
Postharvest Preservation of Flammulina velutipes with Isoamyl Isothiocyanate
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Preparation
2.3. Treatment Preparation
2.4. CO2 Concentration
2.5. Weight Loss and Browning Degree
2.6. Histological Microstructure
2.6.1. Histological Section
2.6.2. Scanning Electron Microscopy (SEM)
2.7. Chitin Content
2.8. Enzyme Activities
2.8.1. PPO Activity
2.8.2. TYR Activity
2.8.3. POD Activity
2.8.4. CAT Activity
2.8.5. SOD Activity
2.8.6. Succinic Dehydrogenase (SDH) Activity
2.9. Microbiological Analysis
2.9.1. Total Colony Count
2.9.2. Deoxyribonucleic Acid (DNA) Extraction and Sequencing Library Construction
2.10. Contents of Total Phenolics and Ascorbic Acid
2.11. Malondialdehyde (MDA) Content
2.12. Nutritional Content
2.13. Statistical Analysis
3. Results
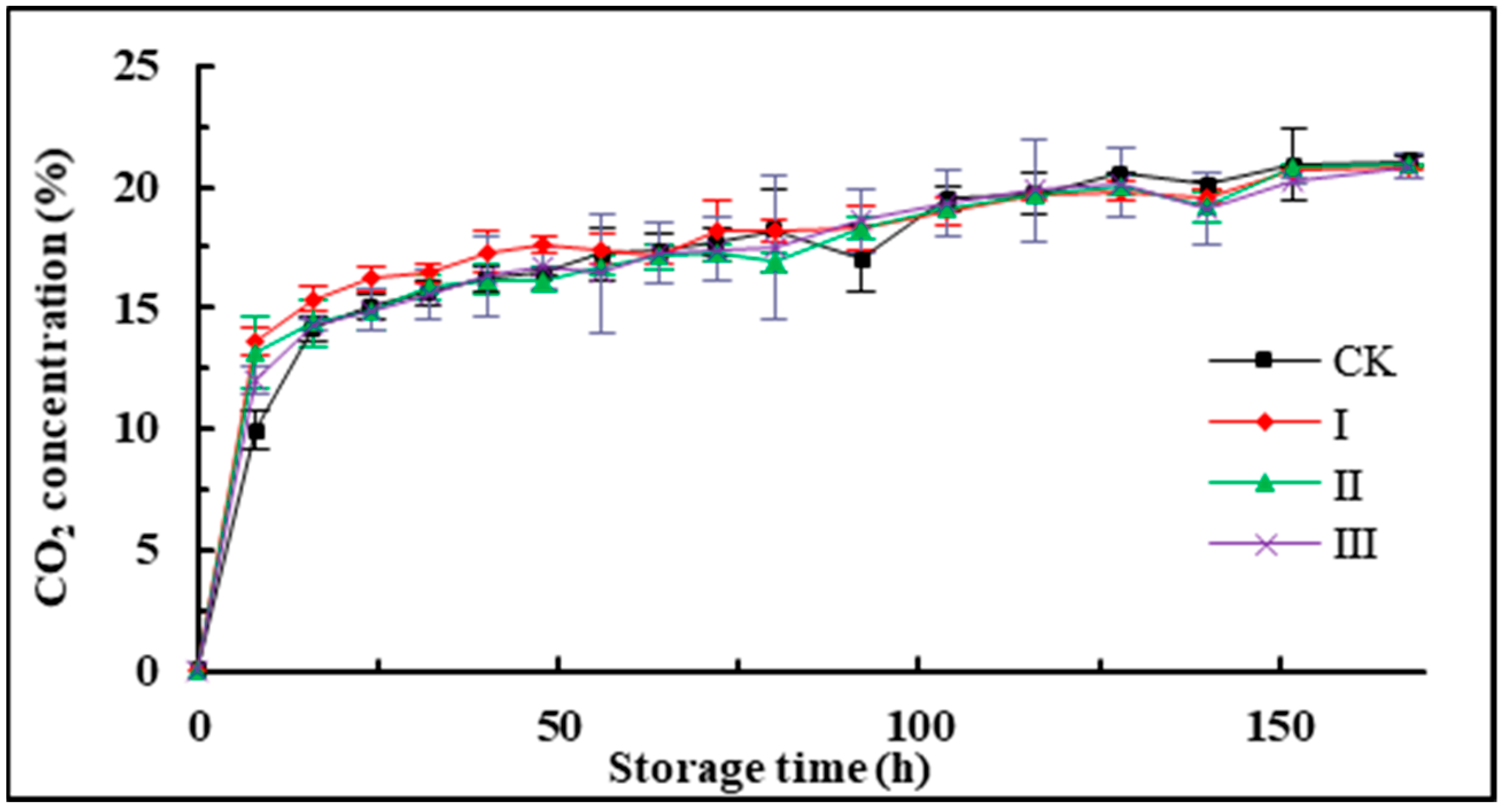
3.1. CO2 Concentration
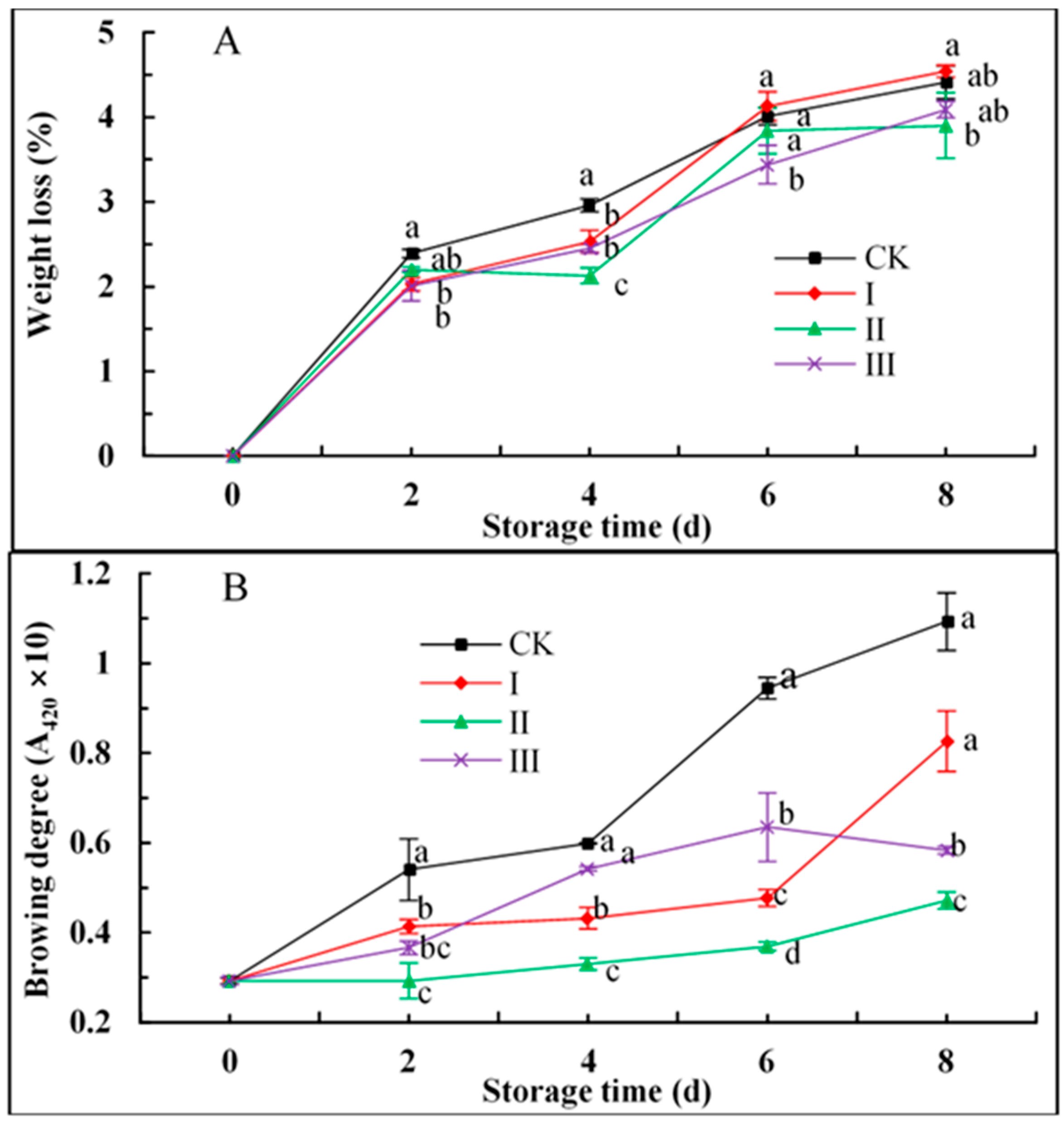
3.2. Weight Loss
3.3. Browning Degree
3.4. Appearance and Microstructure Observation
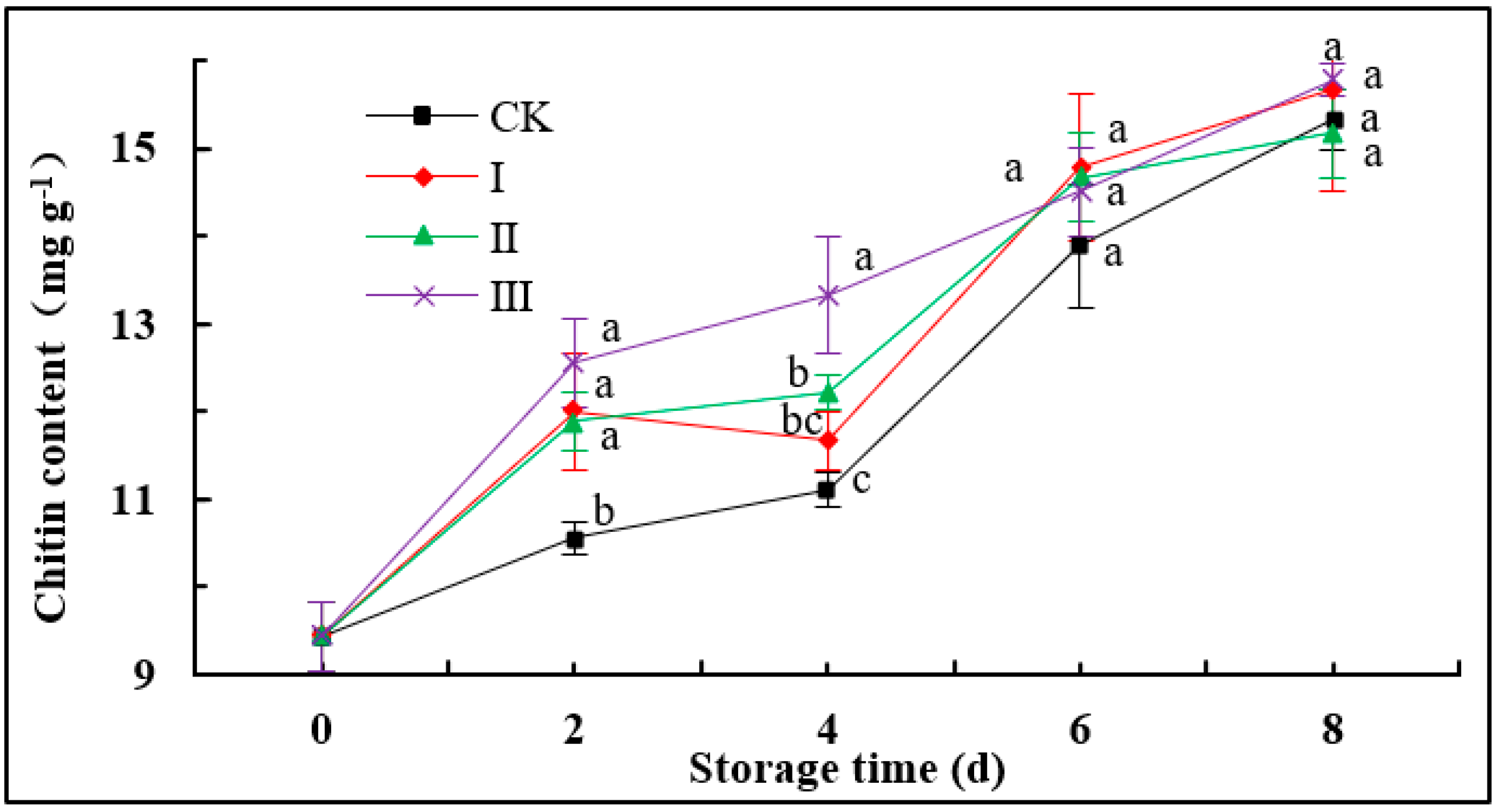
3.5. Chitin Content
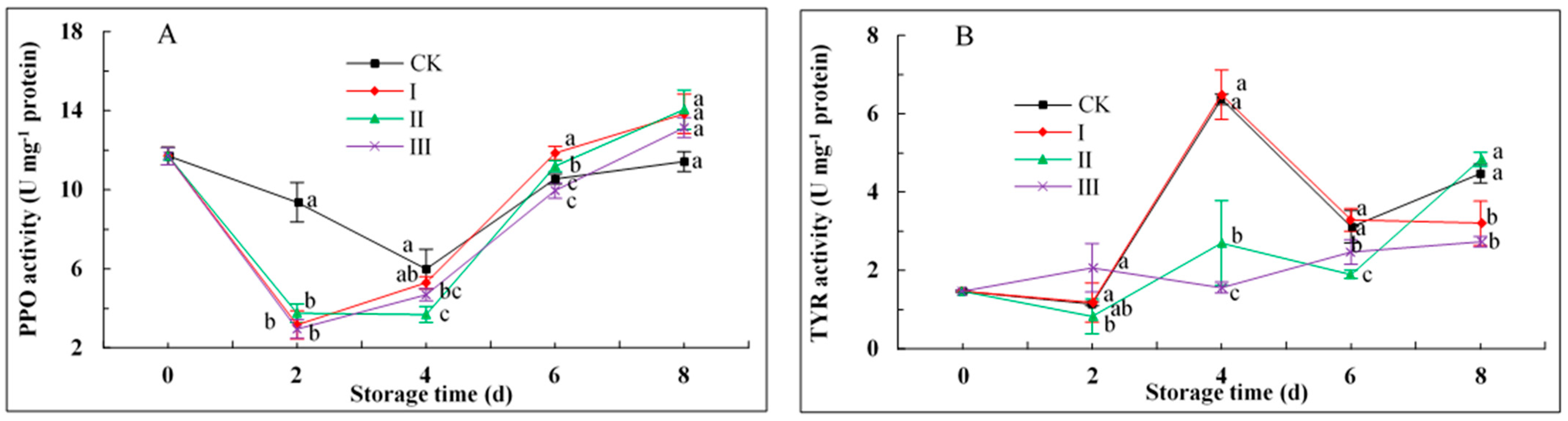
3.6. PPO and TYR Activities
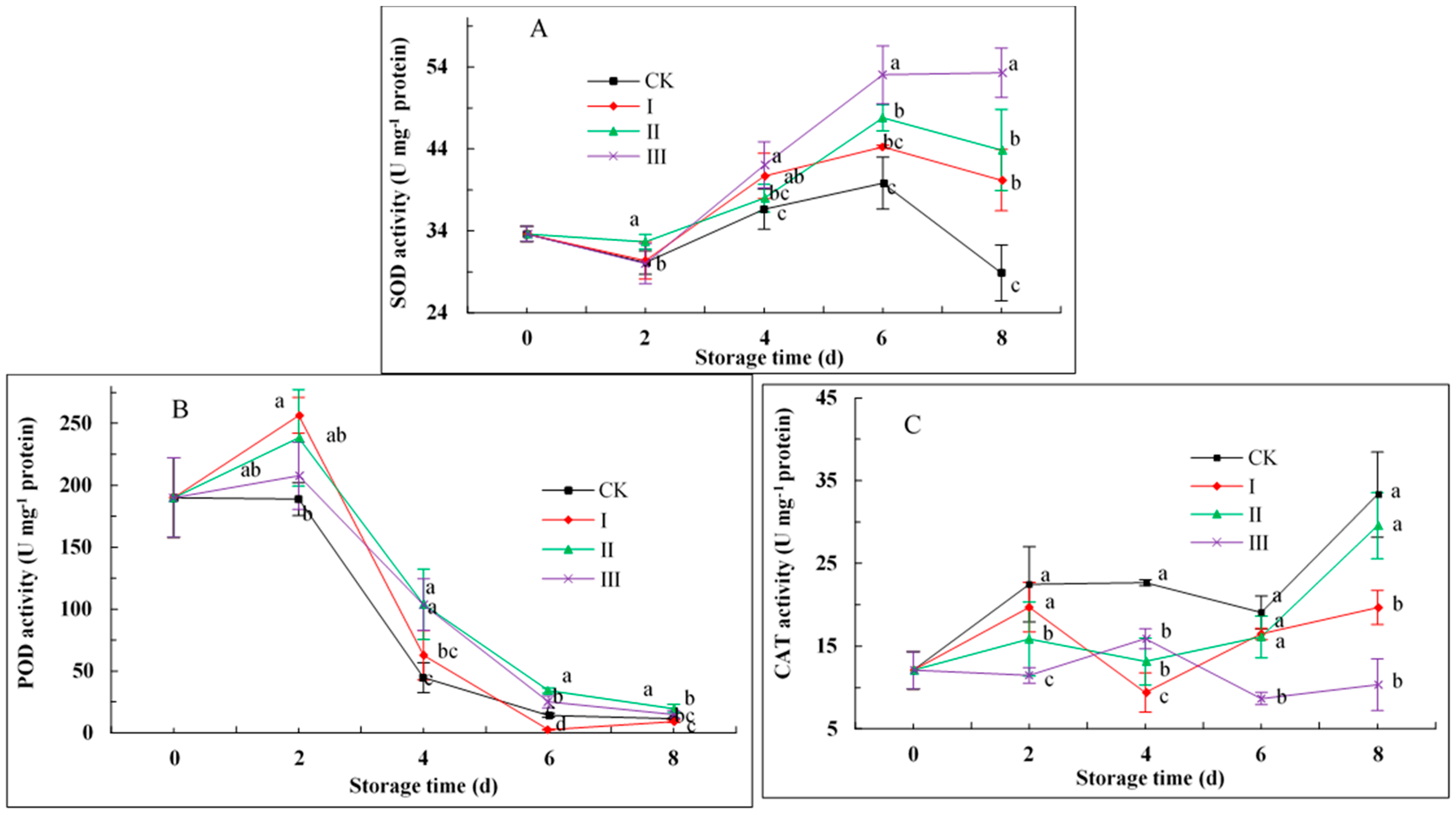
3.7. SOD, POD, and CAT Activities
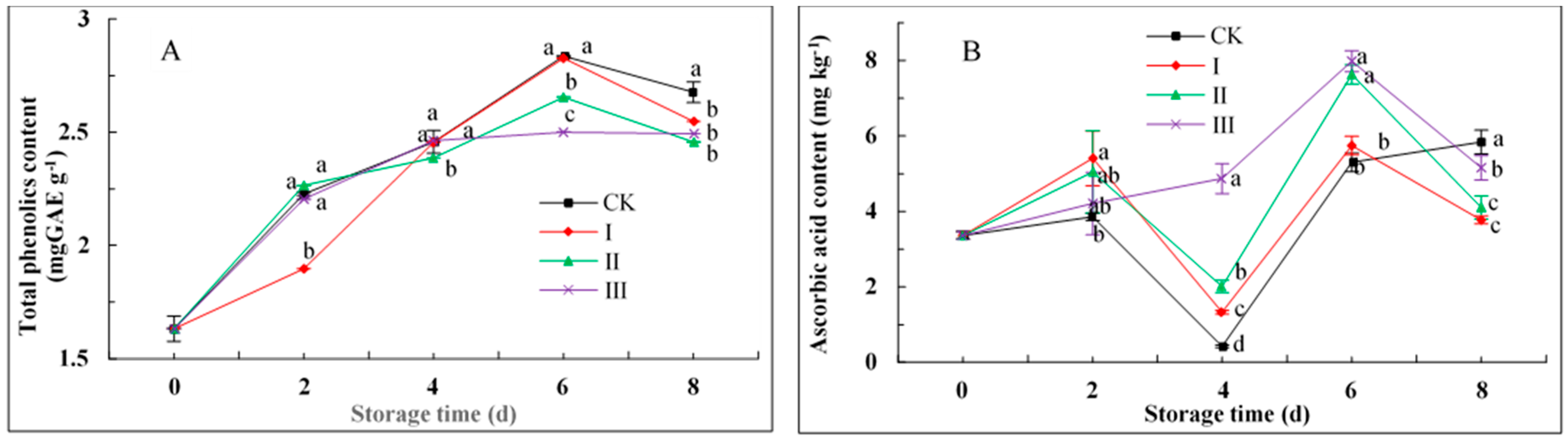
3.8. Total Phenolic and Ascorbic Acid Contents
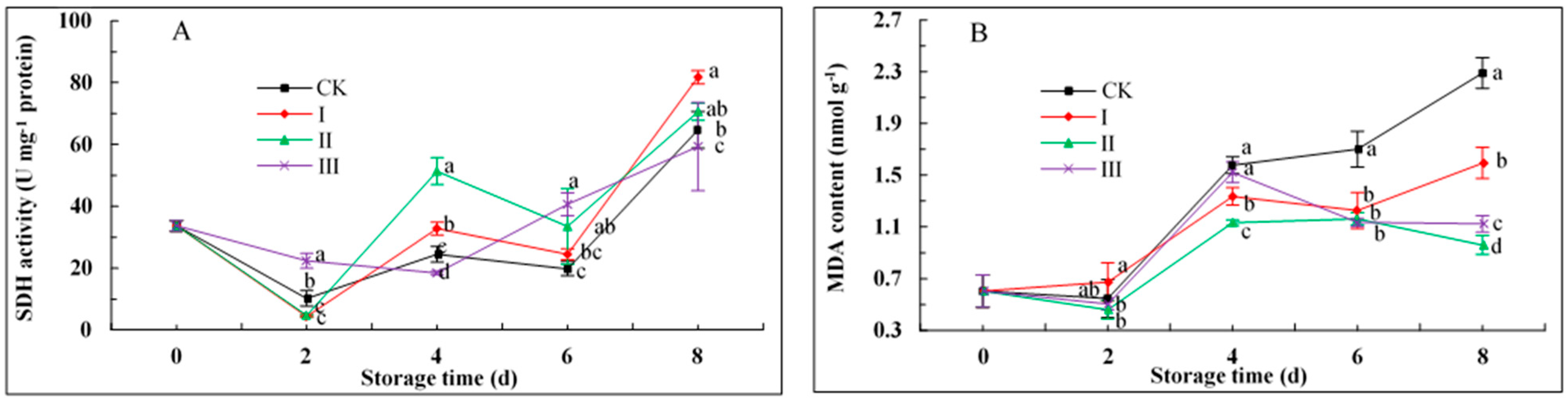
3.9. SDH Activity and MDA Content
3.10. Bacterial Colony Counts
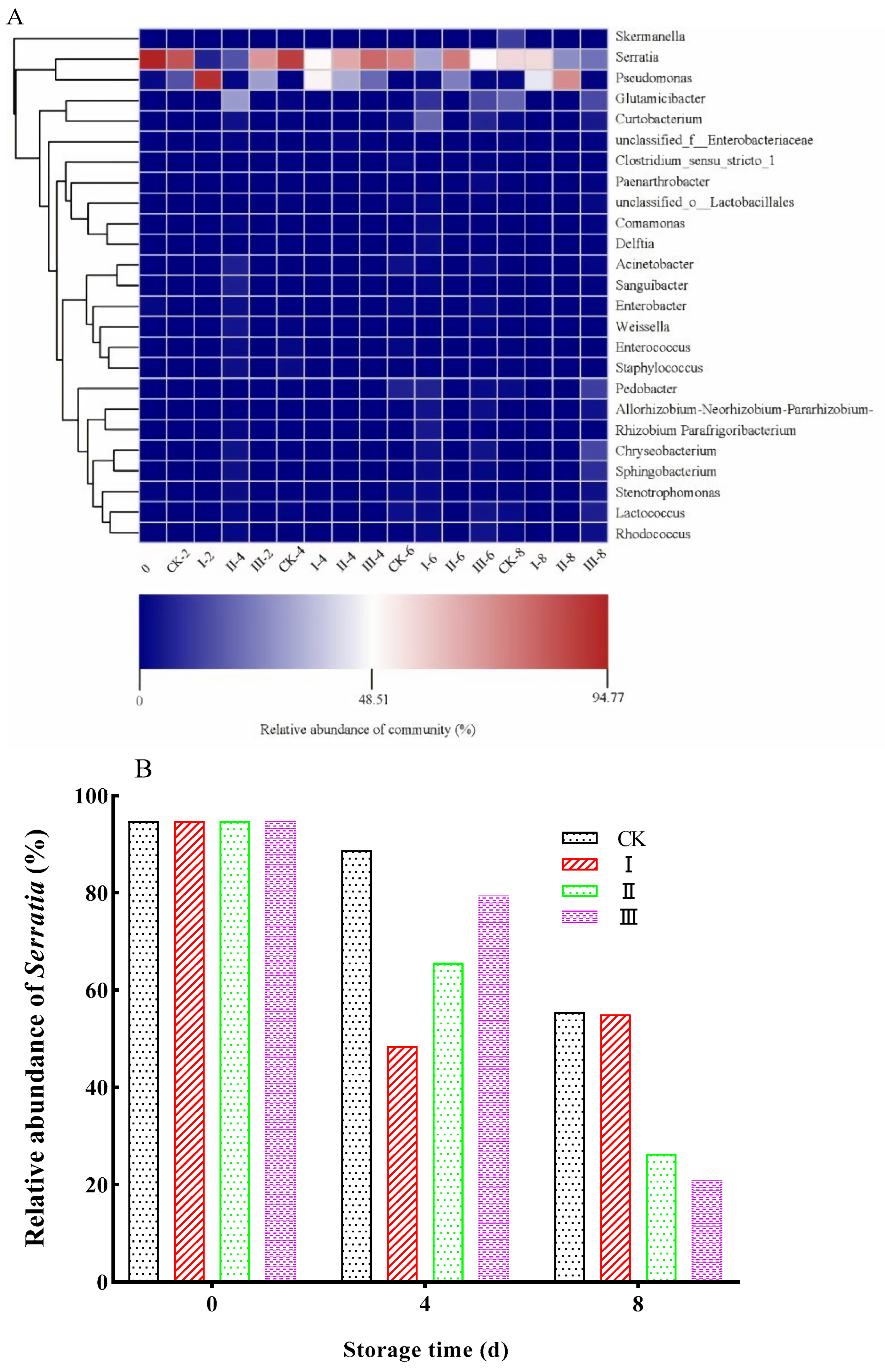
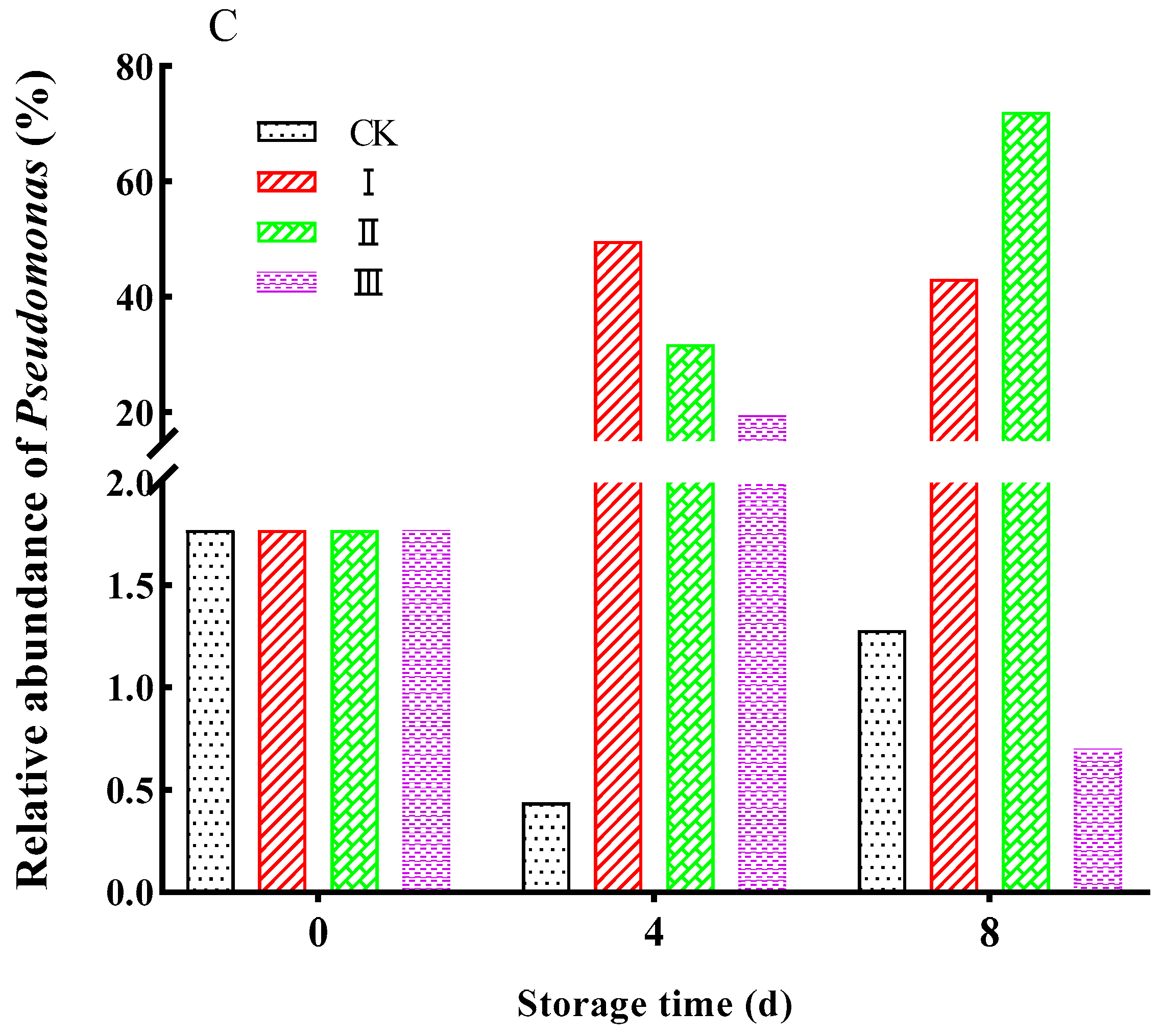
3.11. Bacterial Community Structure
3.12. Amino Acid and Fatty Acid Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Marçal, S.; Sousa, A.S.; Taofiq, O.; Antunes, F.; Morais, A.M.M.B.; Freitas, A.C.; Barros, L.; Ferreira, I.C.F.R.; Pintado, M. Impact of postharvest preservation methods on nutritional value and bioactive properties of mushrooms. Trends Food Sci. Technol. 2021, 110, 418–431. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current overview of mushroom production in the world. Edible Med. Mushrooms Technol. Appl. 2017, 2, 5–13. [Google Scholar] [CrossRef]
