
Legume Cover Crop Alleviates the Negative Impact of No-Till on Tomato Productivity in a Mediterranean Organic Cropping System
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
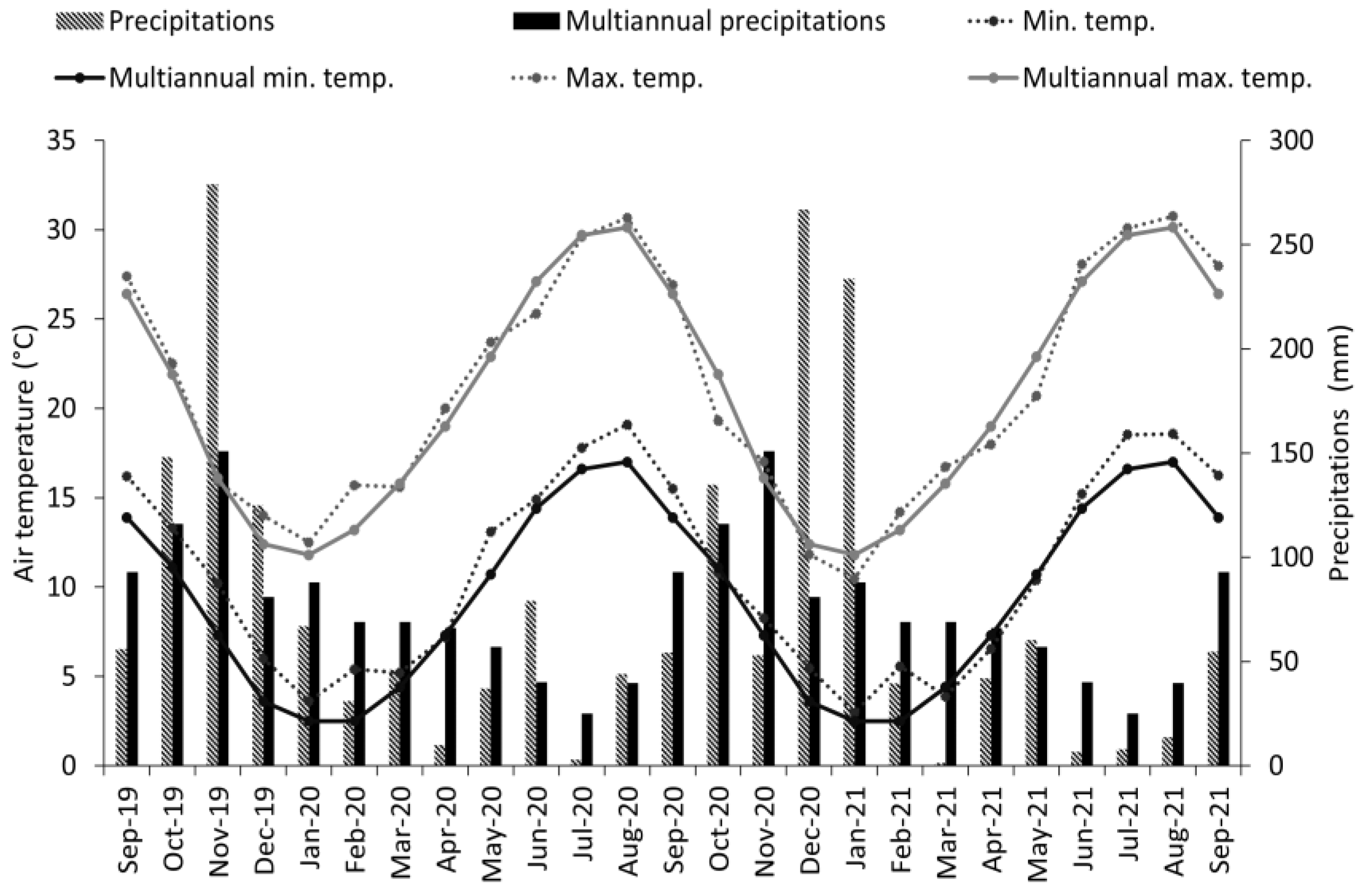
2.1. Site Description
2.2. Experimental Set-Up
2.3. Samplings and Measurements
2.4. Statistical Analysis
3. Results
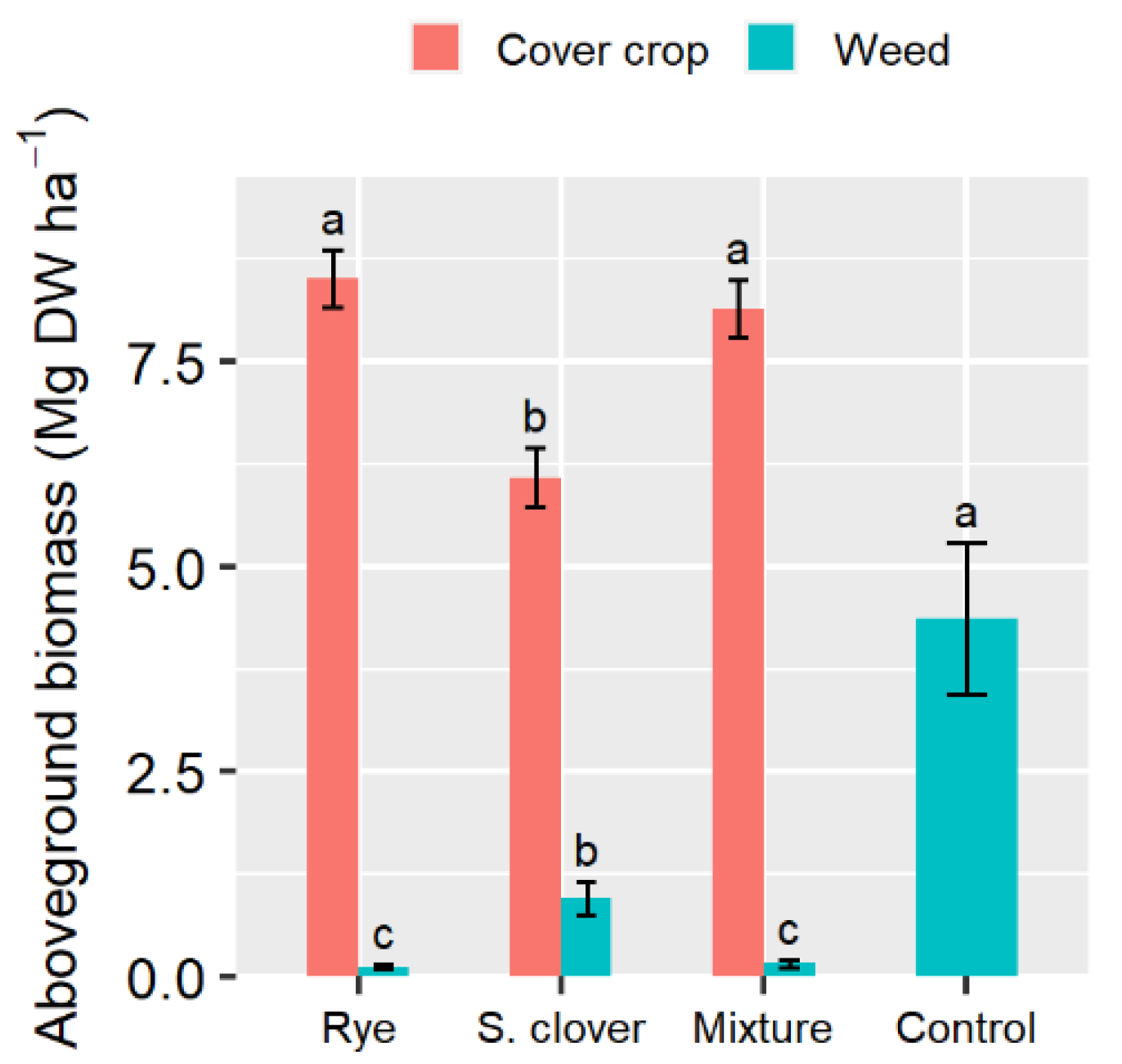
3.1. Cover Crops Performance at Termination Time: Aboveground Dry Biomass, Nutrient Accumulation, and Weed Control
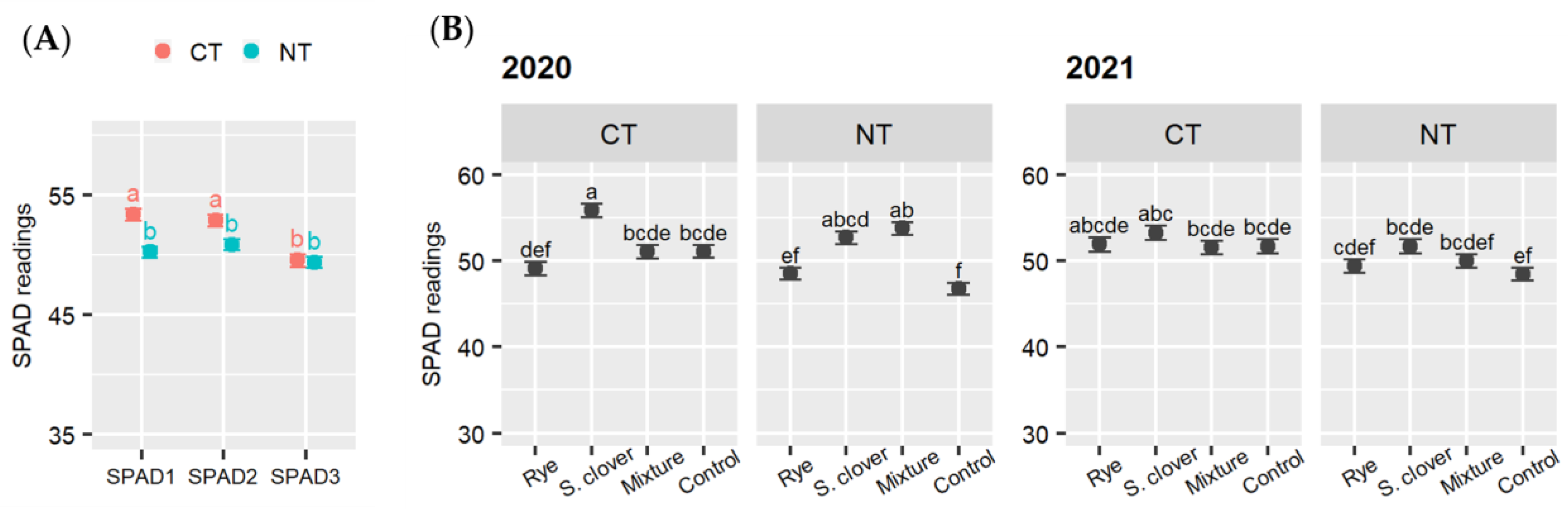
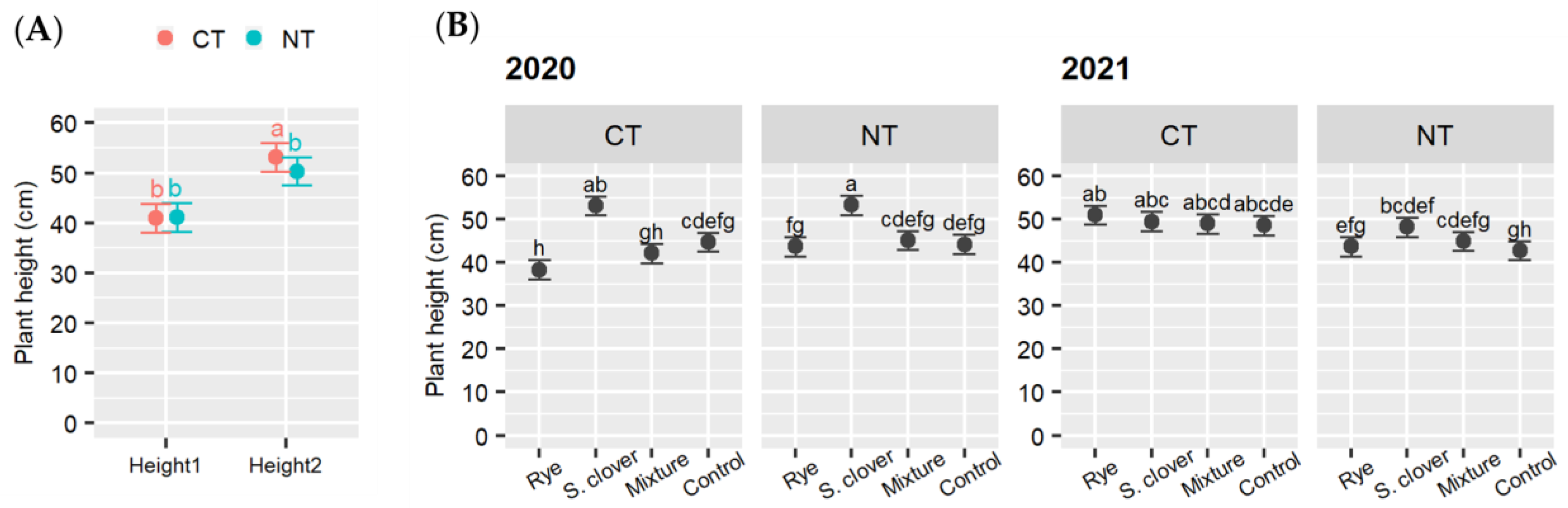
3.2. SPAD and Plant Height
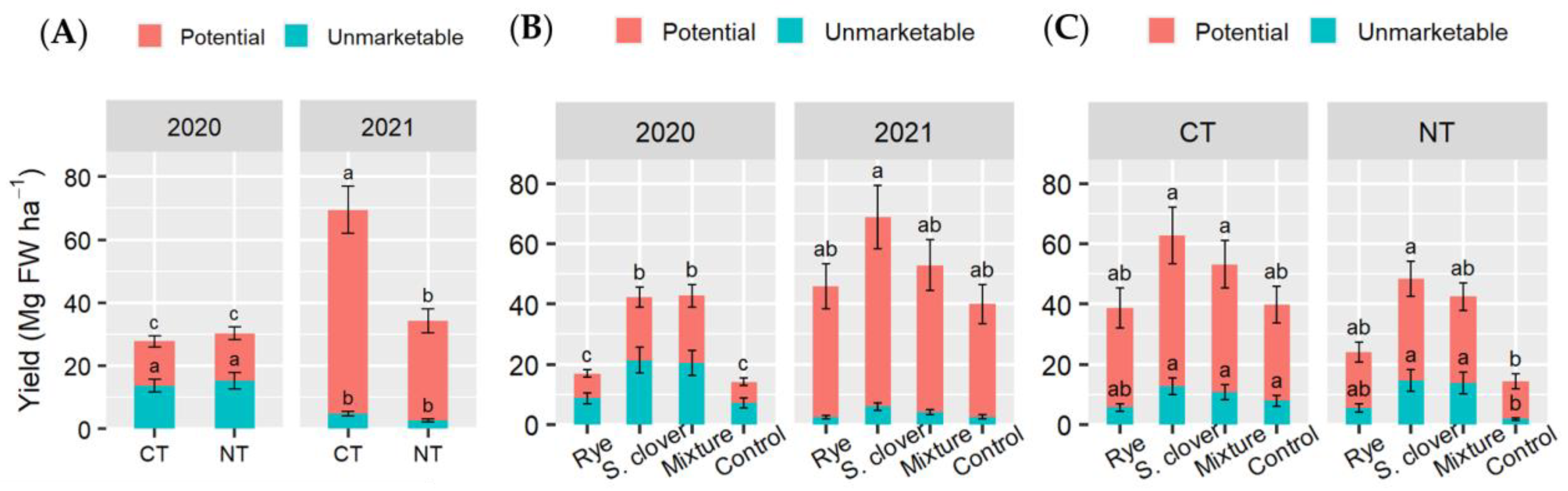
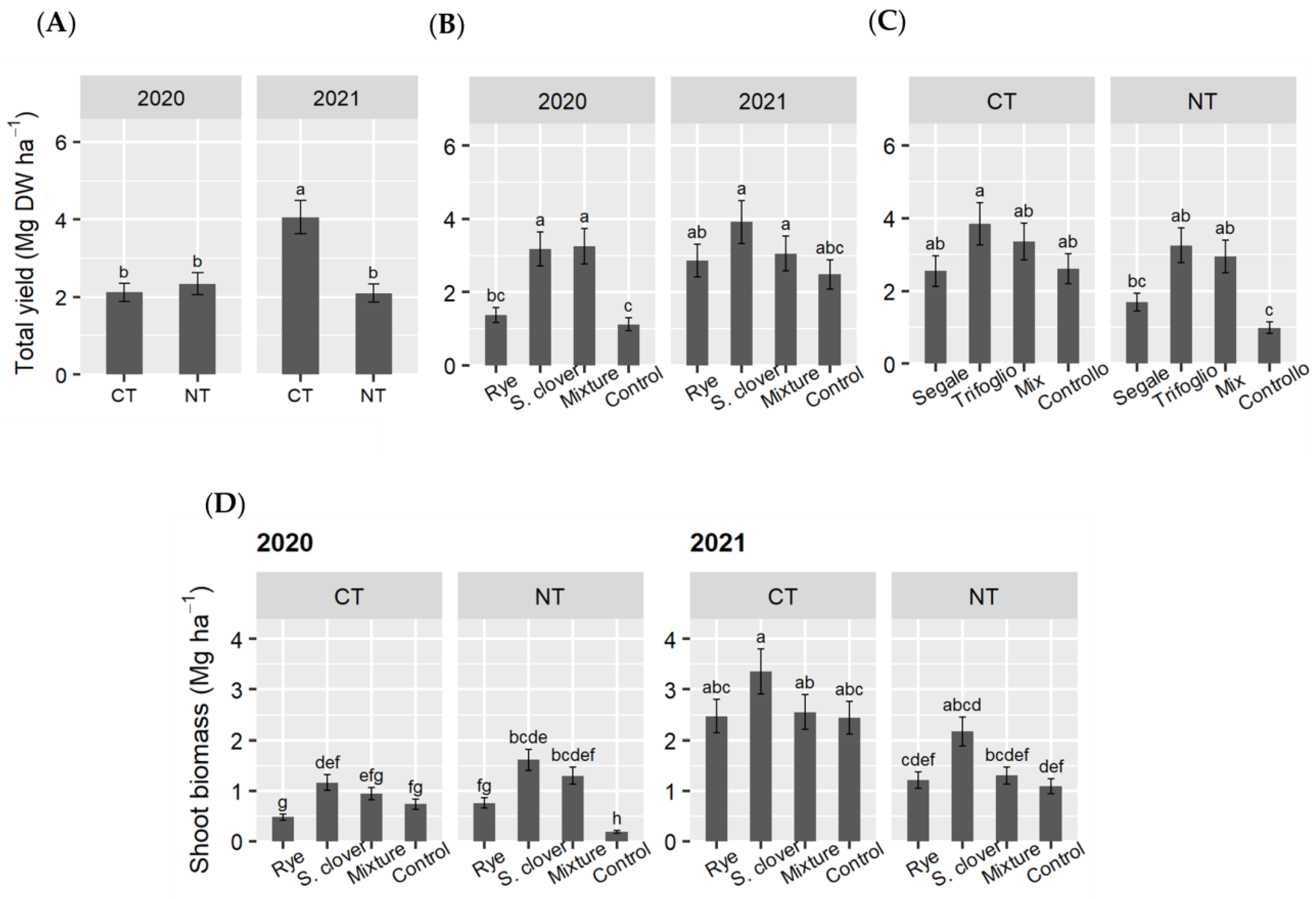
3.3. Tomato Fresh Yield and Aboveground Dry Biomass
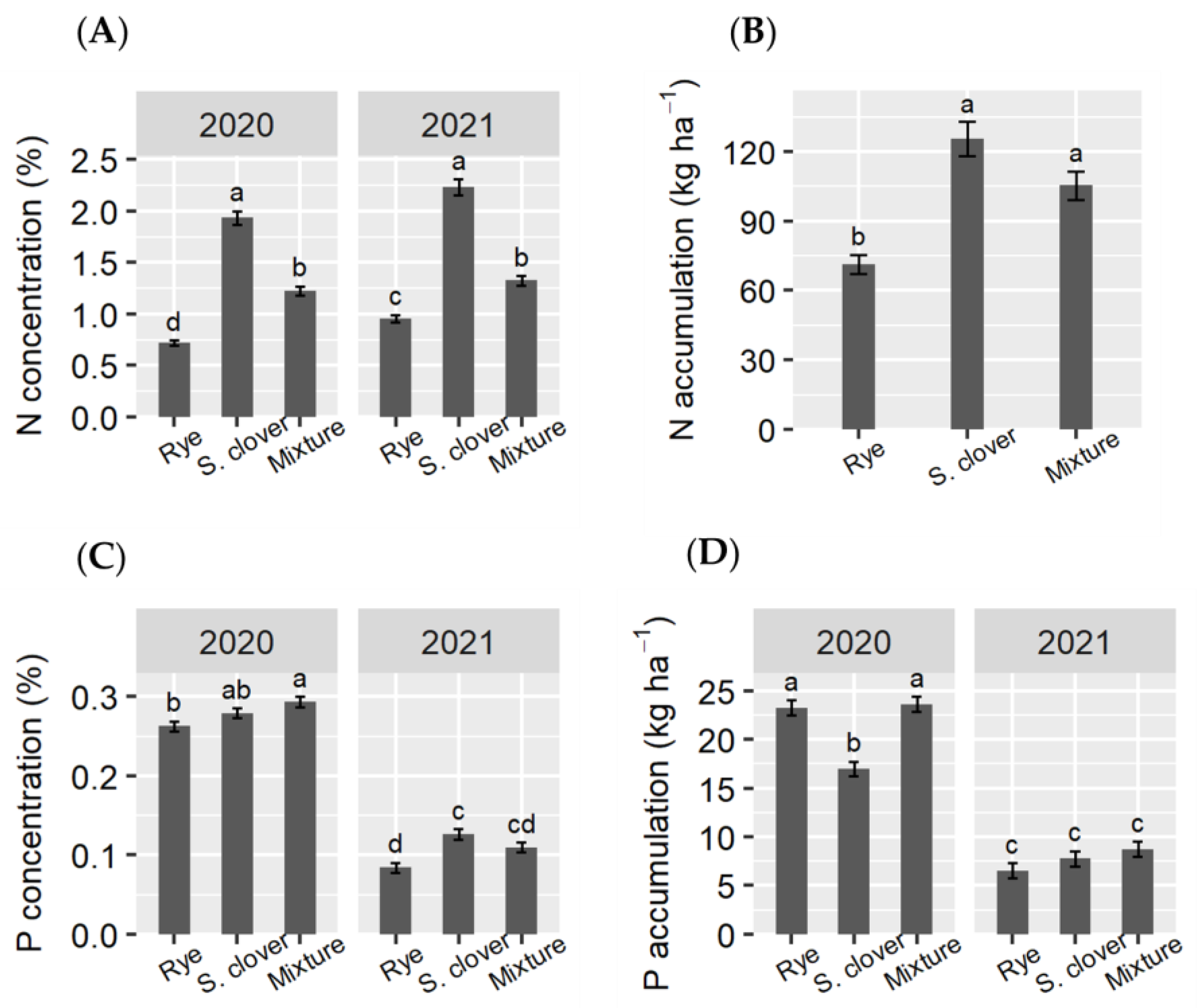
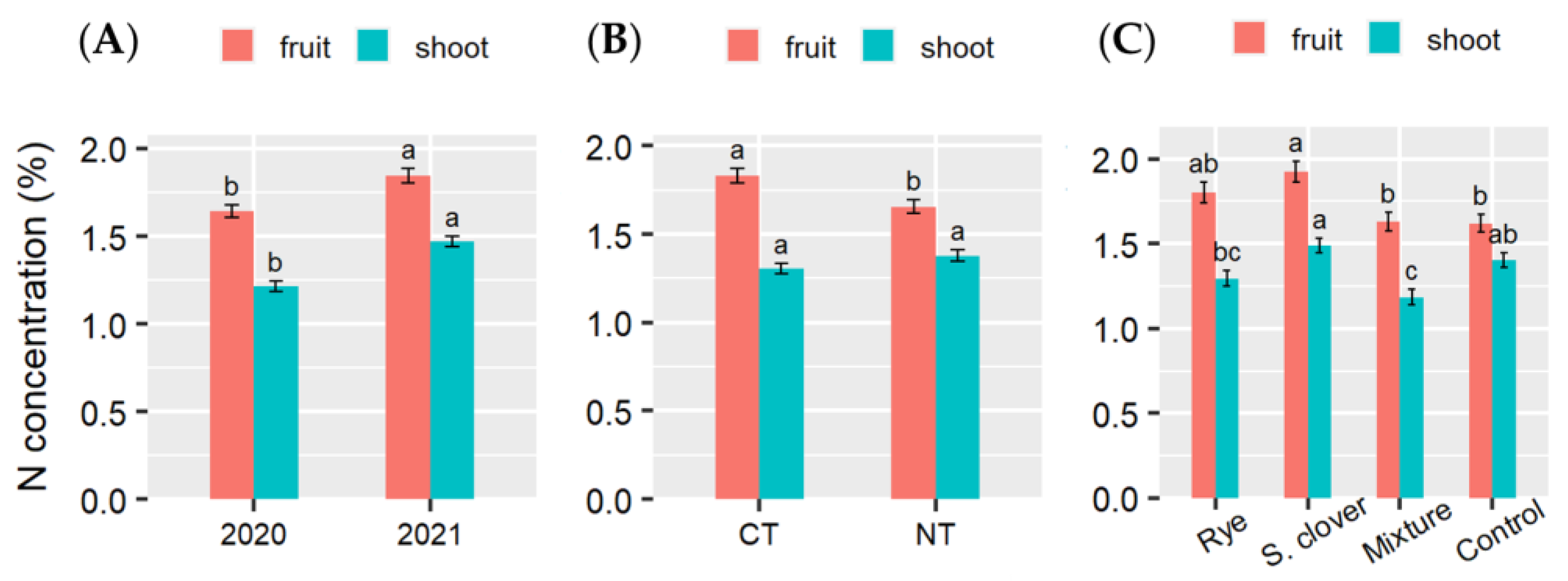
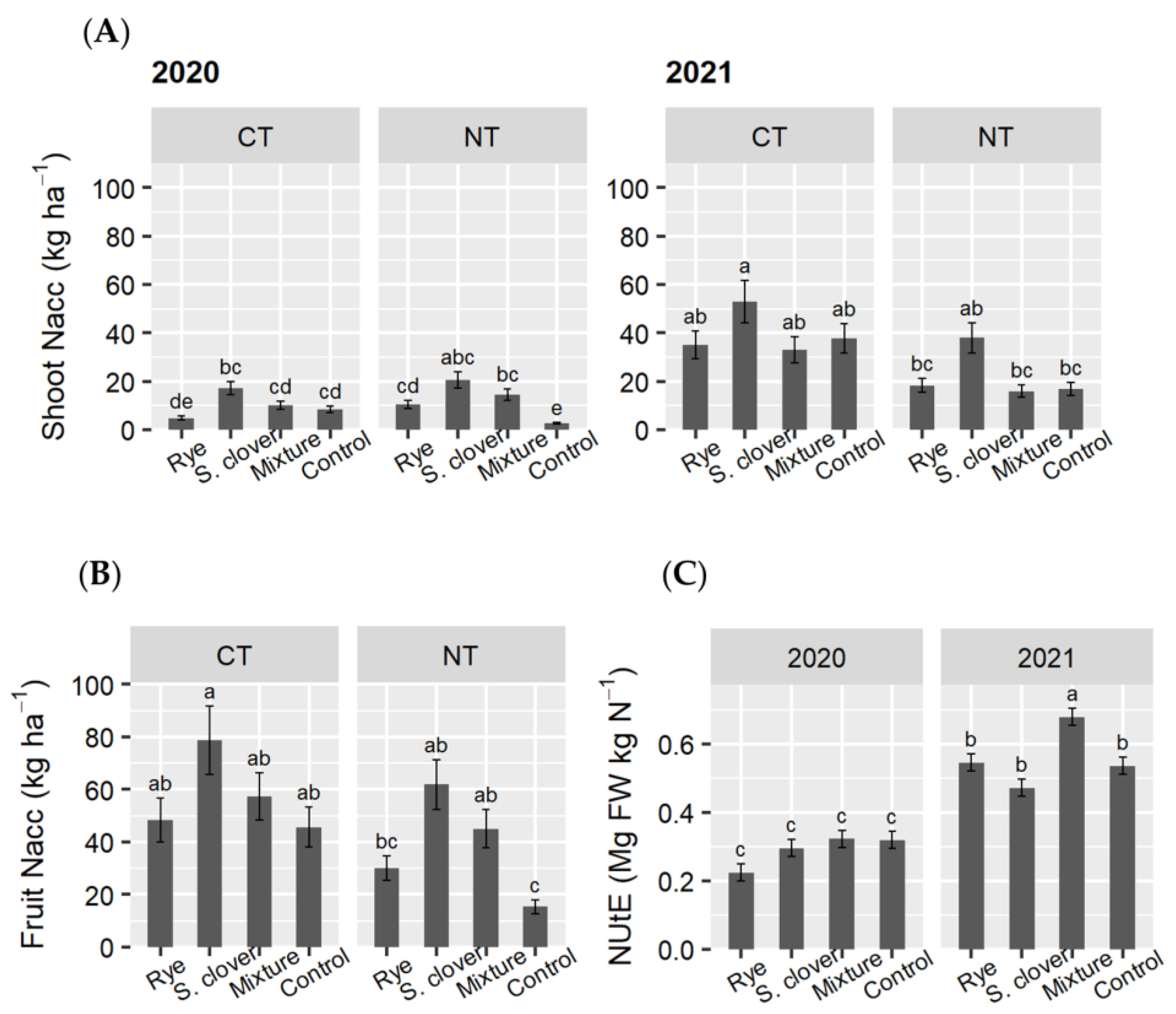
3.4. N and P Concentration and Accumulation in Tomato Shoots and Fruits
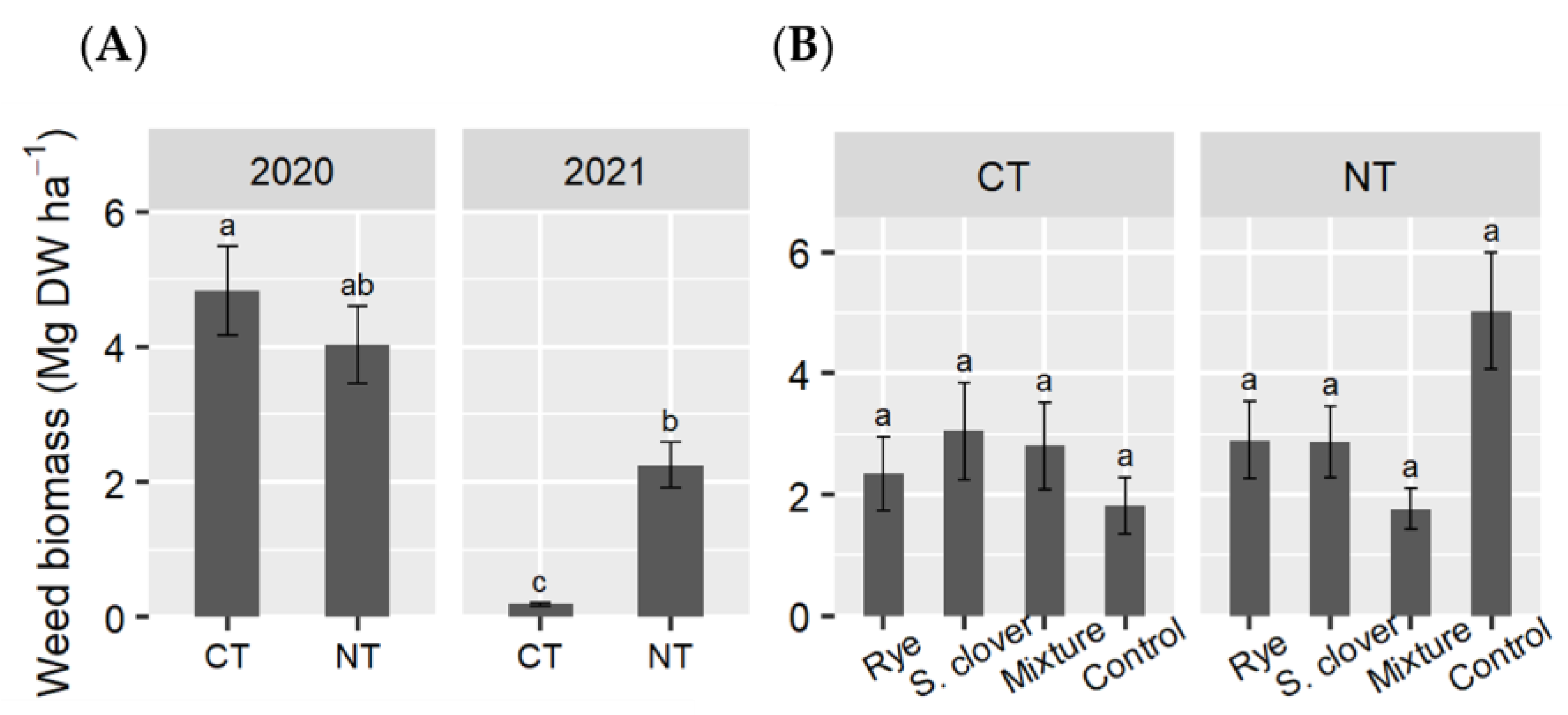
3.5. Weed Biomass at Tomato Harvest
4. Discussion
4.1. Cover Crops Performance at Termination Time: Aboveground Dry Biomass, Nutrient Accumulation, and Weed Control
4.2. Effects of Cover Crops and Tillage System on Tomato Plant Growth, Crop Productivity, Nutrient Accumulation, and Residual Weed Abundance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palm, C.; Blanco-Canqui, H.; Declerck, F.; Gatere, L.; Grace, P. Conservation Agriculture and Ecosystem Services: An Overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Vincent-Caboud, L.; Peigné, J.; Casagrande, M.; Silva, E.M. Overview of Organic Cover Crop-Based No-Tillage Technique in Europe: Farmers’ Practices and Research Challenges. Agronomy 2017, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Blanco-canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef] [Green Version]
