3.2. Effect of Different Rootstocks on Organic Acid Content in ‘Cabernet Sauvignon’ Grapes
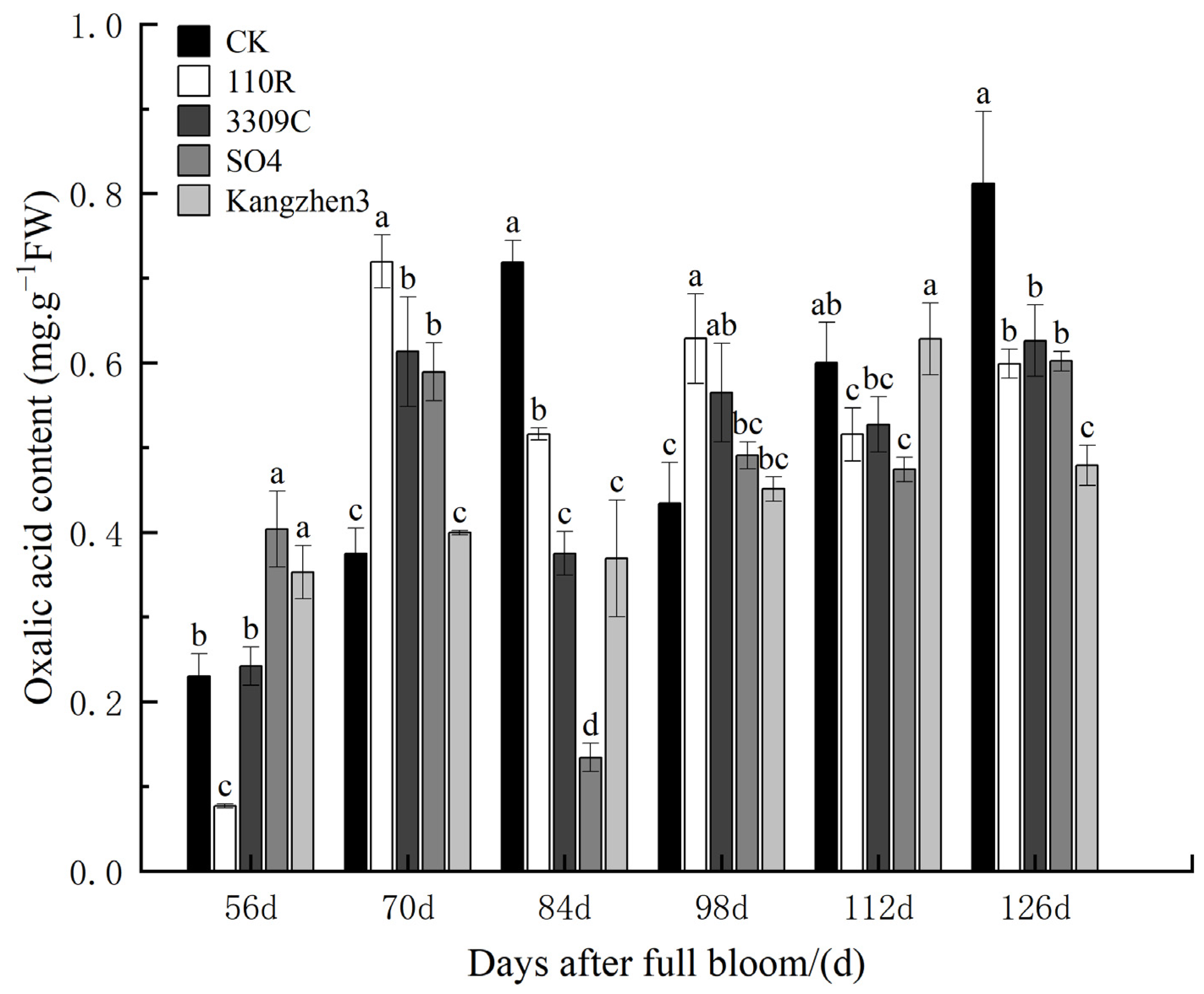
The dynamic changes in the impact of different rootstocks on the oxalic acid content in ‘Cabernet Sauvignon’ grapes are shown in
Figure 2. The trend of oxalic acid content in the fruit between the different treatments exhibited a consistent pattern, initially increasing, then decreasing, and ultimately increasing again. The oxalic acid content at the mature stage was greater than that before the color conversion stage, and the oxalic acid content did not change significantly from flowering to maturity. At 56 d after anthesis, the oxalic acid content in the fruit was 0.35 mg.g
−1 FW and 0.40 mg.g
−1 FW for the CS169/Kangzhen3 and CS169/SO4 rootstock–scion combinations, respectively. These values were significantly higher than those of the control (CK) by 75.76% and 53.62%, respectively. However, in the CS169/110R rootstock–scion combination, the oxalic acid content of the fruit was 0.08 mg.g
−1 FW, which was significantly lower than that of CK (66.65%). At 70 d after anthesis, the oxalic acid content in the fruit of the CS169/110R, CS169/3309C, and CS169/SO4 rootstock–scion combinations was significantly higher than that of CK (91.99%, 63.65%, and 57.23%, respectively). The difference between the CS169/Kangzhen3 rootstock–scion combination and CK was small. At 84 d after anthesis, the oxalic acid content in the fruits of each experimental rootstock–scion combination was significantly lower than that of CK. At 98 d after anthesis, the fruit of the CS169/110R and CS169/3309C rootstock–scion combinations had oxalic acid contents of 0.63 mg.g
−1 FW and 0.56 mg.g
−1 FW, respectively. These values were significantly higher than those of CK (44.82% and 30.09%, respectively). At 112 d after anthesis, the oxalic acid content in the fruits of CS169/110R and CS169/SO4 was 0.52 mg.g
−1 FW and 0.47 mg.g
−1 FW, respectively, while CS169/3309C and CS169/Kangzhen3, had significantly lower oxalic acid contents compared to CK by 14.12% and 12.16%, respectively. At 126 d after anthesis, the oxalic acid content in the fruits of each rootstock–scion combination was significantly lower than that of CK. According to comprehensive analysis, using materials such as 110R, SO4, 3309C, and Kangzhen3 as rootstocks, with ‘Cabernet Sauvignon’ self- rooted seedlings as control, can significantly reduce the oxalic acid content in mature fruits.