- Shi, C.; Wu, Y.; Fang, D.; Pei, F.; Mariga, A.M.; Yang, W.; Hu, Q. Effect of nanocomposite packaging on postharvest senescence of Flammulina velutipes. Food Chem. 2018, 246, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Tano, K.; Arul, J.; Doyon, G.; Castaigne, F. Atmospheric composition and quality of fresh mushrooms in modified atmosphere packages as affected by storage temperature abuse. J. Food Sci. 1999, 64, 1073–1077. [Google Scholar] [CrossRef]
- Fang, D.L.; Yang, W.J.; Kimatu, B.M.; Mariga, A.M.; Zhao, L.Y.; An, X.X.; Hu, Q.H. Effect of nanocomposite-based packaging on storage stability of mushrooms (Flammulina velutipes). Innov. Food Sci. Emerg. Technol. 2016, 33, 489–497. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Tian, Y.; Ma, R.N.; Liu, Q.H.; Zhang, J. Effect of plasma activated water on the postharvest quality of button mushrooms, Agaricus bisporus. Food Chem. 2016, 197, 436–444. [Google Scholar] [CrossRef]
- Zhang, K.X.; Pu, Y.Y.; Sun, D.W. Recent advances in quality preservation of postharvest mushrooms (Agaricus bisporus): A review. Trends Food Sci. Technol. 2018, 78, 72–82. [Google Scholar] [CrossRef]
- Li, N.; Wu, X.; Zhuang, W.; Wu, C.; Rao, Z.; Du, L.; Zhou, Y. Cruciferous vegetable and isothiocyanate intake and multiple health outcomes. Food Chem. 2022, 375, 131816. [Google Scholar] [CrossRef]
- Kyung, K.H. Antimicrobial properties of allium species. Curr. Opin. Biotechnol. 2012, 23, 142–147. [Google Scholar] [CrossRef]