- Shackelford, G.E.; Kelsey, R.; Dicks, L.V. Effects of Cover Crops on Multiple Ecosystem Services: Ten Meta-Analyses of Data from Arable Farmland in California and the Mediterranean. Land Use Policy 2019, 88, 104204. [Google Scholar] [CrossRef]
- Abou Chehade, L.; Antichi, D.; Martelloni, L.; Frasconi, C.; Sbrana, M.; Mazzoncini, M.; Peruzzi, A. Evaluation of the Agronomic Performance of Organic Processing Tomato as Affected by Different Cover Crop Residues Management. Agronomy 2019, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Antichi, D.; Sbrana, M.; Martelloni, L.; Abou Chehade, L.; Fontanelli, M.; Raffaelli, M.; Mazzoncini, M.; Peruzzi, A.; Frasconi, C. Agronomic Performances of Organic Field Vegetables Managed with Conservation Agriculture Techniques: A Study from Central Italy. Agronomy 2019, 9, 810. [Google Scholar] [CrossRef] [Green Version]
- Büchi, L.; Gebhard, C.; Liebisch, F.; Sinaj, S.; Ramseier, H.; Charles, R. Accumulation of Biologically Fixed Nitrogen by Legumes Cultivated as Cover Crops in Switzerland. Plant Soil 2015, 393, 163–175. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, R.M.; Smith, P. A Critical Review of the Impacts of Cover Crops on Nitrogen Leaching, Net Greenhouse Gas Balance and Crop Productivity. Glob. Chang. Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef] [Green Version]
- Thapa, R.; Poffenbarger, H.; Tully, K.L.; Ackroyd, V.J.; Kramer, M.; Mirsky, S.B. Biomass Production and Nitrogen Accumulation by Hairy Vetch–Cereal Rye Mixtures: A Meta-Analysis. Agron. J. 2018, 110, 1197–1208. [Google Scholar] [CrossRef] [Green Version]
