
Assessing Fruit Maturity and Quality of ‘Buckeye Gala’ Grown on a Diverse Panel of Apple (Malus domestica Borkh.) Rootstocks in Western Maryland

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Tree Performance Measurements
2.3. Fruit Internal Ethylene Concentration
2.4. Fruit Physicochemical Measurements
2.5. Climate Data
2.6. Statistical Analysis
3. Results
3.1. Climactological Parameters during the Two Production Seasons of the Study
3.2. Effect of Roostock Genotypes on ‘Buckeye Gala’ Tree Size, Yield and Crop Load
3.3. Internal Ethylene Concentration and Physicochemical Properties of ‘Buckeye Gala’ Apple Fruit Grafted on Ten Different Rootstocks
3.4. Relationships among Tree Perdomance Measurements, Ethylene Concentration and Physicochemical Properties of ‘Buckeye Gala’ Apple Fruit Grafted on Ten Different Rootstocks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Li, W.; Xu, X.; Qiu, C.; Wu, T.; Wei, Q.; Ma, F.; Han, Z. Progress of Apple Rootstock Breeding and Its Use. Hortic. Plant J. 2019, 5, 183–191. [Google Scholar] [CrossRef]
- Russo, N.L.; Robinson, T.L.; Fazio, G.; Aldwinckle, H.S. Field Evaluation of 64 Apple Rootstocks for Orchard Performance and Fire Blight Resistance. HortScience 2007, 42, 1517–1525. [Google Scholar] [CrossRef]
- Autio, W.; Robinson, T.; Black, B.; Blatt, S.; Cochran, D.; Cowgill, W.; Hampson, C.; Hoover, E.; Lang, G.; Miller, D. Budagovsky, Geneva, Pillnitz, and Malling Apple Rootstocks Affect ‘Honeycrisp’Performance over the First Five Years of the 2010 NC-140 ‘Honeycrisp’ Apple Rootstock Trial. J. Am. Pomol. Soc 2017, 71, 149–166. [Google Scholar]
- Lordan, J.; Fazio, G.; Francescatto, P.; Robinson, T.L., II. Horticultural Performance of ‘Honeycrisp’ Grown on a Genetically Diverse Set of Rootstocks under Western New York Climatic Conditions. Sci. Hortic. 2019, 257, 108686. [Google Scholar] [CrossRef]
- Momol, M.T.; Norelli, J.L.; Piccioni, D.E.; Momol, E.A.; Gustafson, H.L.; Cummins, J.N.; Aldwinckle, H.S. Internal Movement of Erwinia Amylovora through Symptomless Apple Scion Tissues into the Rootstock. Plant Dis. 1998, 82, 646–650. [Google Scholar] [CrossRef]
- Drake, S.R.; Larsen, F.E.; Higgins, S.S. Quality and Storage of Granny Smith’ and Greenspur’ Apples on Seedling, M. 26, and MM. 111 Rootstock. J. Am. Soc. Hortic. Sci. 1991, 116, 261–264. [Google Scholar] [CrossRef]
- Valverdi, N.A.; Kalcsits, L. Rootstock Affects Scion Nutrition and Fruit Quality during Establishment and Early Production of ‘Honeycrisp’ Apple. HortScience 2021, 56, 261–269. [Google Scholar] [CrossRef]