- Wilson, A.E.; Bergaentzlé, M.; Bindler, F.; Marchioni, E.; Lintz, A.; Ennahar, S. In vitro efficacies of various isothiocyanates from cruciferous vegetables as antimicrobial agents against foodborne pathogens and spoilage bacteria. Food Control 2013, 30, 318–324. [Google Scholar] [CrossRef]
- Chen, H.; Gao, H.; Fang, X.; Ye, L.; Zhou, Y.; Yang, H. Effects of allyl isothiocyanate treatment on postharvest quality and the activities of antioxidant enzymes of mulberry fruit. Postharvest Biol. Technol. 2015, 108, 61–67. [Google Scholar] [CrossRef]
- Park, D.; Park, S.Y.; Liu, K.H.; Ku, K.M. Optimal allyl isothiocyanate concentration on Botrytis cinerea during the postharvest storage of blackberries and mechanism of blackberry color changes at high concentration of allyl isothiocyanate. Postharvest Biol. Technol. 2023, 199, 112292. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Xu, Y.; Chen, T.; Li, B.; Zhang, Z.; Tian, S. Application and mechanism of benzyl-isothiocyanate, a natural antimicrobial agent from cruciferous vegetables, in controlling postharvest decay of strawberry. Postharvest Biol. Technol. 2021, 180, 111604. [Google Scholar] [CrossRef]
- Wu, T.L.; Hu, Y.M.; Sun, Y.; Zhang, Z.J.; Wu, Z.R.; Zhao, W.B.; Tang, C.; Du, S.S.; He, Y.H.; Ma, Y.; et al. Insights into the mode of action of 2-(4-methoxyphenyl) ethyl isothiocyanate on Aspergillus niger. Food Control 2022, 136, 108871. [Google Scholar] [CrossRef]
- Lei, J.; Li, B.; Zhang, N.; Yan, R.; Guan, W.; Brennan, C.S.; Gao, H.; Peng, B. Effects of UV-C treatment on browning and the expression of polyphenol oxidase (PPO) genes in different tissues of Agaricus bisporus during cold storage. Postharvest Biol. Technol. 2018, 139, 99–105. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, Y.; Gao, J. Anatomy and microscopic characteristics of Picris japonica. Rev. Bras. Farmacogn. 2018, 28, 640–646. [Google Scholar] [CrossRef]