- Mirsky, S.B.; Ryan, M.R.; Teasdale, J.R.; Curran, W.S.; Reberg-Horton, C.S.; Spargo, J.T.; Wells, M.S.; Keene, C.L.; Moyer, J.W. Overcoming Weed Management Challenges in Cover Crop—Based Organic Rotational No-Till Soybean Production in the Eastern United States. Weed Technol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Wells, M.S.; Smith, A.N.; Grossman, J.M. The Reduction of Plant-Available Nitrogen by Cover Crop Mulches and Subsequent Effects on Soybean Performance and Weed Interference. Agron. J. 2013, 105, 539–545. [Google Scholar] [CrossRef]
- Miville, D.; Leroux, G.D. Rolled Winter Rye—Hairy Vetch Cover Crops for Weed Control in No-till Pumpkin. Weed Technol. 2018, 32, 251–259. [Google Scholar] [CrossRef]
- Campanelli, G.; Testani, E.; Canali, S.; Ciaccia, C.; Trinchera, A. Effects of Cereals as Agro-Ecological Service Crops and No-till on Organic Melon, Weeds and N Dynamics. Biol. Agric. Hortic. 2019, 35, 275–287. [Google Scholar] [CrossRef]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Tilman, D.; Lehman, C.L.; Thomson, K.T. Plant Diversity and Ecosystem Productivity: Theoretical Considerations. Proc. Natl. Acad. Sci. USA 1997, 94, 1857–1861. [Google Scholar] [CrossRef]
- Wortman, S.E.; Francis, C.A.; Bernards, M.L.; Drijber, R.A.; Lindquist, J.L. Optimizing Cover Crop Benefits with Diverse Mixtures and an Alternative Termination Method. Agron. J. 2012, 104, 1425–1435. [Google Scholar] [CrossRef] [Green Version]
- Finney, D.M.; White, C.M.; Kaye, J.P. Biomass Production and Carbon/Nitrogen Ratio Influence Ecosystem Services from Cover Crop Mixtures. Agron. J. 2016, 108, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Ranaldo, M.; Carlesi, S.; Costanzo, A.; Arberi, P.B. Functional Diversity of Cover Crop Mixtures Enhances Biomass Yield and Weed Suppression in a Mediterranean Agroecosystem. Weed Res. 2019, 60, 96–108. [Google Scholar] [CrossRef]
