
Ethylene Regulates Combined Drought and Low Nitrogen Tolerance in Wheat: Proteomic Analysis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Proteomics Analysis and Targeted Protein Quantification by PRM
2.3. Total RNA Extraction and qRT-PCR
2.4. Physiological Assays
3. Results
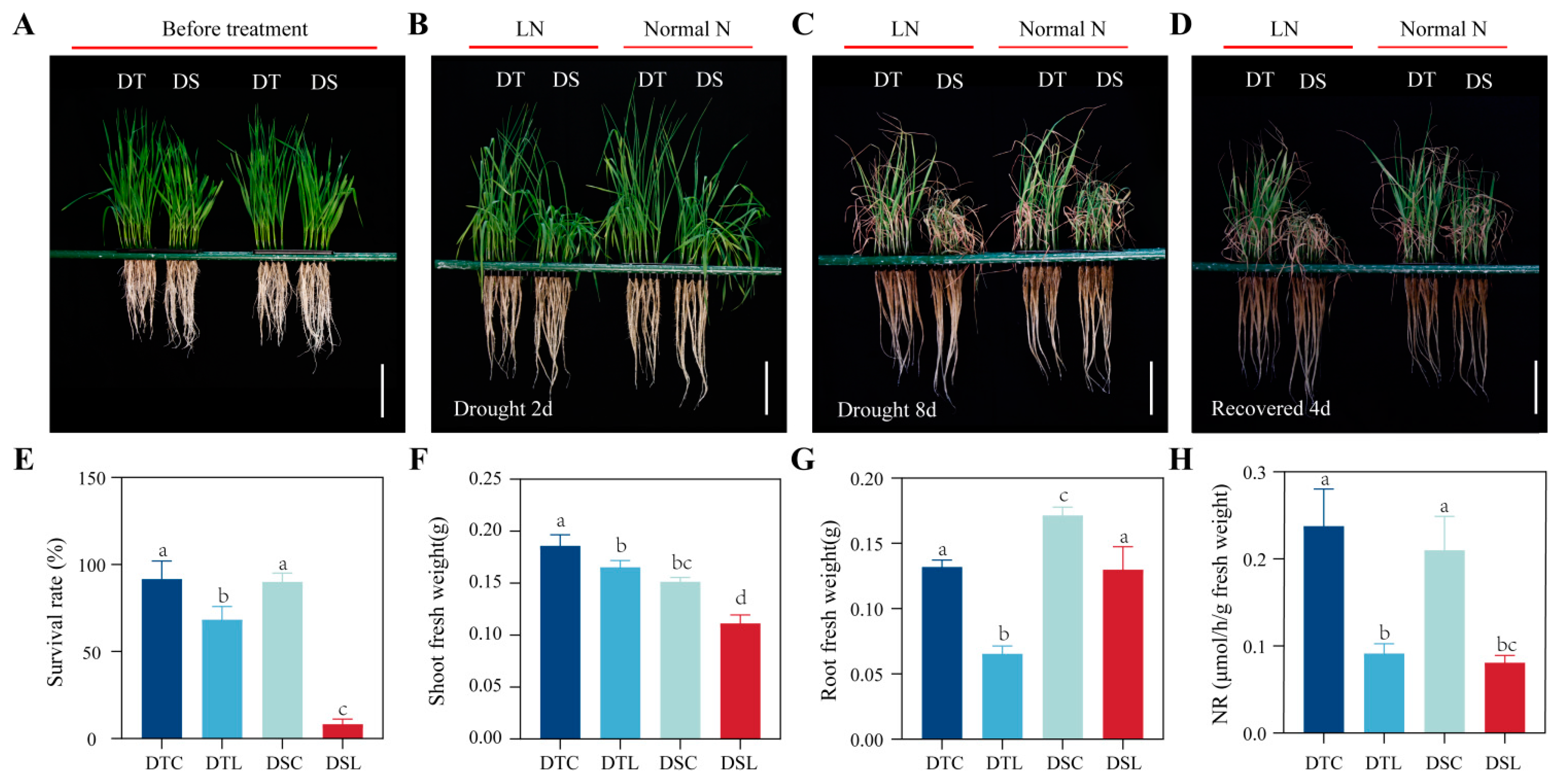
3.1. Various Nitrogen Concentration Treatments Significantly Affected Drought Tolerance
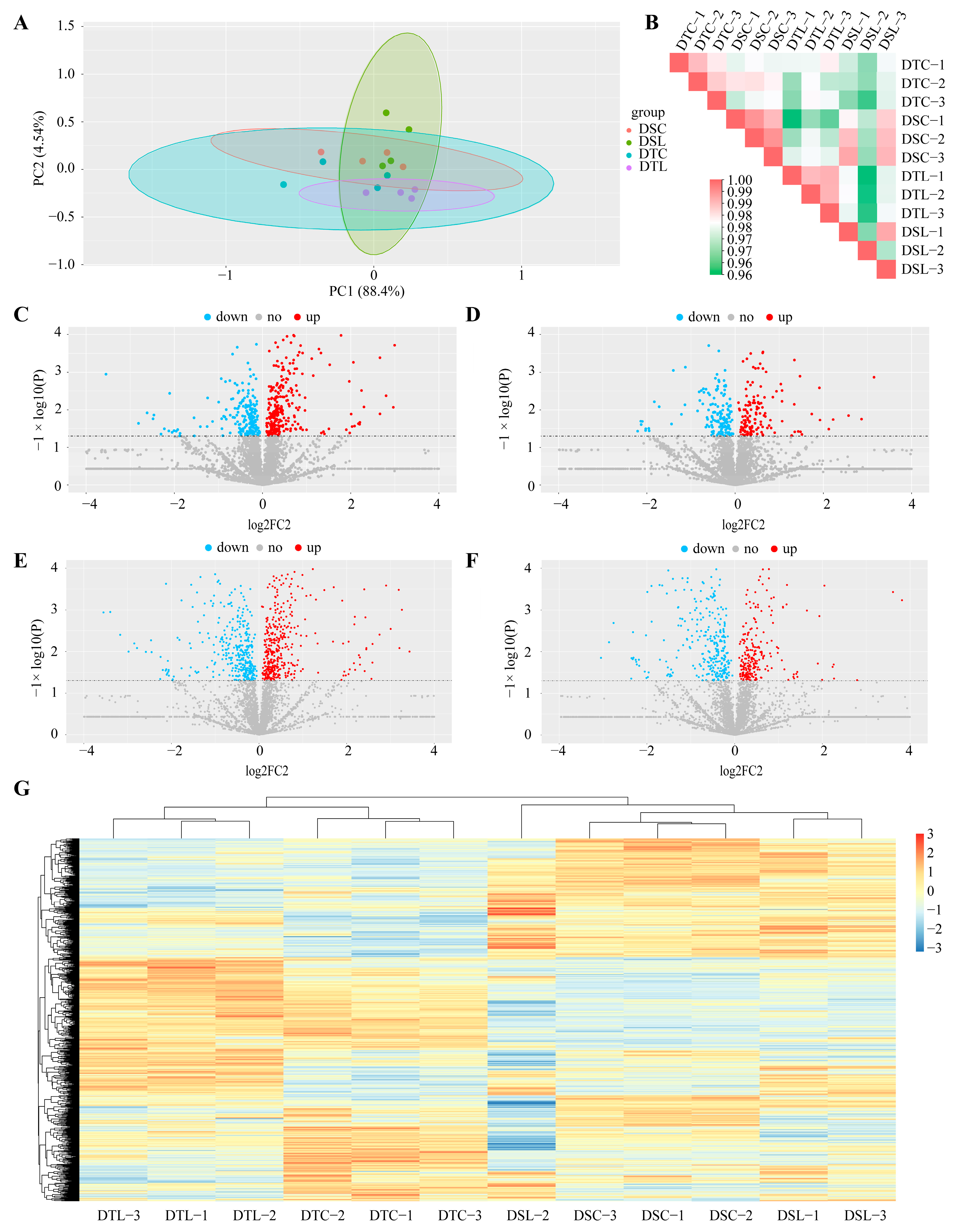
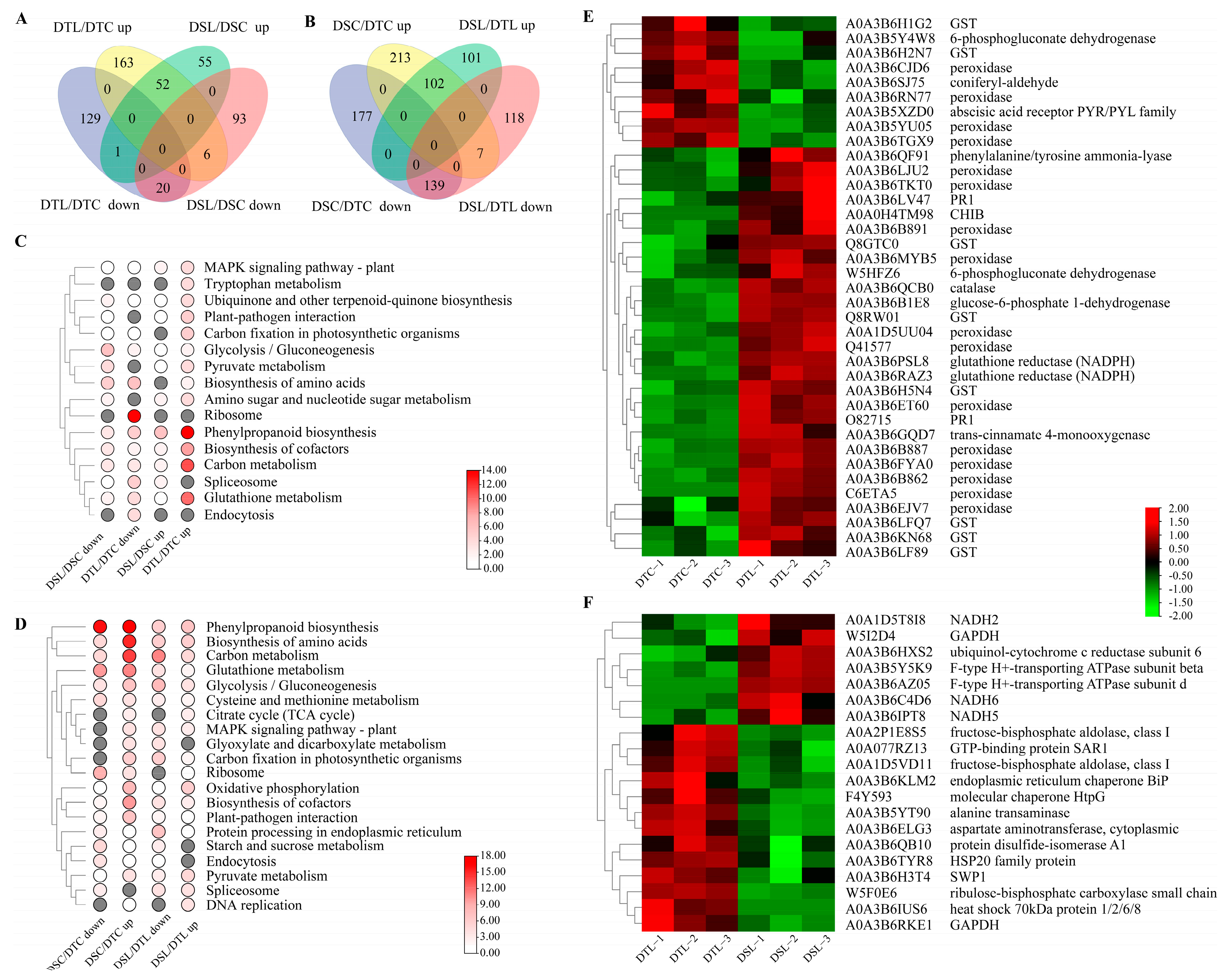
3.2. Responses to Combined Drought and LN Stress and Drought Stress Alone
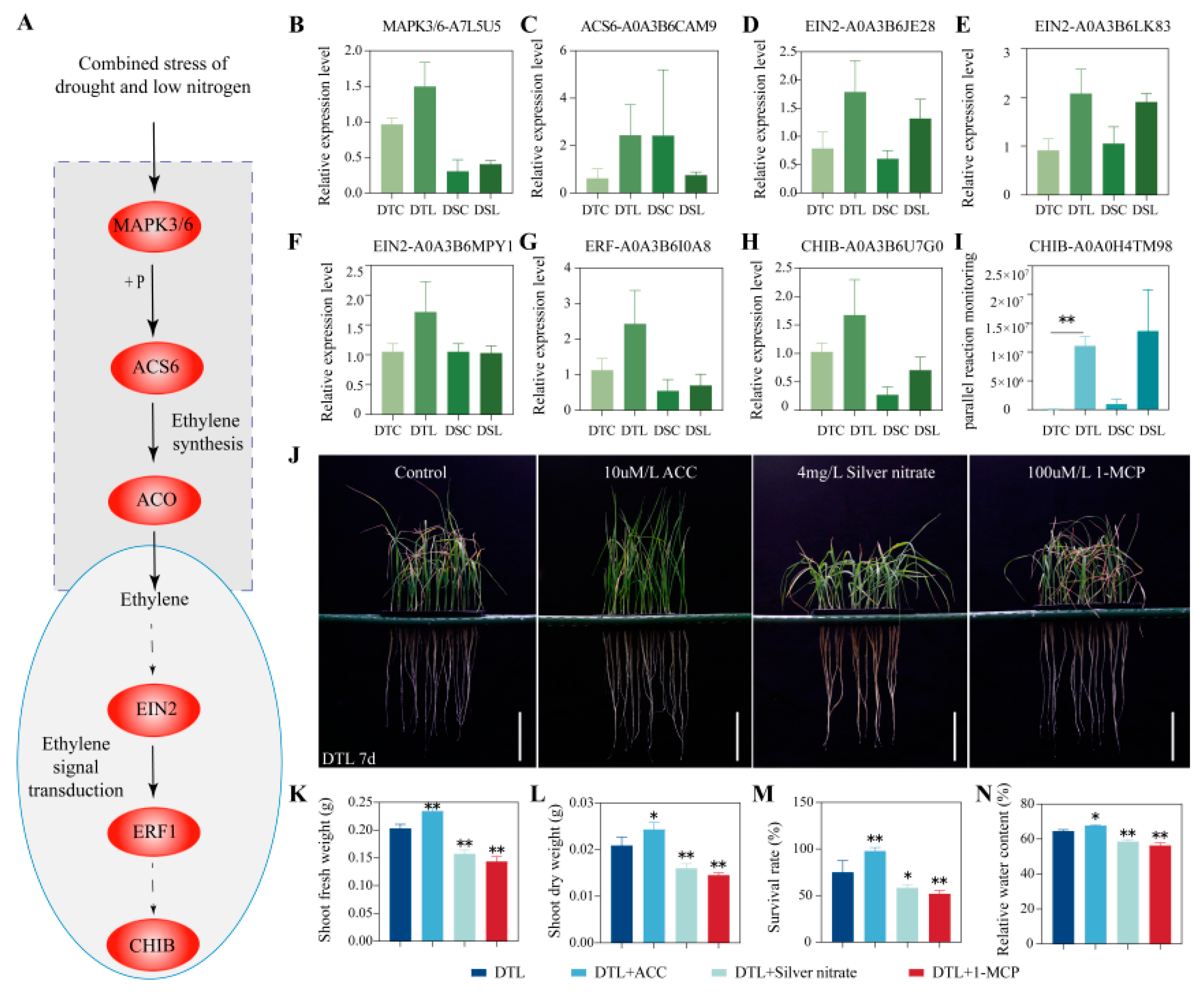
3.3. In Chang6878, Ethylene Biosynthesis-Related and Signal Transduction Proteins Were Significantly Upregulated in the MAPK Signaling Pathway
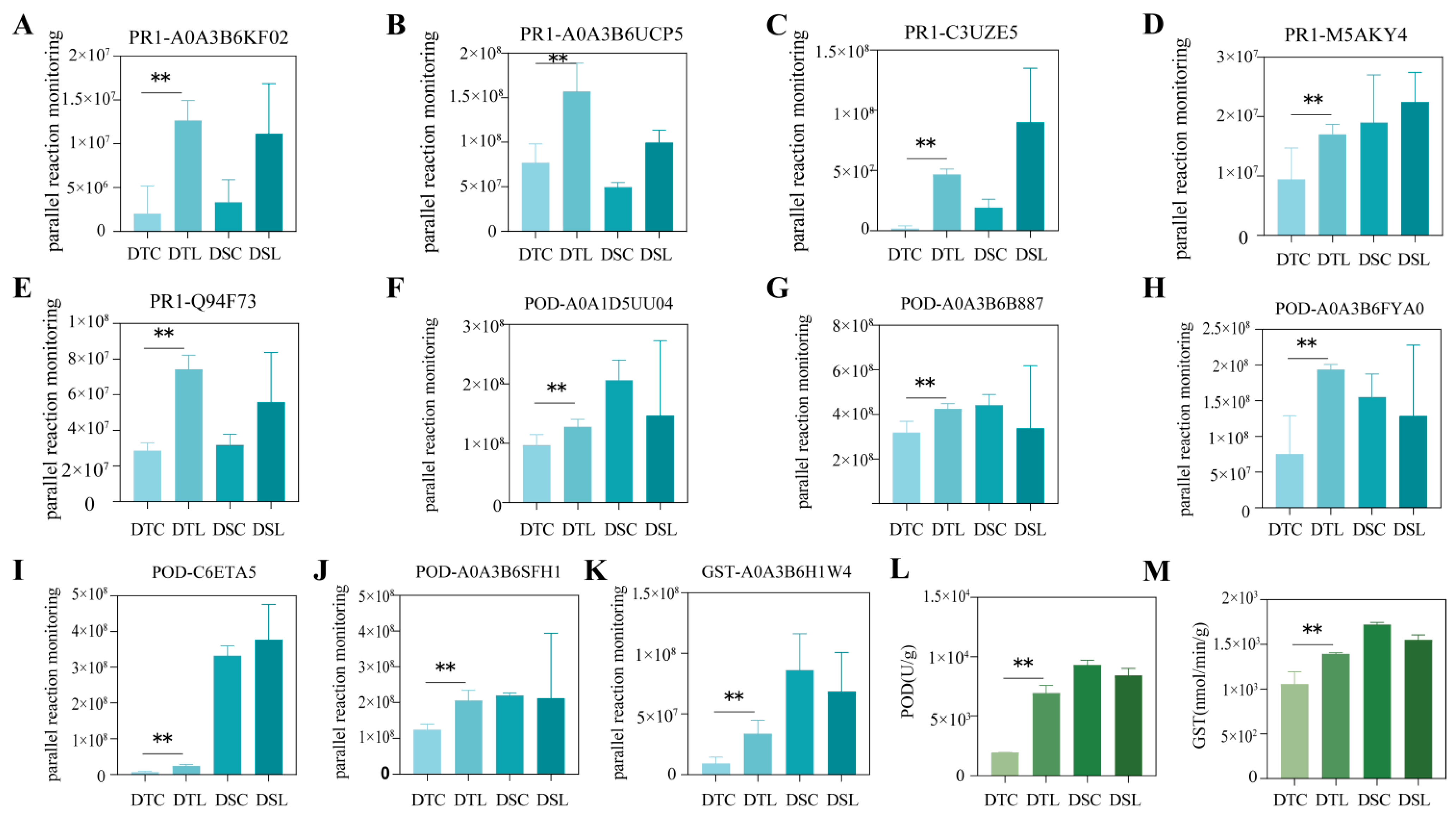
3.4. Enhancement of Oxidation Reaction-Related DAPs in Response to Combined Drought and LN Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lucertini, G.; Giustino, G.D. Urban and peri-urban agriculture as a tool for food security and climate change mitigation and adaptation: The case of Mestre. Sustainability 2021, 13, 5999. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Change 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Paymard, P.; Yaghoubi, F.; Nouri, M.; Bannayan, M. Projecting climate change impacts on rainfed wheat yield, water demand, and water use efficiency in northeast Iran. Theor. Appl. Climatol. 2019, 138, 1361–1373. [Google Scholar] [CrossRef]