The dynamic changes in the influence of different rootstocks on the tartaric acid content in ‘Cabernet Sauvignon’ fruits are shown in
Figure 3. Regardless of the rootstock used (110R, 3309C, SO4, Kangzhen3) or ‘Cabernet Sauvignon’ self-rooted seedlings, the tartaric acid content in the fruit exhibited a similar trend across different treatments. It initially showed a downward trend and underwent significant changes from flowering to maturity. At 56 d after anthesis, the tartaric acid content in the fruits of the CS169/110R and CS169/3309C rootstock–scion combinations was significantly lower than that of CK (10.25% and 7.75%, respectively), but this was not the case for CS169/Kangzhen3. At 70 d after anthesis, the tartaric acid content in the fruits of the CS169/110R and CS169/SO4 rootstock–scion combinations was significantly higher than that of CK by 43.28% and 22.21%, respectively. The tartaric acid content in the fruits of the CS169/3309C and CS169/Kangzhen3 rootstock–scion combinations was significantly lower than that of CK by 27.82% and 26.55%, respectively. At 84 d after anthesis, the tartaric acid content in the fruits of the CS169/110R and CS169/3309C rootstock–scion combinations was significantly lower than that of CK by 6.28% and 16.94%, respectively. The CS169/Kangzhen3 rootstock–scion combination was significantly higher than CK by 9.08%, with a small difference from the CS169/SO4 rootstock–scion combination. In contrast to the CS169/3309C rootstock–scion combination, the tartaric acid content in the fruit of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations was significantly higher than that of CK by 31.34%, 10.06%, and 4.74%, respectively, at 98 d after anthesis. At 112 d after anthesis, the tartaric acid content in the fruits of CS169/110R, CS169/3309C, and CS169/Kangzhen3 was significantly higher than that of CK by 5.22%, 7.61%, and 8.00%, respectively. At 126 d after anthesis, the tartaric acid content in the fruits of CS169/3309C was slightly higher than that in the fruits of CK. The rootstock–scion combinations CS169/110R, CS169/SO4, and CS169/Kangzhen3 were significantly higher than those of CK by 9.03%, 6.30%, and 12.74%, respectively. Throughout the experimental period, the rootstock–scion combinations of CS169/110R, CS169/SO4, and CS169/Kangzhen3 significantly increased the tartaric acid content in the fruits of ‘Cabernet Sauvignon’ grapes to varying degrees.