- Elhakeem, A.; Bastiaans, L.; Houben, S.; Couwenberg, T.; Makowski, D.; Werf, W. Van Der Do Cover Crop Mixtures Give Higher and More Stable Yields than Pure Stands ? Field Crop. Res. 2021, 270, 108217. [Google Scholar] [CrossRef]
- Reiss, E.R.; Drinkwater, L.E. Promoting Enhanced Ecosystem Services from Cover Crops Using Intra- and Interspecific Diversity. Agric. Ecosyst. Environ. 2022, 323, 107586. [Google Scholar] [CrossRef]
- Lawson, A.; Cogger, C.; Bary, A.; Fortuna, A. Influence of Seeding Ratio, Planting Date, and Termination Date on Rye-Hairy Vetch Cover Crop Mixture Performance under Organic Management. PLoS ONE 2015, 10, e0129597. [Google Scholar] [CrossRef]
- Hayden, Z.D.; Brainard, D.C.; Henshaw, B.; Ngouajio, M. Winter Annual Weed Suppression in Rye—Vetch Cover Crop Mixtures. Weed Technol. 2012, 26, 818–825. [Google Scholar] [CrossRef]
- Hayden, Z.D.; Ngouajio, M.; Brainard, D.C. Rye—Vetch Mixture Proportion Tradeoffs: Cover Crop Productivity, Nitrogen Accumulation, and Weed Suppression. Agron. J. 2014, 106, 467. [Google Scholar] [CrossRef]
- White, K.E.; Brennan, E.B.; Cavigelli, M.A.; Smith, R.F. Winter Cover Crops Increased Nitrogen Availability and Efficient Use during Eight Years of Intensive Organic Vegetable Production. PLoS ONE 2022, 17, e0267757. [Google Scholar] [CrossRef] [PubMed]
- Fiorini, A.; Remelli, S.; Boselli, R.; Mantovi, P.; Ardenti, F.; Trevisan, M.; Menta, C.; Tabaglio, V. Driving Crop Yield, Soil Organic C Pools, and Soil Biodiversity with Selected Winter Cover Crops under No-Till. Soil Tillage Res. 2022, 217, 105283. [Google Scholar] [CrossRef]
- Radicetti, E.; Mancinelli, R.; Moscetti, R.; Campiglia, E. Management of Winter Cover Crop Residues under Different Tillage Conditions Affects Nitrogen Utilization Efficiency and Yield of Eggplant (Solanum melanogena L.) in Mediterranean Environment. Soil Tillage Res. 2016, 155, 329–338. [Google Scholar] [CrossRef]