- Fallahi, E.; Colt, W.M.; Fallahi, B.; Chun, I.-J. The Importance of Apple Rootstocks on Tree Growth, Yield, Fruit Quality, Leaf Nutrition, and Photosynthesis with an Emphasis On Fuji. Horttechnology 2002, 12, 38–44. [Google Scholar] [CrossRef]
- Hayat, F.; Qiu, C.; Xu, X.; Wang, Y.; Wu, T.; Zhang, X.; Nawaz, M.A.; Han, Z. Rootstocks Influence Morphological and Biochemical Changes in Young ‘Red Fuji’ Apple Plants. Int. J. Agric. Biol 2019, 21, 1097–1105. [Google Scholar]
- Rufato, L.; Da Silva, P.S.; Kretzschmar, A.A.; Bogo, A.; De Macedo, T.A.; Welter, J.F.; Fazio, G.; Petry, D. Geneva® Series Rootstocks for Apple Trees under Extreme Replanting Conditions in Southern Brazil. Front. Plant Sci. 2021, 12, 712162. [Google Scholar] [CrossRef]
- Reig, G.; Lordan, J.; Sazo, M.M.; Hoying, S.; Fargione, M.; Reginato, G.; Donahue, D.J.; Francescatto, P.; Fazio, G.; Robinson, T. Long-Term Performance of ‘Gala’, Fuji’ and ‘Honeycrisp’ Apple Trees Grafted on Geneva® Rootstocks and Trained to Four Production Systems under New York State Climatic Conditions. Sci. Hortic. 2019, 244, 277–293. [Google Scholar] [CrossRef]
- Rom, R.C.; Rom, C.R.; Quamme, H.A.; Micke, W.C.; Yu, K.C.; Gaus, A.; Myers, S.C.; Meador, D.B.; Kushad, M.; Hayden, R. Performance of the NC-140 Cooperative Apple Rootstock Planting: I. Survival, Tree Size, Yield and Fruit Size. Fruit Var. J. 1996, 50, 6–11. [Google Scholar]
- Larsen, F.E.; Fritts, R. Sixteen-Year Summary of Apple Rootstock Influence on Yield, Yield Efficiency, and Trunk Growth1. J. Am. Soc. Hortic. Sci. 1982, 107, 23–27. [Google Scholar] [CrossRef]
- Hargarten, H.; Mattheis, J.; Honaas, L. Monitoring Effects of Rootstock Genotype and Soil Treatment Strategy on Postharvest Fruit Quality in ‘Gala’ Apple. HortScience 2022, 57, 789–798. [Google Scholar] [CrossRef]
- Barden, J.A.; Marini, M.E. Maturity and Quality of Delicious’ Apples as Influenced by Rootstock. J. Am. Soc. Hortic. Sci. 1992, 117, 547–550. [Google Scholar] [CrossRef]
- Klee, H.J.; Giovannoni, J.J. Genetics and Control of Tomato Fruit Ripening and Quality Attributes. Annu. Rev. Genet. 2011, 45, 41–59. [Google Scholar] [CrossRef]
- Farcuh, M.; Tajima, H.; Lerno, L.A.; Blumwald, E. Changes in Ethylene and Sugar Metabolism Regulate Flavonoid Composition in Climacteric and Non-Climacteric Plums during Postharvest Storage. Food Chem. Mol. Sci. 2022, 4, 100075. [Google Scholar] [CrossRef]