The dynamic changes in the effect of different rootstocks on the malic acid content in ‘Cabernet Sauvignon’ grapes are shown in
Figure 4. The variation trend of the malic acid content in the fruits of the different treatments was generally the same, showing a dynamic downward trend. The malic acid content in the fruits of different rootstock–scion combinations changed significantly from flowering to maturity. At 56 d after anthesis, in contrast to the fruit of the CS169/3309C rootstock–scion combination, the malic acid content of CS169/110R, CS169/SO4 and CS169/Kangzhen3 was significantly higher than CK by 45.83%, 12.15%, and 17.51%, respectively. At 70 d after anthesis, the malic acid content in the fruits of CS169/110R, CS169/SO4, and CS169/Kangzhen3 was significantly higher than that of CK by 17.17%, 8.61%, and 5.67%, respectively, whereas that of the CS169/3309C rootstock–scion combination was significantly lower than that of CK (14.66%). At 84 d after anthesis, the malic acid content in the fruits of CS169/110R, CS169/SO4, and CS169/Kangzhen3 was significantly higher than that of CK (14.09%, 16.34%, and 6.23%, respectively), whereas that of the CS169/3309C rootstock–scion combination was significantly lower than that of CK (15.32%). At 98 d after anthesis, the malic acid content in the fruits of CS169/110R, CS169/SO4, and CS169/Kangzhen3 was significantly higher than that of CK by 16.38%, 5.31%, and 13.11%, respectively, whereas that of the CS169/3309C rootstock–scion combination was significantly lower than that of CK by 26.16%. At 112 and 126 d after anthesis, the content of the CS169/3309C rootstock–scion combination was slightly lower than that of CK, whereas that of CS169/110R, CS169/SO4, and CS169/Kangzhen3 was significantly higher than that of CK. Comprehensive analysis showed that the rootstock–scion combination of CS169/110R, CS169/SO4, and CS169/Kangzhen3 significantly increased malic acid content in ‘Cabernet Sauvignon’ grapes at different stages after anthesis.
The dynamic changes in the influence of different rootstocks on the citric acid content in ‘Cabernet Sauvignon’ fruit are shown in
Figure 5. The citric acid content in the fruits of the different treatments exhibited a consistent trend of initial decrease, followed by an increase and then another decrease. The citric acid content changed significantly from flowering to maturity. During the entire experimental period, there were differences between the rootstock–scion combinations at different stages and the ‘Cabernet Sauvignon’ self-rooted seedlings. At 56 d after anthesis, the citric acid content of all combinations, except the CS169/110R rootstock–scion combination, was significantly higher than that of CK. At 70 d after anthesis, there was no significant difference between the citric acid content and CK in the fruits of each rootstock–scion combination. At 84 d after anthesis, the citric acid content in the fruits of each rootstock–scion combination was significantly lower than that of CK, with CS169/3309C and CS169/SO4 being 51.79% and 51.68% lower, respectively. At 98 d after anthesis, the citric acid content in the fruits of CS169/110R and CS169/3309C was significantly higher than that of CK 64.07% and 33.90%, with a small difference compared to other rootstock–scion combinations. At 112 d after anthesis, the citric acid content in the fruits of CS169/110R and CS169/3309C was significantly lower than that of CK (36.03% and 26.80%, respectively). The rootstock–scion combinations of CS169/SO4 and CS169/Kangzhen3 were significantly higher than those of CK (24.23% and 46.88%, respectively). At 126 d after anthesis, the citric acid content in the fruits of each rootstock–scion combination was significantly lower than that of CK. Comprehensive analysis showed that using materials such as 110R, SO4, 3309C, and Kangzhen3 as rootstocks, with ‘Cabernet Sauvignon’ self-rooted seedlings as control, can significantly reduce the citric acid content in the fruit to varying degrees.