- Hassainia, A.; Satha, H.; Boufi, S. Chitin from Agaricus bisporus: Extraction and characterization. Int. J. Biol. Macromol. 2018, 117, 1334–1342. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Zhang, F.; Hou, F.; Yi, F.; Sun, X.; Yang, Q.; Han, X.; Liu, Z. Characterization of chitosan/zein composite film combined with tea polyphenol and its application on postharvest quality improvement of mushroom (Lyophyllum decastes Sing.). Food Packag. Shelf Life 2022, 33, 100869. [Google Scholar] [CrossRef]
- Mahdavi, A.; Mohammadsadeghi, N.; Mohammadi, F.; Saadati, F.; Nikfard, S. Evaluation of inhibitory effects of some novel phenolic derivatives on the mushroom tyrosinase activity: Insights from spectroscopic analyses, molecular docking and in vitro assays. Food Chem. 2022, 387, 132938. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, Y.; Liu, Z.; Huang, X.; Yi, F.; Hou, F.; Zhang, F. Preparation and characterization of chitosan/zein film loaded with lemon essential oil: Effects on postharvest quality of mushroom (Agaricus bisporus). Int. J. Biol. Macromol. 2021, 192, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Yuan, B.; Xie, Y.; Cheng, S.; Huang, H.; Zhang, W.; Chen, J.; Cao, C. Improvement of postharvest quality, enzymes activity and polyphenoloxidase structure of postharvest Agaricus bisporus in response to high voltage electric field. Postharvest Biol. Technol. 2020, 166, 111230. [Google Scholar] [CrossRef]
- Roy, S.; Anantheswaran, R.C.; Beelman, R.B. Fresh mushroom quality as affected by modified atmosphere packaging. J. Food Sci. 1995, 60, 334–340. [Google Scholar] [CrossRef]
- Mau, J.L.; Lin, H.C.; Chen, C.C. Non-volatile components of several medicinal mushrooms. Food Res. Int. 2001, 34, 521–526. [Google Scholar] [CrossRef]