- Osorio, S.; Fernie, A.R. Biochemistry of Fruit Ripening. The Molecular Biology and Biochemistry of Fruit Ripening; Wiley-Blackwell: Chichester, UK, 2013; pp. 1–19. [Google Scholar] [CrossRef]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Cantu, D.; Bradford, K.J.; Guinard, J.-X.; Van Deynze, A. Sensory, Physicochemical and Volatile Compound Analysis of Short and Long Shelf-Life Melon (Cucumis Melo L.) Genotypes at Harvest and after Postharvest Storage. Food Chem. X 2020, 8, 100107. [Google Scholar] [CrossRef]
- Farcuh, M.; Rivero, R.M.; Sadka, A.; Blumwald, E. Ethylene Regulation of Sugar Metabolism in Climacteric and Non-Climacteric Plums. Postharvest Biol. Technol. 2018, 139, 20–30. [Google Scholar] [CrossRef]
- Costa, F.; Stella, S.; Van de Weg, W.E.; Guerra, W.; Cecchinel, M.; Dallavia, J.; Koller, B.; Sansavini, S. Role of the Genes Md-ACO1 and Md-ACS1 in Ethylene Production and Shelf Life of Apple (Malus domestica Borkh). Euphytica 2005, 141, 181–190. [Google Scholar] [CrossRef]
- Bi, S.; An, J.; Wang, X.; Hao, Y.; Rui, L.; Li, T.; Han, Y.; You, C. Ethylene Response Factor MdERF3 Promotes Anthocyanin and Proanthocyanidin Accumulation in Apple. Acta Hortic. Sin. 2019, 46, 2277–2285. [Google Scholar]
- Defilippi, B.G.; Dandekar, A.M.; Kader, A.A. Relationship of Ethylene Biosynthesis to Volatile Production, Related Enzymes, and Precursor Availability in Apple Peel and Flesh Tissues. J. Agric. Food Chem. 2005, 53, 3133–3141. [Google Scholar] [CrossRef] [PubMed]
- Farcuh, M.; Hopfer, H. Aroma Volatiles as Predictors of Chilling Injury Development during Peach (Prunus persica (L) Batsch) Cold Storage and Subsequent Shelf-Life. Postharvest Biol. Technol. 2023, 195, 112137. [Google Scholar] [CrossRef]
- Drake, S.R.; Larsen, F.E.; Fellman, J.K.; Higgins, S.S. Maturity, Storage Quality, Carbohydrate, and Mineral Content of ‘Goldspur’ Apples as Influenced by Rootstock. J. Am. Soc. Hortic. Sci. 1988, 113, 949–952. [Google Scholar] [CrossRef]
- Fallahi, E.; Richardson, D.G.; Westwood, M.N. Influence of Rootstocks and Fertilizers on Ethylene in Apple Fruit during Maturation and Storage. J. Am. Soc. Hortic. Sci. 1985, 110, 149–153. [Google Scholar] [CrossRef]
- Marini, R.P.; Barden, J.A.; Cline, J.A.; Perry, R.L.; Robinson, T. Effect of Apple Rootstocks on Average Gala’ fruit Weight at Four Locations after Adjusting for Crop Load. J. Am. Soc. Hortic. Sci. 2002, 127, 749–753. [Google Scholar] [CrossRef]
- Brown, G.R.; Wolfe, D. Rootstock Affects Maturity OfStarkspur Supreme Delicious’ Apples. HortScience 1992, 27, 76. [Google Scholar] [CrossRef]
- Larsen, F.E.; Fritts, R., Jr.; Olsen, K.L. Rootstock Influence on ‘Delicious’ and ‘Golden Delicious’ Apple Fruit Quality at Harvest and after Storage. Sci. Hortic. 1985, 26, 339–349. [Google Scholar] [CrossRef]