- SINAB Sistema d’Informazione Nazionale sull’Agricoltura Biologica. Available online: https://www.sinab.it/superfici (accessed on 21 May 2023).
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Braunschweig, Germany, 2001. [Google Scholar]
- Frasconi, C.; Martelloni, L.; Raffaelli, M.; Fontanelli, M.; Abou Chehade, L.; Peruzzi, A.; Antichi, D. A field vegetable transplanter for use in both tilled and no-till soils. Trans ASABE 2019, 62, 593–602. [Google Scholar] [CrossRef]
- Smith, R.G.; Atwood, L.W.; Warren, N.D. Increased Productivity of a Cover Crop Mixture Is Not Associated with Enhanced Agroecosystem Services. PLoS ONE 2014, 9, e097351. [Google Scholar] [CrossRef] [Green Version]
- Mason, N.W.H.; Orwin, K.H.; Lambie, S.; Waugh, D.; Pronger, J.; Perez, C.; Paul, C. Resource-Use Efficiency Drives Overyielding via Enhanced Complementarity. Oecologia 2020, 193, 995–1010. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amossé, C.; Jeangros, B.; Walter, A.; Charles, R. Specific Interactions Leading to Transgressive Overyielding in Cover Crop Mixtures. Agric. Ecosyst. Environ. 2017, 241, 88–99. [Google Scholar] [CrossRef]
- Siebenkäs, A.; Schumacher, J.; Roscher, C. Resource Availability Alters Biodiversity Effects in Experimental Grass-Forb Mixtures Alrun. PLoS ONE 2016, 11, e0158110. [Google Scholar] [CrossRef] [Green Version]
- Blesh, J. Functional Traits in Cover Crop Mixtures: Biological Nitrogen Fixation and Multifunctionality. J. Appl. Ecol. 2018, 55, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.G.; Warren, N.D.; Cordeau, S. Are Cover Crop Mixtures Better at Suppressing Weeds than Cover Crop Monocultures? Weed Sci. 2020, 68, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Baraibar, B.; Hunter, M.C.; Schipanski, M.E.; Hamilton, A.; Mortensen, D.A. Weed Suppression in Cover Crop Monocultures and Mixtures. Weed Sci. 2018, 66, 121–133. [Google Scholar] [CrossRef]