- Sissons, J.; Davila, M.; Du, X. Sautéing and roasting effect on free amino acid profiles in portobello and shiitake mushrooms, and the effect of mushroom- and cooking-related volatile aroma compounds on meaty flavor enhancement. Int. J. Gastron. Food Sci. 2022, 28, 100550. [Google Scholar] [CrossRef]
- Zivanovic, S.; Busher, R.W.; Kim, K.S. Textural changes in mushrooms (Agaricus bisporus) associated with tissue ultrastructure and composition. J. Food Sci. 2000, 65, 1404–1408. [Google Scholar] [CrossRef]
- Xia, R.; Wang, L.; Xin, G.; Bao, X.; Sun, L.; Xu, H.; Hou, Z. Preharvest and postharvest applications of 1-MCP affect umami taste and aroma profiles of mushrooms (Flammulina velutipes). LWT 2021, 144, 111176. [Google Scholar] [CrossRef]
- Niu, Y.; Yun, J.; Bi, Y.; Wang, T.; Zhang, Y.; Liu, H.; Zhao, F. Predicting the shelf life of postharvest Flammulina velutipes at various temperatures based on mushroom quality and specific spoilage organisms. Postharvest Biol. Technol. 2020, 167, 111235. [Google Scholar] [CrossRef]
- Lin, X.; Sun, D.W. Research advances in browning of button mushroom (Agaricus bisporus): Affecting factors and controlling methods. Trends Food Sci. Technol. 2019, 90, 63–75. [Google Scholar] [CrossRef]
- Li, D.; Wang, D.; Fang, Y.; Li, L.; Lin, X.; Xu, Y.; Chen, H.; Zhu, M.; Luo, Z. A novel phase change coolant promoted quality attributes and glutamate accumulation in postharvest shiitake mushrooms involved in energy metabolism. Food Chem. 2021, 351, 129227. [Google Scholar] [CrossRef]
- Fu, Y.; Yu, Y.; Tan, H.; Wang, B.; Peng, W.; Sun, Q. Metabolomics reveals dopa melanin involved in the enzymatic browning of the yellow cultivars of East Asian golden needle mushroom (Flammulina filiformis). Food Chem. 2022, 370, 131295. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Polyphenol oxidases in plants-recent progress. Phytochemistry 1986, 26, 11–20. [Google Scholar] [CrossRef]