- Lhn, A.; Rins, A.; Gidst, A.; Dgda, B.; Wvds, B.; Hap, A. Leaf gas exchange, photochemical responses and oxidative damages in assai (Euterpe oleracea Mart.) seedlings subjected to high temperature stress. Sci. Hortic. 2019, 257, 108733. [Google Scholar]
- Ramond, J.B.; Jordaan, K.; Díez, B.; Heinzelmann, S.M.; Cowan, D.A. Microbial biogeochemical cycling of nitrogen in arid ecosystems. Microbiol. Mol. Biol. Rev. 2022, 86, e0010921. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Wu, J.; Huang, M.; Li, Y.; Fu, G.; Tian, W. Effects of water-limited irrigation on grain yield, protein content and water nitrogen utilization efficiency of extremely late sowing densely planted wheat. J. Wheat Crops 2017, 10, 1349. [Google Scholar]
- Han, M.; Lv, Q.; Zhang, J.; Wang, T.; Zhang, C.; Tan, R.; Wang, Y.; Zhong, L.; Gao, Y.; Chao, Z. Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of Nitrate Reductase 1.2 in rice. Mol. Plant 2021, 15, 167–178. [Google Scholar] [CrossRef]
- Saneoka, H.; Moghaieb, R.; Premachandra, G.S.; Fujita, K. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Ru, C.; Wang, K.; Hu, X.; Chen, D.; Wang, W.; Yang, H. Nitrogen modulates the effects of heat, drought, and combined stresses on photosynthesis, antioxidant capacity, cell osmoregulation, and grain yield in winter wheat. J. Plant Growth Regul. 2022, 14, 1407. [Google Scholar] [CrossRef]