- Barritt, B.H.; Konishi, B.S.; Drake, S.R.; Rom, C.R. Influence of Sunlight Level and Rootstock on Apple Fruit Quality. In Proceedings of the VI International Symposium on Integrated Canopy, Rootstock, Environmental Physiology in Orchard Systems 451, Stellenbosch, South Africa, 3–6 December 1996; pp. 569–578. [Google Scholar]
- Autio, W.R. Rootstock Affect Ripening and Other Qualities of Delicious’ Apples. J. Am. Soc. Hortic. Sci. 1991, 116, 378–382. [Google Scholar] [CrossRef]
- Fallahi, E.; Kiester, M.J.; Fallahi, B.; Mahdavi, S. Rootstock, Canopy Architecture, Bark Girdling, and Scoring Influence on Growth, Productivity, and Fruit Quality at Harvest in ‘Aztec Fuji’ Apple. HortScience 2018, 53, 1629–1633. [Google Scholar] [CrossRef]
- Fallahi, E.; Fallahi, B.; Shafii, B. Irrigation and Rootstock Influence on Water Use, Tree Growth, Yield, and Fruit Quality at Harvest at Different Ages of Trees in ‘Pacific Gala’ Apple. HortScience 2013, 48, 588–593. [Google Scholar] [CrossRef]
- Dallabetta, N.; Guerra, A.; Pasqualini, J.; Fazio, G. Performance of Semi-Dwarf Apple Rootstocks in Two-Dimensional Training Systems. HortScience 2021, 56, 234–241. [Google Scholar] [CrossRef]
- Musacchi, S.; Serra, S. Apple Fruit Quality: Overview on Pre-Harvest Factors. Sci. Hortic. 2018, 234, 409–430. [Google Scholar] [CrossRef]
- Corelli-Grappadelli, L.; Lakso, A.N. Fruit Development in Deciduous Tree Crops as Affected by Physiological Factors and Environmental Conditions (Keynote). In Proceedings of the XXVI International Horticultural Congress: Key Processes in the Growth and Cropping of Deciduous Fruit and Nut Trees 636, Toronto, ON, Canada, 11–17 August 2002; pp. 425–441. [Google Scholar]
- Treder, W. Relationship between Yield, Crop Density Coefficient and Average Fruit Weight of ‘Gala’ Apple. J. Fruit Ornam. Plant Res. 2008, 16, 53–63. [Google Scholar]
- Embree, C.; Wright, H.; Nichols, D. Evaluating the Accountability of Trunk Size and Canopy Volume Models for Determining Apple Tree Production Potential across Diverse Management Regimes. In Proceedings of the VII International Symposium on Modelling in Fruit Research and Orchard Management 707, Copenhague, Danemark, 20–24 June 2004; pp. 237–243. [Google Scholar]
- Lordan, J.; Fazio, G.; Francescatto, P.; Robinson, T. Effects of Apple (Malus×domestica) Rootstocks on Scion Performance and Hormone Concentration. Sci. Hortic. 2017, 225, 96–105. [Google Scholar] [CrossRef]
- Al-Hinai, Y.K.; Roper, T.R. Rootstock Effects on Growth and Quality Of ‘gala’ apples. HortScience 2004, 39, 1231–1233. [Google Scholar] [CrossRef]