The dynamic changes in the impact of different rootstocks on succinic acid content in ‘Cabernet Sauvignon’ grapes are shown in
Figure 6. The differences in the succinic acid content of the fruits between the different treatments were significant. During the entire experimental period, there were differences between rootstock–scion combinations at different stages and between ‘Cabernet Sauvignon’ self-rooted seedlings. At 56 d after anthesis, the succinic acid content in the fruits of the CS169/3309C and CS169/Kangzhen3 rootstock–scion combinations was slightly different from that of CK. The CS169/110R rootstock–scion combination was 30.42% lower than CK, and the CS169/SO4 rootstock–scion combination was 62.35% higher than CK. At 70 d after anthesis, the succinic acid content in the fruit of the CS169/SO4 rootstock–scion combination was slightly different from that of CK, with that of CS169/3309C and CS169/Kangzhen3 being significantly lower than that of CK (57.77% and 56.54%, respectively) and was significantly higher in CS169/110R than of CK (37.71%). At 84 d after anthesis, the succinic acid content in the fruit of CS169/110R was significantly lower than that of CK by 22.14%, and the other rootstock–scion combinations showed little difference from CK. At 98 d after anthesis, there was a small difference in succinic acid content between each rootstock–scion combination and the CK fruit. At 112 d after anthesis, the succinic acid content in the fruit of CS169/3309C was significantly lower than that of CK by 34.37%, and that of CS169/Kangzhen3 was significantly higher than that of CK by 46.84%. The differences among CS169/110R, CS169/SO4, and CK were relatively small. At 126 d after anthesis, the succinic acid content in the fruit of CS169/SO4 was slightly higher than that of CK, and that of the other rootstock–scion combinations was significantly higher than that of CK. According to a comprehensive analysis, utilizing rootstocks such as 110R, SO4, 3309C, and Kangzhen3, with ‘Cabernet Sauvignon’ self-rooted seedlings as a control, demonstrated the potential to enhance the content of succinic acid in ‘Cabernet Sauvignon’ grapes to varying degrees.
Dynamic changes in the impact of different rootstocks on the total accumulation of organic acids in ‘Cabernet Sauvignon’ grapes are shown in
Figure 7. The accumulation of organic acids in the CS169/3309C fruit was lower than that in the CK fruit throughout the experimental period. With the exception of 70 d after anthesis, the acid accumulation in the fruits of the CS169/110R, CS169/Kangzhen3, and CS169/SO4 rootstock–scion combinations was significantly higher than that of CK during the remaining color conversion stages. This indicates that CS169/110R, CS169/Kangzhen3, and CS169/SO4 rootstock–scion combinations could significantly increase the organic acid content in ‘Cabernet Sauvignon’ grapes at different stages after anthesis.