- Si, Z.; Zain, M.; Mehmood, F.; Wang, G.; Gao, Y.; Duan, A. Effects of nitrogen application rate and irrigation regime on growth, yield, and water-nitrogen use efficiency of drip-irrigated winter wheat in the North China Plain. Agric. Water Manag. 2020, 231, 106002. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Chen, J.; Chen, A.; Wang, L.; Guo, X.; Niu, Y.; Liu, S.; Mi, G.; Gao, Q. Reducing basal nitrogen rate to improve maize seedling growth, water and nitrogen use efficiencies under drought stress by optimizing root morphology and distribution. Agric. Water Manag. 2019, 212, 328–337. [Google Scholar] [CrossRef]
- Li, G.; Tillard, P.; Gojon, A.; Christopher, M. Dual regulation of root hydraulic conductivity and plasma membrane aquaporins by plant nitrate accumulation and high-affinity nitrate transporter NRT2.1. Plant Cell Physiol. 2016, 57, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liu, J.; Guo, J.; Wang, Y.; Ji, H.; Chu, X.; Xiao, K.; Qi, X.; Hu, L.; Li, H.; et al. GmTDN1 improves wheat yields by inducing dual tolerance to both drought and low-N stress. Plant Biotechnol. J. 2022, 20, 1606–1621. [Google Scholar] [CrossRef]