- Mckenzie-Gopsill, A.; Mills, A.; Macdonald, A.N.; Wyand, S. The Importance of Species Selection in Cover Crop Mixture Design. Weed Sci. 2022, 70, 436–447. [Google Scholar] [CrossRef]
- Maclaren, C.; Swanepoel, P.; Bennett, J.; Wright, J.; Dehnen-schmutz, K. Cover Crop Biomass Production Is More Important than Diversity for Weed Suppression. Crop Sci. 2019, 59, 733–748. [Google Scholar] [CrossRef] [Green Version]
- Christina, M.; Négrier, A.; Marnotte, P.; Viaud, P.; Auzoux, S.; Techer, P.; Hoarau, E. A Trait-Based Analysis to Assess the Ability of Cover Crops to Control Weeds in a Tropical Island. Eur. J. Agron. 2021, 128, 126316. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy 2021, 11, 2104. [Google Scholar] [CrossRef]
- Boselli, R.; Anders, N.; Fiorini, A.; Ganimede, C.; Faccini, N.; Marocco, A.; Schulz, M.; Tabaglio, V. Improving Weed Control in Sustainable Agro-Ecosystems: Role of Cultivar and Termination Timing of Rye Cover Crop. Ital. J. Agron. 2021, 16, 1807. [Google Scholar] [CrossRef]
- Grint, K.R.; Arneson, N.J.; Oliveira, M.C.; Smith, D.H.; Werle, R. Cereal Rye Cover Crop Terminated at Crop Planting Reduces Early-Season Weed Density and Biomass in Wisconsin Corn—Soybean Production. Agrosystems Geosci. Environ. 2022, 5, e20245. [Google Scholar] [CrossRef]
- Abou Chehade, L.; Puig, C.G.; Souto, C.; Antichi, D.; Mazzoncini, M.; Pedrol, N. Rye (Secale cereale L.) and Squarrose Clover (Trifolium squarrosum L.) Cover Crops Can Increase Their Allelopathic Potential for Weed Control When Used Mixed as Dead Mulch. Ital. J. Agron. 2021, 16, 1869. [Google Scholar] [CrossRef]
