitive asymmetry increases with soil production. Oikos 1997, 80, 253–256. [Google Scholar] [CrossRef]
- Yang, G.J.; Hautier, Y.; Zhang, Z.J.; Lü, X.T.; Han, X.G. Decoupled responses of above- and below-ground stability of productivity to nitrogen addition at the local and larger spatial scale. Glob. Chang. Biol. 2022, 28, 2711–2720. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Chang, S.X.; Cai, Z.; Mueller, C.; Zhang, J. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review. Environ. Pollut. 2019, 244, 608–616. [Google Scholar] [CrossRef]
- Mikola, J.; Setälä, H.; Virkajärvi, P.; Saarijärvi, K.; Ilmarinen, K.; Voigt, W.; Vestberg, M. Defoliation and patchy nutrient return drive grazing effects on plant and soil properties in a dairy cow pasture. Ecol. Monogr. 2009, 79, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Houle, D.; Moore, J.D.; Ouimet, R.; Marty, C. Tree species partition N uptake by soil depth in boreal forests. Ecology 2014, 95, 1127–1133. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | ||||||||
|---|---|---|---|---|---|---|---|---|
| Whole Plot | Sub Plot | |||||||
| B | M | N | M * N | |||||
| df (4, 12) | df (3, 12) | df (4, 64) | df (12, 64) | |||||
| F-Test | p | F-Test | p | F-Test | p | F-Test | p | |
| Species richness | 1.03 | 0.43 | 4.41 | 0.03 | 10.63 | 0.00 | 0.52 | 0.89 |
| Shannon–Wiener index | 0.07 | 0.99 | 11.68 | <0.001 | 1.45 | 0.23 | 0.80 | 0.65 |
| Pielou evenness index | 0.17 | 0.95 | 4.85 | 0.02 | 0.50 | 0.73 | 0.69 | 0.75 |
| Community biomass | 0.21 | 0.93 | 6.21 | 0.01 | 26.24 | 0.00 | 2.29 | 0.02 |
| Litter production | 0.51 | 0.74 | 145.12 | <0.001 | 66.00 | <0.001 | 3.92 | <0.001 |
| L. chinensis biomass | 0.97 | 0.46 | 6.20 | 0.01 | 16.14 | 0.00 | 1.04 | 0.43 |
| C. squarrosa biomass | 0.40 | 0.81 | 7.96 | 0.00 | 8.78 | 0.00 | 2.98 | 0.00 |
| Gini coefficient | 1.60 | 0.32 | 5.52 | 0.01 | 4.56 | 0.01 | 0.40 | 0.96 |
| Community density | 10.39 | 0.02 | 8.69 | 0.00 | 14.27 | 0.00 | 0.61 | 0.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Minggagud, H.; Wang, Q.; Pan, H. Interacting Effects of Nitrogen Addition and Mowing on Plant Diversity and Biomass of a Typical Grassland in Inner Mongolia. Agronomy 2023, 13, 2125. https://doi.org/10.3390/agronomy13082125
Yang Z, Minggagud H, Wang Q, Pan H. Interacting Effects of Nitrogen Addition and Mowing on Plant Diversity and Biomass of a Typical Grassland in Inner Mongolia. Agronomy. 2023; 13(8):2125. https://doi.org/10.3390/agronomy13082125
Chicago/Turabian StyleYang, Zhaoping, Hugjiltu Minggagud, Qian Wang, and Hongyuan Pan. 2023. "Interacting Effects of Nitrogen Addition and Mowing on Plant Diversity and Biomass of a Typical Grassland in Inner Mongolia" Agronomy 13, no. 8: 2125. https://doi.org/10.3390/agronomy13082125