- Yang, C.; Ma, B.; He, S.J.; Xiong, Q.; Duan, K.X.; Yin, C.C.; Chen, H.; Lu, X.; Chen, S.Y.; Zhang, J.S. MAOHUZI6/ETHYLENE INSENSITIVE3-LIKE1 and ETHYLENE INSENSITIVE3-LIKE2 regulate ethylene response of roots and coleoptiles and negatively affect salt tolerance in rice. Plant Physiol. 2015, 169, 148–165. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lu, X.; Li, M.; Yang, S.; Wen, M.; Zhao, Y.; Liu, Y.; Ma, F. Effects of ethylene inhibitors on cotton growth and yield formation. Xinjiang Agric. Sci. 2020, 57, 1009–1017. [Google Scholar]
- Zhu, L.; Wang, D.; Sun, J.; Mu, Y.; Pu, W.; Ma, B.; Ren, F.; Yan, W.; Zhang, Z.; Li, G.; et al. Phenotypic and proteomic characteristics of sorghum (Sorghum bicolor) albino lethal mutant sbe6-a1. Plant Physiol. Biochem. 2019, 139, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Mu, Y.; Hu, X.; Ma, B.; Pan, Y. Comparative proteomic analysis reveals that the heterosis of two maize hybrids is related to enhancement of stress response and photosynthesis respectively. BMC Plant Biol. 2020, 21, 34. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Luo, M.; Sun, X.; Yan, J.; Shi, H.; Yan, H.; Yan, R.; Wang, S.; Tang, W.; Zhou, Y.; et al. SiMYB19 from foxtail millet (Setaria italica) confers transgenic rice tolerance to high salt stress in the field. Int. J. Mol. Sci. 2022, 23, 756. [Google Scholar] [CrossRef]