- Robinson, T.L.; Lakso, A.N.; Ren, Z. Modifying Apple Tree Canopies for Improved Production Efficiency. HortScience 1991, 26, 1005–1012. [Google Scholar] [CrossRef]
- Robinson, T.L. Interaction of Tree Form and Rootstock on Light Interception, Yield and Efficiency of “Empire”, ‘ Delicious’ and ‘ Jonagold’ apple Trees Trained to Different Systems. In Proceedings of the VI International Symposium on Integrated Canopy, Rootstock, Environmental Physiology in Orchard Systems 451, Wenatchee, WA, USA, 17–25 July 1996; pp. 427–436. [Google Scholar]
- Telias, A.; Bradeen, J.M.; Luby, J.J.; Hoover, E.E.; Allen, A.C. 9 Regulation of Anthocyanin Accumulation in Apple Peel. Hortic. Rev. (Am. Soc. Hortic. Sci). 2011, 38, 357. [Google Scholar]
- Zhang, J.; Serra, S.; Leisso, R.S.; Musacchi, S. Effect of Light Microclimate on the Quality of ‘d’ Anjou’ pears in Mature Open-Centre Tree Architecture. Biosyst. Eng. 2016, 141, 1–11. [Google Scholar] [CrossRef]
- Wertheim, S.J. Rootstock Guide: Apple, Pear, Cherry, European Plum. In Proefstation Voor de Fruitteelt (Fruit Research Station); CAB Direct: Glasgow, UK, 1998; ISBN 9080346225. [Google Scholar]
- Webster, A.D. Rootstock and Interstock Effects on Deciduous Fruit Tree Vigour, Precocity, and Yield Productivity. N. Zeal. J. Crop Hortic. Sci. 1995, 23, 373–382. [Google Scholar] [CrossRef]
- Iglesias, I.; Echeverria, G.; Soria, Y. Differences in Fruit Colour Development, Anthocyanin Content, Fruit Quality and Consumer Acceptability of Eight ‘Gala’ Apple Strains. Sci. Hortic. 2008, 119, 32–40. [Google Scholar] [CrossRef]
- Anthony, B.; Serra, S.; Musacchi, S. Optimizing Crop Load for New Apple Cultivar: “WA38”. Agronomy 2019, 9, 107. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Farcuh, M.; Cohen, Y.; Crisosto, C.; Sadka, A.; Blumwald, E. Non-Climacteric Ripening and Sorbitol Homeostasis in Plum Fruits. Plant Sci. 2015, 231, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Infante, R.; Farcuh, M.; Meneses, C. Monitoring the Sensorial Quality and Aroma through an Electronic Nose in Peaches during Cold Storage. J. Sci. Food Agric. 2008, 88, 2073–2078. [Google Scholar] [CrossRef]
- Ziosi, V.; Noferini, M.; Fiori, G.; Tadiello, A.; Trainotti, L.; Casadoro, G.; Costa, G. A New Index Based on Vis Spectroscopy to Characterize the Progression of Ripening in Peach Fruit. Postharvest Biol. Technol. 2008, 49, 319–329. [Google Scholar] [CrossRef]
- Blanpied, G.D.; Silsby, J. Predicting Harvest Date Windows for Apples; Cornell Cooperative Extension: Long Island, NY, USA, 1992; pp. 7–9. [Google Scholar]