3.3. Effect of Different Rootstocks on the Activity of Organic Acid Metabolism Enzymes in ‘Cabernet Sauvignon’ Grapes
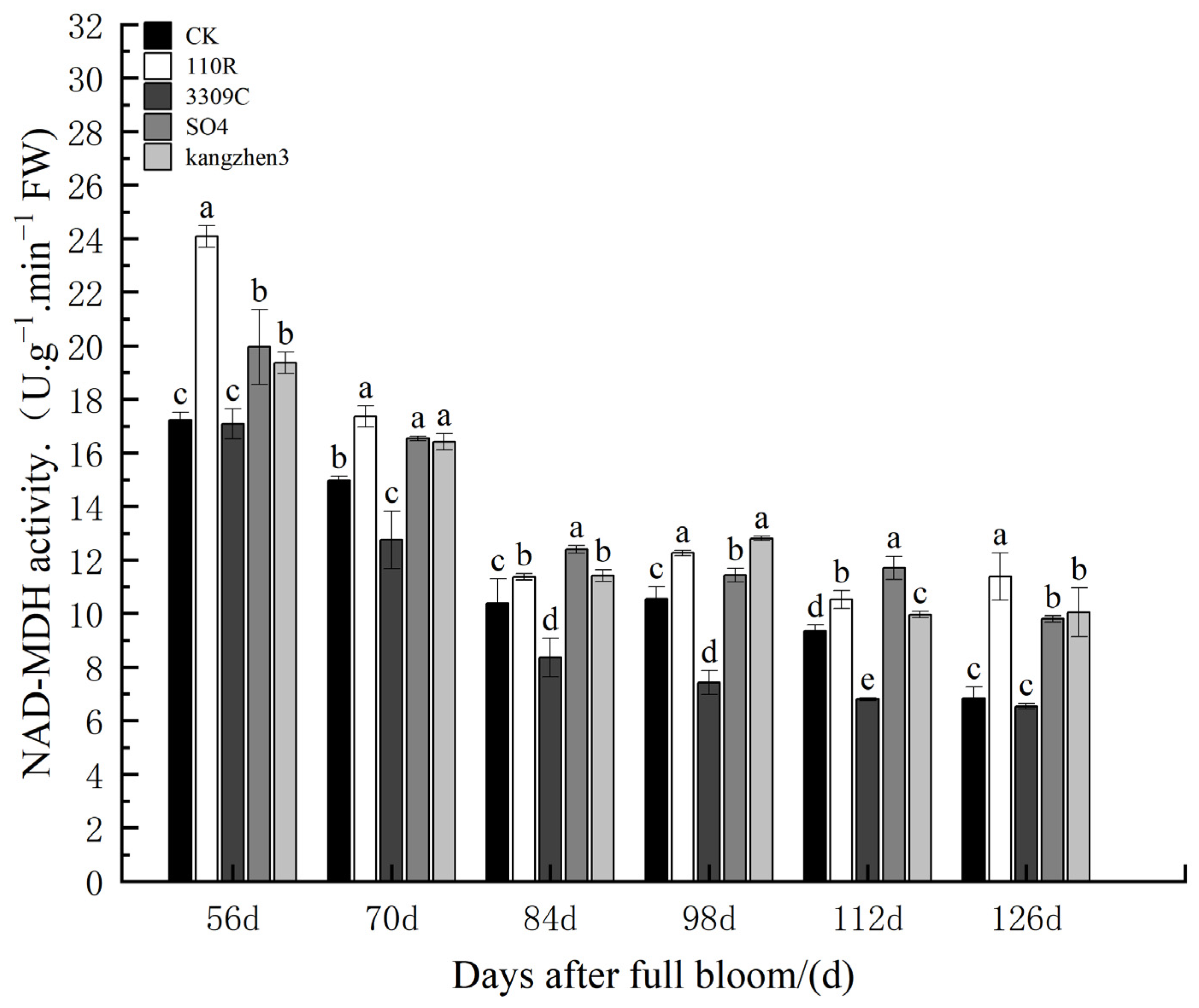
The NAD-MDH enzyme activity in ‘Cabernet Sauvignon’ grapes grown on different rootstocks is shown in
Figure 8. The trend of change in NAD-MDH enzyme activity in fruits under different treatments was generally the same, showing a dynamic and violent decline, followed by a relatively stable rise and decline. NAD-MDH enzyme activity changed significantly from flowering to maturity. During the entire experimental period, there were differences between the rootstock–scion combinations at different stages and the ‘Cabernet Sauvignon’ self-rooted seedlings. At 56 d after anthesis, the NAD-MDH enzyme activity in the fruits of the CS169/SO4 and CS169/Kangzhen3 rootstock–scion combinations was significantly higher than that of CK, with differences of 11.48% and 19.28%, respectively. Similarly, the CS169/110R rootstock–scion combination showed a substantial difference from CK, displaying an increase of 46.90% in NAD-MDH enzyme activity. However, the CS169/3309C rootstock–scion combination demonstrated a minor difference in NAD-MDH enzyme activity compared to CK. At 70 d after anthesis, the NAD-MDH enzyme activity in the fruits of CS169/3309C was significantly lower than that of CK by 19.25%, and the spike combinations of CS169/110R, CS169/SO4, and CS169/Kangzhen3 were significantly higher than that of CK by 12.64%, 4.78%, and 3.94%, respectively. At 84 d after anthesis, NAD-MDH enzyme activity in the fruit of the CS169/SO4 rootstock–scion combination was significantly higher than that of CK by 19.04%, whereas that of CS169/110R and CS169/Kangzhen3 was significantly higher than that of CK by 9.54% and 6.41%, respectively. Other rootstock–scion combinations showed little difference compared to CK. At 98 d after anthesis, the NAD-MDH enzyme activity in the fruit of the CS169/3309C rootstock–scion combination was significantly lower than that of CK by 29.55%, that of the CS169/Kangzhen3 combination was significantly higher than that of CK by 21.43%, and that of the CS169/110R and CS169/SO4 rootstock–scion combinations was significantly higher than that of CK by 16.37% and 8.44%, respectively. At 112 d after anthesis, the NAD-MDH enzyme activity in the fruit of CS169/3309C was significantly lower than that of CK, the NAD-MDH enzyme activity in the fruit of CS169/SO4 was significantly higher than that of CK, and that of the combination of CS169/110R and CS169/Kangzhen3 was significantly lower than that of CK. At 126 d after anthesis, NAD-MDH enzyme activity in the fruits of CS169/110R, CS169/SO4, and CS169/Kangzhen3 was significantly higher than that of CK, whereas that of the CS169/3309C rootstock–scion combination was slightly lower than that of CK. Comprehensive analysis showed that using 110R, SO4, and Kangzhen3 as rootstocks improved the NAD-MDH enzyme activity of ‘Cabernet Sauvignon’ grapes to varying degrees.