- Yang, J.; Wang, M.; Li, W.; He, X.; Teng, W.; Ma, W.; Zhao, X.; Hu, M.; Li, H.; Zhang, Y.; et al. Reducing expression of a nitrate-responsive bZIP transcription factor increases grain yield and N use in wheat. Plant Biotechnol. J. 2019, 17, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.E.; Craine, J.M.; Lany, N.K.; Jonard, M.; Ollinger, S.V.; Groffman, P.M.; Fulweiler, R.W.; Angerer, J.; Read, Q.D.; Reich, P.B. Evidence, causes, and consequences of declining nitrogen availability in terrestrial ecosystems. Science 2022, 376, eabh3767. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.D.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y. Independent and combined influence of drought stress and nitrogen deficiency on physiological and proteomic changes of barley leaves. Environ. Exp. Bot. 2023, 210, 105346. [Google Scholar] [CrossRef]
- Jin, K.; Chen, G.; Yoo, M.J.; Zhu, N.; Chen, S. Comparative proteomic analysis of Brassica napus in response to drought stress. J. Proteome Res. 2015, 14, 3068–3081. [Google Scholar]
- Wen, D.; Xu, H.; He, M.; Zhang, C. Proteomic analysis of wheat seeds produced under different nitrogen levels before and after germination. Food Chem. 2020, 340, 127937. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Raina, S.K.; Sultan, S.M. Arabidopsis MAPK signaling pathways and their cross talks in abiotic stress response. J. Plant Biochem. Biotechnol. 2020, 29, 700–714. [Google Scholar] [CrossRef]