- Chen, Q.X.; Song, K.K.; Qiu, L.; Liu, X.D.; Huang, H.; Guo, H.Y. Inhibitory effects on mushroom tyrosinase by p-alkoxybenzoic acids. Food Chem. 2005, 91, 269–274. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Nasiri, M.; Barzegar, M.; Sahari, M.A.; Niakousari, M. Application of Tragacanth gum impregnated with Satureja khuzistanica essential oil as a natural coating for enhancement of postharvest quality and shelf life of button mushroom (Agaricus bisporus). Int. J. Biol. Macromol. 2018, 106, 218–226. [Google Scholar] [CrossRef]
- Húngaro, H.M.; Caturla, M.Y.R.; Horita, C.N.; Furtado, M.M.; Sant’Ana, A.S. Blown pack spoilage in vacuum-packaged meat: A review on clostridia as causative agents, sources, detection methods, contributing factors and mitigation strategies. Trends Food Sci. Technol. 2016, 52, 123–138. [Google Scholar] [CrossRef]
- Louis, E.; Villalobos-Carvajal, R.; Reyes-Parra, J.; Jara-Quijada, E.; Ruiz, C.; Andrades, P.; Gacitúa, J.; Beldarraín-Iznaga, T. Preservation of mushrooms (Agaricus bisporus) by an alginate-based-coating containing a cinnamaldehyde essential oil nanoemulsion. Food Packag. Shelf Life 2021, 28, 100662. [Google Scholar] [CrossRef]
- Gao, M.; Feng, L.; Jiang, T. Browning inhibition and quality preservation of button mushroom (Agaricus bisporus) by essential oils fumigation treatment. Food Chem. 2014, 149, 107–113. [Google Scholar] [CrossRef]
- Fang, D.; Wang, C.; Deng, Z.; Ma, N.; Hu, Q.; Zhao, L. Microflora and umami alterations of different packaging material preserved mushroom (Flammulina filiformis) during cold storage. Food Res. Int. 2021, 147, 110481. [Google Scholar] [CrossRef]