- Adeux, G.; Antichi, D.; Carlesi, S.; Mazzoncini, M.; Munier-jolain, N.; Paolo, B. Cover Crops Promote Crop Productivity but Do Not Enhance Weed Management in Tillage-Based Cropping Systems. Eur. J. Agron. 2021, 123, 126221. [Google Scholar] [CrossRef]
- Campiglia, E.; Mancinelli, R.; Radicetti, E. Influence of No-Tillage and Organic Mulching on Tomato (Solanum lycopersicum L.) Production and Nitrogen Use in the Mediterranean Environment of Central Italy. Sci. Hortic. 2011, 130, 588–598. [Google Scholar] [CrossRef]
- Little, N.G.; Ditommaso, A.; Westbrook, A.S.; Ketterings, Q.M.; Mohler, C.L. Effects of Fertility Amendments on Weed Growth and Weed—Crop Competition: A Review. Weed Sci. 2021, 69, 132–146. [Google Scholar] [CrossRef]
- Testani, E.; Montemurro, F.; Ciaccia, C.; Testani, E. Agroecological Practices for Organic Lettuce: Effects on Yield, Nitrogen Status and Nitrogen Utilisation Efficiency Nitrogen Status and Nitrogen Utilisation efficiency. Biol. Agric. Hortic. 2019, 36, 84–95. [Google Scholar] [CrossRef]
- Radicetti, E.; Mancinelli, R.; Campiglia, E. Impact of Managing Cover Crop Residues on the Floristic Composition and Species Diversity of the Weed Community of Pepper Crop (Capsicum annuum L.). Crop Prot. 2013, 44, 109–119. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Nicoletti, F.; Leteo, F.; Campanelli, G. The Impact of Legume and Cereal Cover Crops on Rhizosphere Microbial Communities of Subsequent Vegetable Crops for Contrasting Crop Decline. Biol. Control 2018, 120, 17–25. [Google Scholar] [CrossRef]
- Kramberger, B.; Gselman, A.; Kristl, J.; Lesnik, M.; Šuštar, V.; Muršec, M.; Podvršnik, M. Winter Cover Crop: The Effects of Grass—Clover Mixture Proportion and Biomass Management on Maize and the Apparent Residual N in the Soil. Eur. J. Agron. 2014, 55, 63–71. [Google Scholar] [CrossRef]