- Li, S.; Han, X.; Yang, L.; Deng, X.; Wu, H.; Zhang, M.; Liu, Y.; Zhang, S.; Xu, J. Mitogen-activated protein kinases and calcium-dependent protein kinases are involved in wounding-induced ethylene biosynthesis in Arabidopsis thaliana. Plant Cell Environ. 2018, 41, 134–147. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, J.; Zhang, H.; Zhang, Z.; Quan, R.; Zhou, S.; Huang, R. Transcriptional activation of OsDERF1 in OsERF3 and OsAP2-39 negatively modulates ethylene synthesis and drought tolerance in rice. PLoS ONE 2011, 6, e25216. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Ma, T.; Xian, W.; Hu, B.; Chu, C. Interplay between ethylene and nitrogen nutrition: How ethylene orchestrates nitrogen responses in plants. J. Integr. Plant Biol. 2022, 65, 399–407. [Google Scholar] [CrossRef]
- Khan, M.I.; Trivellini, A.; Fatma, M.; Masood, A.; Francini, A.; Iqbal, N.; Ferrante, A.; Khan, N.A. Role of ethylene in responses of plants to nitrogen availability. Front. Plant Sci. 2015, 6, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Liu, Y.; Qin, G.; Wu, P.; Zi, H.; Xu, Z.; Zhao, X.; Wang, Y.; Li, Y.; Yang, S.; et al. The TOR–EIN2 axis mediates nuclear signalling to modulate plant growth. Nature 2021, 591, 288–292. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, C.; Li, K.; Sun, F.; Hu, H.; Li, X.; Zhao, Y.; Han, C.; Zhang, W.; Duan, Y.; et al. Arabidopsis EIN2 modulates stress response through abscisic acid response pathway. Plant Mol. Biol. 2007, 64, 633–644. [Google Scholar] [CrossRef]