- Sande, D.; de Oliveira, G.P.; Moura, M.A.F.; Martins, B.d.A.; Lima, M.T.N.S.; Takahashi, J.A. Edible mushrooms as a ubiquitous source of essential fatty acids. Food Res. Int. 2019, 125, 108524. [Google Scholar] [CrossRef]
- Combet, E.; Eastwood, D.C.; Burton, K.S.; Combet, E.; Henderson, J.; Henderson, J.; Combet, E. Eight-carbon volatiles in mushrooms and fungi: Properties, analysis, and biosynthesis. Mycoscience 2006, 47, 317–326. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Storage Time (d) | ||||
|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | |
| CK | 4.36 ± 0.04 | 5.74 ± 0.11 a | 5.57 ± 0.01 a | 6.33 ± 0.06 a | 7.34 ± 0.04 a |
| Ⅰ | 4.36 ± 0.04 | 5.15 ± 0.01 b | 5.48 ± 0.03 b | 6.14 ± 0.03 b | 6.87 ± 0.06 c |
| II | 4.36 ± 0.04 | 5.12 ± 0.05 b | 5.41 ± 0.01 c | 6.03 ± 0.03 b | 6.76 ± 0.08 d |
| III | 4.36 ± 0.04 | 5.05 ± 0.02 b | 5.34 ± 0.01 d | 6.04 ± 0.04 b | 7.01 ± 0.01 b |
| Amino Acid (mg g−1) | 0 d | 8 d | ||||
|---|---|---|---|---|---|---|
| CK | I | II | III | |||
| MSG-like | Aspartic acid | 10.85 ± 0.06 | 11.59 ± 0.09 b | 11.67 ± 0.21 b | 11.71 ± 0.18 b | 12.85 ± 0.02 a |
| Glutamic acid | 23.67 ± 1.18 | 34.04 ± 0.56 b | 34.97 ± 0.67 b | 40.48 ± 0.67 a | 39.72 ± 0.05 a | |
| Total | 33.76 | 45.63 | 46.64 | 52.19 | 52.57 | |
| Sweet | Alanine | 6.63 ± 0.83 | 10.46 ± 0.02 a | 9.50 ± 0.18 b | 10.34 ± 0.17 a | 9.06 ± 0.10 c |
| Glycine | 5.36 ± 1.03 | 6.98 ± 0.03 b | 6.91 ± 0.12 b | 7.04 ± 0.12 b | 7.25 ± 0.01 a | |
| Serine | 6.00 ± 0.76 | 6.80 ± 0.05 c | 6.82 ± 0.12 c | 7.02 ± 0.11 b | 7.29 ± 0.02 a | |
| Threonine | 5.84 ± 0.43 | 7.43 ± 0.08 a | 7.19 ± 0.13 b | 7.36 ± 0.12 ab | 7.39 ± 0.02 a | |
| Total | 23.83 | 31.67 | 30.42 | 31.76 | 30.99 | |
| Bitter | Arginine | 5.38 ± 0.74 | 7.10 ± 0.20 b | 6.99 ± 0.09 b | 7.37 ± 0.12 a | 7.35 ± 0.02 a |