The effects of the different rootstocks on NADP-ME enzyme activity in ‘Cabernet Sauvignon’ grapes are shown in
Figure 9. The overall trend of the changes in NADP-ME enzyme activity in the fruits of the different treatments was the same, showing a dynamic upward trend. NADP-ME enzyme activity changed significantly from flowering to maturity. At 56 d after flowering, the NADP-ME enzyme activity in the fruits of the CS169/110R and CS169/3309C rootstock–scion combinations was significantly higher than that of CK by 23.65% and 5.37%, respectively, whereas the CS169/SO4 and CS169/Kangzhen3 rootstock–scion combinations were significantly lower than CK by 6.65% and 15.58%, respectively. At 70 d after anthesis, the NADP-ME enzyme activity in the fruits of the same CS169/110R and CS169/3309C rootstock–scion combinations was significantly higher than that of CK, whereas the activities of CS169/SO4 and CS169/Kangzhen3 rootstock–scion combinations were significantly lower than that of CK. At 84 d after anthesis, NADP-ME enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK. At 98 d after flowering, NADP-ME enzyme activity in the fruit of CS169/3309C was significantly higher than that of CK by 12.02%, and that of CS169/110R was significantly lower than that of CK by 17.23%. The differences among CS169/SO4, CS169/Kangzhen3, and CK were relatively small. At 112 d after anthesis, the NADP-ME enzyme activity in the fruits of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations was significantly lower than that of CK by 6.83%, 20.19%, and 19.28%, respectively, whereas that of CS169/3309C was significantly higher than that of CK. At 126 d after flowering, the NADP-ME enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK. The overall analysis showed that, with the exception of using 110R, SO4, and Kangzhen3 materials as rootstocks at 56 and 70 d, with ‘Cabernet Sauvignon’ self-rooted seedlings as controls, all other treatments resulted in a varying decrease in NADP-ME enzyme activity in ‘Cabernet Sauvignon’ grapes.
The effects of the different rootstocks on PEPC enzyme activity in ‘Cabernet Sauvignon’ grapes are shown in
Figure 10. The overall trend of PEPC enzyme activity in fruits under different treatments was the same, showing a dynamic downward trend. At 56 d after flowering, the PEPC enzyme activities of the CS169/110R, CS169/3309C, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations were significantly lower than that of CK by 21.50%, 14.24%, 7.98%, and 42.60%, respectively. At 70 d after flowering, PEPC enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK. At 84 d after anthesis, the PEPC enzyme activity in the fruits of the CS169/110R, CS169/3309C, and CS169/SO4 rootstock–scion combinations was significantly lower than that of CK by 23.81%, 17.15%, and 5.04%, respectively. There was a small difference between CS169/Kangzhen3 and CK. At 98 d after flowering, PEPC enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK. At 112 d after flowering, the rootstock–scion combinations CS169/110R, CS169/3309C, and CS169/Kangzhen3 were significantly lower than CK, and there was no difference in PEPC enzyme activity between CK fruit and CS169/SO4. At 126 d after anthesis, PEPC enzyme activity in the CK fruit was significantly lower than that of CS169/SO4 and significantly higher than that of CS169/110R, CS169/3309C, and CS169/Kangzhen3 rootstock–scion combinations. Overall analysis showed that the use of 110R, SO4, 3309C, and Kangzhen3 materials as rootstocks, with ‘Cabernet Sauvignon’ self-rooted seedlings as a control, resulted in a varying decrease in PEPC enzyme activity in ‘Cabernet Sauvignon’ fruit.