- Canali, S.; Campanelli, G.; Ciaccia, C.; Leteo, F.; Testani, E.; Montemurro, F. Conservation Tillage Strategy Based on the Roller Crimper Technology for Weed Control in Mediterranean Vegetable Organic Cropping Systems. Eur. J. Agron. 2013, 50, 11–18. [Google Scholar] [CrossRef]
- Ciaccia, C.; Canali, S.; Campanelli, G.; Testani, E.; Montemurro, F.; Leteo, F.; Delate, K. Effect of Roller-Crimper Technology on Weed Management in Organic Zucchini Production in a Mediterranean Climate Zone. Renew. Agric. Food Syst. 2016, 31, 111–121. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 2019–2020 | 2020–2021 |
|---|---|---|
| pH | 6.72 | 8.27 |
| Organic matter (g 100 g−1) | 2.19 | 1.45 |
| Total N (g 100 g−1) | 0.12 | 0.09 |
| Available P (mg 100 g−1) | 0.49 | 0.91 |
| Cation exchange capacity (meq 100 g−1) | 10.81 | 3.56 |
| Electrical conductivity (µS cm−1) | 113.54 | 54.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Chehade, L.; Antichi, D.; Frasconi, C.; Sbrana, M.; Tramacere, L.G.; Mazzoncini, M.; Peruzzi, A. Legume Cover Crop Alleviates the Negative Impact of No-Till on Tomato Productivity in a Mediterranean Organic Cropping System. Agronomy 2023, 13, 2027. https://doi.org/10.3390/agronomy13082027
Abou Chehade L, Antichi D, Frasconi C, Sbrana M, Tramacere LG, Mazzoncini M, Peruzzi A. Legume Cover Crop Alleviates the Negative Impact of No-Till on Tomato Productivity in a Mediterranean Organic Cropping System. Agronomy. 2023; 13(8):2027. https://doi.org/10.3390/agronomy13082027
Chicago/Turabian StyleAbou Chehade, Lara, Daniele Antichi, Christian Frasconi, Massimo Sbrana, Lorenzo Gabriele Tramacere, Marco Mazzoncini, and Andrea Peruzzi. 2023. "Legume Cover Crop Alleviates the Negative Impact of No-Till on Tomato Productivity in a Mediterranean Organic Cropping System" Agronomy 13, no. 8: 2027. https://doi.org/10.3390/agronomy13082027