- Zheng, D.; Han, X.; An, Y.I.; Guo, H.; Xia, X.; Yin, W. The nitrate transporter NRT2.1 functions in the ethylene response to nitrate deficiency in Arabidopsis. Plant Cell Environ. 2013, 36, 1328–1337. [Google Scholar] [CrossRef]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef]
- De Gernier, H.; De Pessemier, J.; Xu, J.; Cristescu, S.M.; Van Der Straeten, D.; Verbruggen, N.; Hermans, C. A comparative study of ethylene emanation upon nitrogen deficiency in natural accessions of Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Gao, G.; Chen, P.; Chen, K.; Wang, X.; Bai, L.; Yu, C.; Zhu, A. Integrative transcriptome and proteome analysis identifies major molecular regulation pathways involved in ramie (Boehmeria nivea (L.) Gaudich) under nitrogen and water co-limitation. Plants 2020, 9, 1267. [Google Scholar] [CrossRef]
- Chu, N.; Zhou, J.R.; Rott, P.C.; Li, J.; Fu, H.Y.; Huang, M.T.; Zhang, H.L.; Gao, S.J. ScPR1 plays a positive role in the regulation of resistance to diverse stresses in sugarcane (Saccharum spp.) and Arabidopsis thaliana. Ind. Crops Prod. 2022, 180, 114736. [Google Scholar] [CrossRef]
- Liu, T.; Chen, T.; Kan, J.; Yao, Y.; Guo, D.; Yang, Y.; Ling, X.; Wang, J.; Zhang, B. The GhMYB36 transcription factor confers resistance to biotic and abiotic stress by enhancing PR1 gene expression in plants. Plant Biotechnol. J. 2022, 20, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Rakwal, R.; Jwa, N.S.; Agrawal, V.P. Signaling molecules and blast pathogen attack activates rice OsPR1a and OsPR1b genes: A model illustrating components participating during defence/stress response. Plant Physiol. Biochem. 2001, 39, 1095–1103. [Google Scholar] [CrossRef]
- Zhong, C.; Bai, Z.G.; Zhu, L.F.; Zhang, J.H.; Zhu, C.Q.; Huang, J.L.; Jin, Q.Y.; Cao, X.C. Nitrogen-mediated alleviation of photosynthetic inhibition under moderate water deficit stress in rice (Oryza sativa L.). Environ. Exp. Bot. 2019, 157, 269–282. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, J.; Wang, D.; Kang, S.; He, Z.; Li, X.; Tang, W.; Chen, K.; Pan, Y.; Zhou, Y.; Xu, Z.; et al. Ethylene Regulates Combined Drought and Low Nitrogen Tolerance in Wheat: Proteomic Analysis. Agronomy 2023, 13, 1950. https://doi.org/10.3390/agronomy13071950
Yan J, Wang D, Kang S, He Z, Li X, Tang W, Chen K, Pan Y, Zhou Y, Xu Z, et al. Ethylene Regulates Combined Drought and Low Nitrogen Tolerance in Wheat: Proteomic Analysis. Agronomy. 2023; 13(7):1950. https://doi.org/10.3390/agronomy13071950
Chicago/Turabian StyleYan, Jiji, Daoping Wang, Shuyu Kang, Zhang He, Xin Li, Wensi Tang, Kai Chen, Yinghong Pan, Yongbin Zhou, Zhaoshi Xu, and et al. 2023. "Ethylene Regulates Combined Drought and Low Nitrogen Tolerance in Wheat: Proteomic Analysis" Agronomy 13, no. 7: 1950. https://doi.org/10.3390/agronomy13071950