The effects of different rootstocks on the CS enzyme activity in ‘Cabernet Sauvignon’ grapes are shown in
Figure 11. The changes in CS enzyme activity in the fruits of the different treatments were not the same and showed dynamic changes. The CS enzyme activity changed significantly from flowering to maturity. At 56 d after flowering, the CS enzyme activity of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 different rootstock–scion combinations was significantly lower than that of CK (62.76%, 55.20%, and 71.72%, respectively), but that of the CS169/3309C rootstock–scion combination was significantly higher than that of CK (44.84%). At 70 d after flowering, the CS enzyme activity in the fruit of the CS169/110R rootstock–scion combination was significantly higher than that of CK by 12.77%. However, all other rootstock–scion combinations showed significantly lower CS enzyme activity compared to CK. At 84 d after flowering, CS enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK. At 98 d after flowering, CS enzyme activity in the fruits of CS169/110R and CS169/Kangzhen3 was significantly lower than that of CK by 25.18% and 77.33%, respectively. The difference between the CS169/SO4 rootstock–scion combination and CK was relatively small. At 112 and 126 d after flowering, CS enzyme activity in the fruits of each rootstock–scion combination was significantly lower than that of CK.
3.4. Effects of Different Rootstocks on the Gene Expression of Key Enzymes for Organic Acid Metabolism in ‘Cabernet Sauvignon’ Grapes
The effects of different rootstocks on the relative expression of NAD-MDH in ‘Cabernet Sauvignon’ grapes are shown in
Figure 12. The trend of changes in the combination of rootstock–scion at different stages was the same, and there were differences in the relative expression of the NAD-MDH gene in fruits among the different treatments. At 56 d after flowering, the relative expression of the NAD-MDH gene in the fruit of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 different rootstock–scion combinations was significantly higher than that of CK (41.76%, 12.18%, and 19.40%, respectively). The difference between the CS169/3309C rootstock–scion combination and CK was small. Seventy days after flowering, the relative expression level of the NAD-MDH gene in the fruits of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations was also significantly higher than that of CK, but the CS169/3309C rootstock–scion combination was significantly lower than that of CK by 17.15%. At 84 d after flowering, the expression level of the NAD-MDH gene in the fruit of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations was significantly higher than that of CK, whereas that of the CS169/3309C rootstock–scion combination was significantly lower than that of CK, by 22.53%. At 98 d after flowering, the expression of NAD-MDH in the fruit of the CS169/3309C rootstock–scion combination was significantly lower than that of CK by 22.46%. The CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations were significantly lower than CK by 26.89%, 5.06%, and 12.51%, respectively. At 112 d after flowering, the expression level of the NAD-MDH gene in the fruit of CS169/3309C was significantly lower than that of CK by 31.93%, and that of CS169/110R was significantly higher than that of CK by 28.89%. The difference between the CS169/SO4 and CS169/Kangzhen3 rootstock–scion combinations and CK was small. At 126 d after flowering, the NAD-MDH gene expression in the CS169/3309C rootstock–scion combination fruit was slightly lower than that of CK. However, the CS169/110R, CS169/SO4, and CS169/Kangzhen3 rootstock–scion combinations showed significantly higher NAD-MDH gene expression compared to CK by 55.83%, 32.87%, and 44.14%, respectively. Overall, when materials such as 110R, SO4, and Kangzhen3 were used as rootstocks, the relative gene expression of NAD-MDH in ‘Cabernet Sauvignon’ grapes under the rootstock–scion combination was significantly higher than that of CK, indicating a good correlation with NAD-MDH.