- Johnson, R.A.; Wichern, D.W. Applied Multivariate Statistical Analysis; Qatar University: Doha, Qatar, 2007. [Google Scholar]
- Tomala, K.; Andziak, J.; Jeziorek, K.; Dziuban, R. Influence of Rootstock on the Quality of ‘Jonagold’ Apples at Harvest and after Storage. J. fruit Ornam. Plant Res. 2008, 16, 31–38. [Google Scholar]
- Mahdavi, S.; Fallahi, E.; Fazio, G. The Influence of Rootstock on Fruit Ethylene, Respiration, Index of Absorbance Difference, Fruit Quality, and Production of ‘Aztec Fuji’ Apple under a Full-Crop Condition. HortScience 2022, 57, 1–9. [Google Scholar] [CrossRef]
- Autio, W.; Robinson, T.; Archbold, D.; Cowgill, W.; Hampson, C.; Quezada, R.P.; Wolfe, D. ‘Gala’ Apple Trees on Supporter 4, P. 14, and Different Strains of B. 9, M. 9 and M. 26 Rootstocks: Final 10-Year Report on the 2002 NC-140 Apple Rootstock Trial. J. Am. Pomol. Soc. 2013, 67, 62–71. [Google Scholar]
- Autio, W.R.; Robinson, T.L.; Black, B.; Bradshaw, T.; Cline, J.A.; Crassweller, R.M.; Embree, C.G.; Hoover, E.E.; Hoying, S.A.; Iungerman, K.A. Performance of’ Fuji’ and’ McIntosh’ apple Trees after 10 Years as Affected by Several Dwarf Rootstocks in the 1999 NC-140 Apple Rootstock Trial. J. Am. Pomol. Soc. 2011, 65, 2. [Google Scholar]
- Norelli, J.L.; Holleran, H.T.; Johnson, W.C.; Robinson, T.L.; Aldwinckle, H.S. Resistance of Geneva and Other Apple Rootstocks to Erwinia Amylovora. Plant Dis. 2003, 87, 26–32. [Google Scholar] [CrossRef]
- Chávez-González, C.H.; Parra-Quezada, R.A.; Ramírez-Legarreta, M.R.; Jacobo-Cuellar, J.L. Early Performance of’ Buckeye Gala’ grafted on 13 Apple Rootstocks in Mexico. In Proceedings of the IX International Symposium on Integrating Canopy, Rootstock and Environmental Physiology in Orchard Systems 903, Geneva, NY, USA, 2–9 August 2008; pp. 301–304. [Google Scholar]
- Kuldoshin, I.A. Perspective Clonal Rootstocks for Intensive Orchards in the Central Areas of Russia. In Proceedings of the International Scientific Conference Seminar, Warsaw, Poland, 28–31 May 1999; Volume 63, p. 64. [Google Scholar]
- Elfving, D.C.; Schechter, I. Fruit Count, Fruit Weight, and Yield Relationships InDelicious’ Apple Trees on Nine Rootstock. HortScience 1993, 28, 793–795. [Google Scholar] [CrossRef]
- Forshey, C.G.; Elfving, D.C. Fruit Numbers, Fruit Size, and Yield Relationships in ‘McIntosh’ Apples1. J. Am. Soc. Hortic. Sci. 1977, 102, 399–402. [Google Scholar] [CrossRef]
- Palmer, J.W.; Giuliani, R.; Adams, H.M. Effect of Crop Load on Fruiting and Leaf Photosynthesis of ‘Braeburn’ /M. 26 Apple Trees. Tree Physiol. 1997, 17, 741–746. [Google Scholar] [CrossRef]
- Autio, W.R.; Hayden, R.A.; Micke, W.C.; Brown, G.R. Rootstock Affects Ripening, Color, and Shape of ‘ Starkspur Supreme Delicious’ Apples in the 1984 NC-140 Cooperative Planting. Fruit Var. J. 1996, 50, 45–53. [Google Scholar]
- Parvaneh, T.; Abedi, B.; Davarynejad, G.H.; Moghadam, E.G. Enzyme Activity, Phenolic and Flavonoid Compounds in Leaves of Iranian Red Flesh Apple Cultivars Grown on Different Rootstocks. Sci. Hortic. 2019, 246, 862–870. [Google Scholar] [CrossRef]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Jaunet, T.; Chi-Ham, C.; Cantu, D.; Bradford, K.J.; Van Deynze, A. Texture Diversity in Melon (Cucumis melo L.): Sensory and Physical Assessments. Postharvest Biol. Technol. 2020, 159, 111024. [Google Scholar] [CrossRef]
- Harker, F.R.; Redgwell, R.J.; Hallett, I.C.; Murray, S.H.; Carter, G. Texture of Fresh Fruit. Hortic. Rev. (Am. Soc. Hortic. Sci.) 1997, 20, 121–224. [Google Scholar]
- Autio, W.R.; Schupp, J.R.; Embree, C.G.; Moran, R.E. Early Performance of’ Cortland,’ Macoun,’ ‘ McIntosh,’ and’ Pioneer Mac’ apple Trees on Various Rootstocks in Maine, Massachusetts, and Nova Scotia. J. Am. Pomol. Soc. 2003, 57, 7. [Google Scholar]
- Farcuh, M.; Li, B.; Rivero, R.M.; Shlizerman, L.; Sadka, A.; Blumwald, E. Sugar Metabolism Reprogramming in a Non-Climacteric Bud Mutant of a Climacteric Plum Fruit during Development on the Tree. J. Exp. Bot. 2017, 68, 5813–5828. [Google Scholar] [CrossRef]
- Hu, Y.; Han, Z.; Sun, Y.; Wang, S.; Wang, T.; Wang, Y.; Xu, K.; Zhang, X.; Xu, X.; Han, Z. ERF4 Affects Fruit Firmness through TPL4 by Reducing Ethylene Production. Plant J. 2020, 103, 937–950. [Google Scholar] [CrossRef]
- Gonçalves, R.G.; Couto, J.; Almeida, D.P.F. On-Tree Maturity Control of Peach Cultivars: Comparison between Destructive and Nondestructive Harvest Indices. Sci. Hortic. 2016, 209, 293–299. [Google Scholar] [CrossRef]
- Zhang, B.; Peng, B.; Zhang, C.; Song, Z.; Ma, R. Determination of Fruit Maturity and Its Prediction Model Based on the Pericarp Index of Absorbance Difference (I AD) for Peaches. PLoS ONE 2017, 12, e0177511. [Google Scholar]
- Harker, F.R.; Maindonald, J.; Murray, S.H.; Gunson, F.A.; Hallett, I.C.; Walker, S.B. Sensory Interpretation of Instrumental Measurements 1: Texture of Apple Fruit. Postharvest Biol. Technol. 2002, 24, 225–239. [Google Scholar] [CrossRef]
- Johnston, J.W.; Hewett, E.W.; Hertog, M.L. Postharvest Softening of Apple (Malus domestica) Fruit: A Review. N. Z. J. Crop Hortic. Sci. 2002, 30, 145–160. [Google Scholar] [CrossRef]