The relative expression level of the NADP-ME gene in ‘Cabernet Sauvignon’ grapes under different rootstocks is shown in
Figure 13. There were differences in the relative expression levels of the NADP-ME gene in fruits under different treatments. At 56 d after flowering, the relative expression level of the NADP-ME gene in the fruits of the CS169/110R, CS169/SO4, and CS169/Kangzhen3 different rootstock–scion combinations was significantly higher than that of CK. The difference between the CS169/3309C rootstock–scion combination and CK was small. At 70 d after flowering, the relative expression levels of NADP-ME genes in fruits of CS169/110R and CS169/Kangzhen3 were significantly lower than those of CK (26.20% and 14.93%, respectively), whereas the relative expression levels of NADP-ME genes in fruits of CS169/3309C and CS169/SO4 rootstock–scion combinations were significantly higher than those of CK (42.84% and 28.00%, respectively). After 84 d of flowering, the expression level of the NADP-ME gene in the fruit of the CS169/110R, CS169/3309C, and CS169/SO4 different rootstock–scion combinations was significantly lower than that of CK, but that of CS169/Kangzhen3 was significantly higher than that of CK. At 98 d after flowering, the expression level of the NADP-ME gene in the fruit of CS169/3309C was significantly lower than that of CK by 16.45%. The expression level of the NADP-ME gene in CS169/110R and CS169/SO4 was significantly higher than that of CK by 62.92% and 13.94%, respectively. The difference between the CS169/Kangzhen3 rootstock–scion combination and CK was relatively small. At 112 d after flowering, the expression level of the NADP-ME gene in the CS169/110R and CS169/3309C rootstock–scion combinations were significantly lower than that of CK, whereas that of CS169/Kangzhen3 was significantly higher than that of CK. The difference between the CS169/SO4 rootstock–scion combination and CK was small. At 126 d after flowering, the expression level of the NADP-ME gene in the fruit of the CS169/3309C rootstock–scion combination was significantly higher than that of CK by 10.34%, whereas that of CS169/110R and CS169/Kangzhen3 was significantly lower than that of CK by 10.49% and 53.04%, respectively. The differences between the CS169/SO4 and CK treatments were small.
The expression level of the PEPC gene in ‘Cabernet Sauvignon’ grapes under different rootstocks is shown in
Figure 14. There were certain differences in the expression level of the PEPC gene in fruits among different treatments. At 56 d after flowering, except for a small difference in PEPC gene expression between the CS169/3309 rootstock–scion combination and CK, all other rootstock–scion combinations were significantly higher than CK. At 70 d after flowering, the expression level of the PEPC gene in the fruit of CS169/110R and CS169/Kangzhen3 different rootstock–scion combinations was significantly higher than that of CK (14.16% and 19.10%, respectively), while that of CS169/3309C was significantly lower than that of CK (22.65%). The differences between the CS169/SO4 and CK treatments were small. At 84 d after flowering, except for CS169/3309C, which showed a significantly lower expression level of the PEPC gene compared to CK by 44.34%, the fruits of CS169/110R, CS169/SO4, and CS169/Kangzhen3 different rootstock–scion combinations exhibited significantly higher expression levels of the PEPC gene than CK. At 98 d after flowering, the expression level of the PEPC gene in the fruit of the CS169/110R, CS169/3309C, and CS169/Kangzhen3 rootstock–scion combinations was significantly lower than that of CK, while the expression level of the PEPC gene in the fruit of the CS169/SO4 rootstock–scion combination was significantly higher than that of CK. At 112 d after flowering, the expression level of the PEPC gene in the fruit of CS169/110R, CS169/3309C, and CS169/Kangzhen3 different rootstock–scion combinations was significantly lower than that of CK (62.38%, 33.29%, and 31.97%, respectively). The expression level of the PEPC gene in the fruit of CS169/SO4 rootstock–scion combination was significantly higher than that of CK (44.63%). At 126 d after flowering, the expression level of the PEPC gene in the fruit of the CS169/110R and CS169/Kangzhen3 different rootstock–scion combinations was significantly lower than that of CK by 28.49% and 14.95%, respectively. However, the expression level of the PEPC gene in the fruit of the CS169/SO4 rootstock–scion combination was significantly higher than that of CK by 26.15%. Moreover, the difference between the CS169/3309C rootstock–scion combination and CK was small.
During the analysis of the key enzyme genes for organic acid metabolism in ‘Cabernet Sauvignon’ grapes, it was found that the gene expression of NAD-MDH, NADP-ME, and PEPC in the self-rooted seedlings of ‘Cabernet Sauvignon’ grapes as controls remained almost unchanged at different stages of the experiment. However, the expression of these three genes in ‘Cabernet Sauvignon’ grapes under the action of rootstocks 110R, 3309C, SO4, and Kangzhen3 varied.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}