- Li, M.; Li, P.; Ma, F.; Dandekar, A.M.; Cheng, L. Sugar Metabolism and Accumulation in the Fruit of Transgenic Apple Trees with Decreased Sorbitol Synthesis. Hortic. Res. 2018, 5, 60. [Google Scholar] [CrossRef]
- Yamaki, S. Physiology and Metabolism of Fruit Development-Biochemistry of Sugar Metabolism and Compartmentation in Fruits. Postharvest Physiol. Fruits 398 1994, 21, 109–120. [Google Scholar] [CrossRef]
- Watari, J.; Kobae, Y.; Yamaki, S.; Yamada, K.; Toyofuku, K.; Tabuchi, T.; Shiratake, K. Identification of Sorbitol Transporters Expressed in the Phloem of Apple Source Leaves. Plant Cell Physiol. 2004, 45, 1032–1041. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
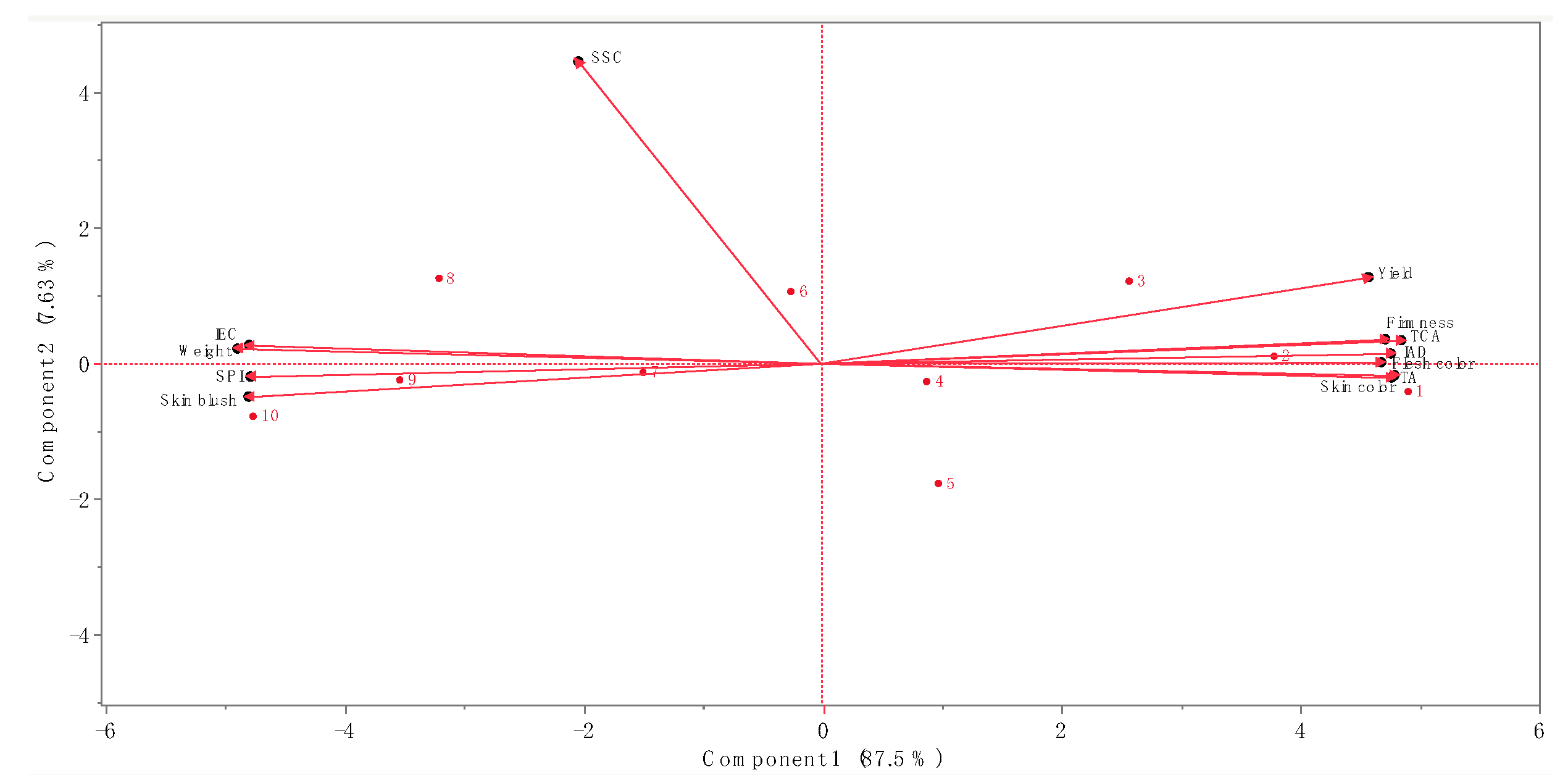{kind=link}
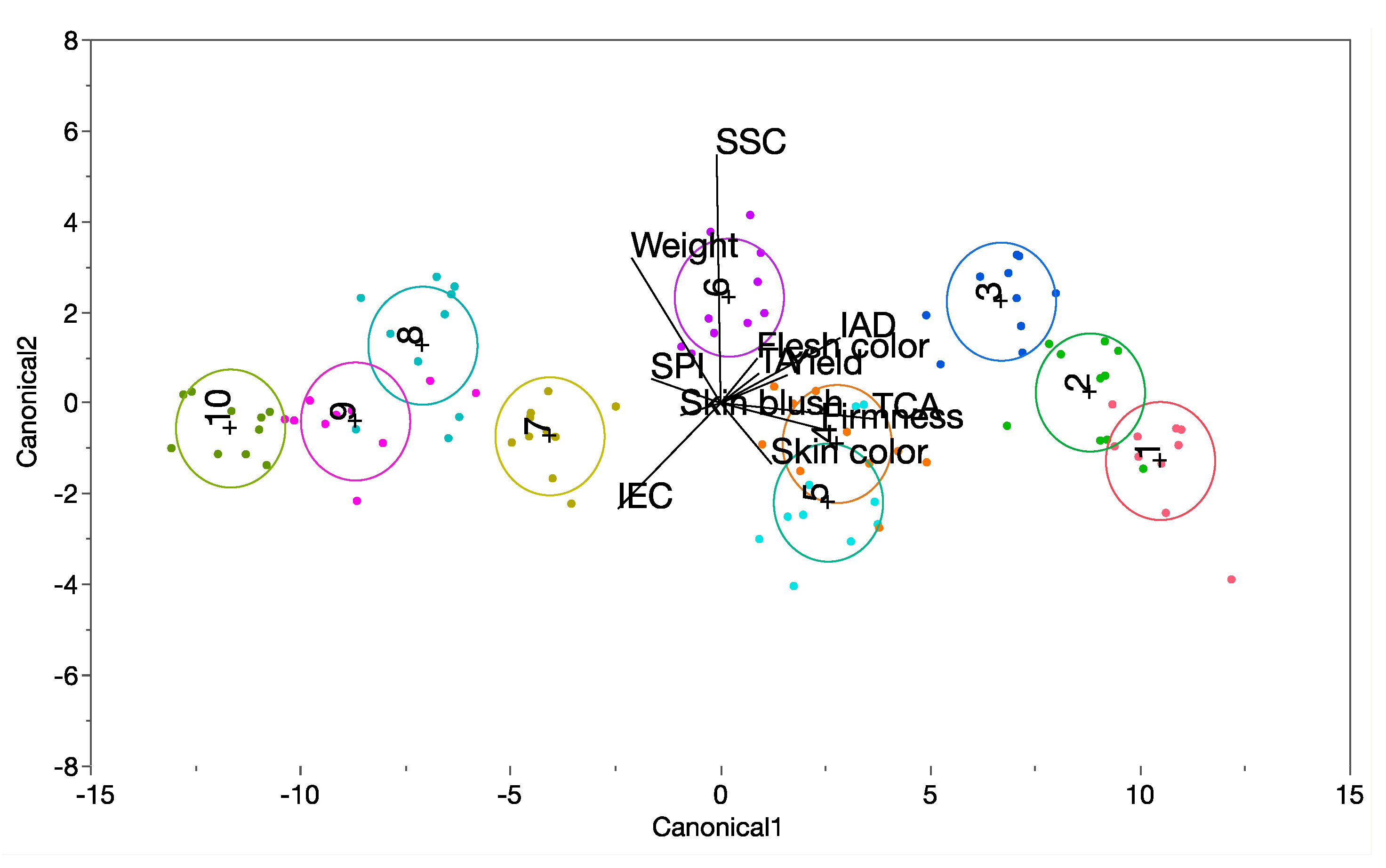{kind=link}
| Feature | TCA | Yield | Weight | IEC | SkinHue | FleshHue | IAD | Skinblush | Firmness | SPI | SSC | TA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TCA | 1.00 | 0.97 * | −0.98 * | −0.94 * | 0.95 * | 0.93 * | 0.95 * | −0.95 * | 0.97 * | −0.97 * | NS | 0.93 * |
| Yield | 1.00 | −0.92 * | −0.85 * | 0.86 * | 0.87 * | 0.88 * | −0.93 * | 0.93 * | −0.95 * | NS | 0.84 * | |
| Weight | 1.00 | 0.97 * | −0.95 * | −0.91 * | −0.96 * | 0.95 * | −0.94 * | 0.96 * | NS | −0.96 * | ||
| IEC | 1.00 | −0.95 * | −0.89 * | −0.98 * | 0.93 * | −0.91 * | 0.90 * | NS | −0.97 * | |||
| SkinHue | 1.00 | 0.95 * | 0.95 * | −0.93 * | 0.94 * | −0.92 * | NS | 0.97 * | ||||
| FleshHue | 1.00 | 0.89 * | −0.95 * | 0.89 * | −0.92 * | NS | 0.92 * | |||||
| IAD | 1.00 | −0.92 * | 0.94 * | −0.89 * | NS | 0.95 * | ||||||
| Skinblush | 1.00 | −0.89 * | 0.96 * | NS | −0.95 * | |||||||
| Firmness | 1.00 | −0.92 * | NS | 0.89 * | ||||||||
| SPI | 1.00 | NS | −0.92 * | |||||||||
| SSC | 1.00 | NS | ||||||||||
| TA | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miah, M.S.; Hinson, C.; Farcuh, M. Assessing Fruit Maturity and Quality of ‘Buckeye Gala’ Grown on a Diverse Panel of Apple (Malus domestica Borkh.) Rootstocks in Western Maryland. Agronomy 2023, 13, 2528. https://doi.org/10.3390/agronomy13102528
Miah MS, Hinson C, Farcuh M. Assessing Fruit Maturity and Quality of ‘Buckeye Gala’ Grown on a Diverse Panel of Apple (Malus domestica Borkh.) Rootstocks in Western Maryland. Agronomy. 2023; 13(10):2528. https://doi.org/10.3390/agronomy13102528
Chicago/Turabian StyleMiah, Md Shipon, Chloe Hinson, and Macarena Farcuh. 2023. "Assessing Fruit Maturity and Quality of ‘Buckeye Gala’ Grown on a Diverse Panel of Apple (Malus domestica Borkh.) Rootstocks in Western Maryland" Agronomy 13, no. 10: 2528. https://doi.org/10.3390/agronomy13102528