| Histidine | 3.14 ± 1.75 | 4.17 ± 0.06 a | 4.10 ± 0.05 ab | 4.19 ± 0.09 a | 4.03 ± 0.02 b | |
| Leucine | 7.87 ± 0.64 | 9.34 ± 0.05 a | 8.79 ± 0.23 b | 8.67 ± 0.21 b | 8.92 ± 0.08 b | |
| Phenylalanine | 5.23 ± 0.76 | 7.12 ± 0.04 a | 6.89 ± 0.12 b | 6.90 ± 0.14 b | 7.15 ± 0.12 a | |
| Methionine | 1.69 ± 0.04 | 1.82 ± 0.08 a | 1.89 ± 0.05 a | 1.88 ± 0.03 a | 1.91 ± 0.07 a | |
| Cysteine | 1.11 ± 0.04 | 1.92 ± 0.02 a | 1.82 ± 0.02 b | 1.80 ± 0.03 b | 1.90 ± 0.04 a | |
| Valine | 5.94 ± 0.59 | 7.81 ± 0.05 a | 7.27 ± 0.17 b | 7.31 ± 0.14 b | 7.28 ± 0.02 b | |
| Total | 30.36 | 39.28 | 37.75 | 38.12 | 38.54 | |
| Tasteless | Isoleucine | 4.61 ± 0.03 | 5.54 ± 0.04 a | 5.14 ± 0.20 b | 5.07 ± 0.18 b | 5.19 ± 0.02 b |
| Lysine | 7.63 ± 0.02 | 10.22 ± 0.07 b | 10.33 ± 0.19 b | 10.60 ± 0.18 a | 10.82 ± 0.07 a | |
| Proline | 5.12 ± 2.64 | 5.41 ± 0.11 b | 5.44 ± 0.14 ab | 5.23 ± 0.14 ab | 5.54 ± 0.07 a | |
| Tryptophan | 2.22 ± 0.28 | 2.42 ± 0.01 c | 2.40 ± 0.01 d | 2.47 ± 0.01 b | 2.53 ± 0.01 a | |
| Tyrosine | 4.58 ± 1.20 | 5.99 ± 0.11 a | 5.87 ± 0.13 a | 5.98 ± 0.17 a | 6.02 ± 0.04 a | |
| Total | 24.16 | 29.58 | 29.14 | 29.35 | 30.1 | |
| Total amino acids | 112.11 | 145.3 | 143.12 | 150.51 | 151.20 | |
| Fatty Acids (mg g−1) | 0 d | 8 d | ||||
|---|---|---|---|---|---|---|
| CK | I | II | III | |||
| Saturated fatty acids (SFA) | Lauric acid C12:0 | 0.16 ± 0.01 | 0.30 ± 0.02 b | 0.36 ± 0.01 a | 0.27 ± 0.01 c | 0.27 ± 0.02 c |
| Myristic acid C14:0 | 0.32 ± 0.03 | 0.38 ± 0.02 b | 0.47 ± 0.01 a | 0.38 ± 0.02 b | 0.38 ± 0.02 b | |
| Pentadecanoic acid C15:0 | 0.09 ± 0.01 | 0.22 ± 0.01 b | 0.24 ± 0.0 b | 0.29 ± 0.02 a | 0.23 ± 0.01 b | |
| Palmitic acid C16:0 | 2.76 ± 0.21 | 2.61 ± 0.15 b | 3.05 ± 0.04 a | 2.72 ± 0.16 b | 2.49 ± 0.13 b | |
| Heptadecanoic acid C17:0 | n.d. | n.d. | n.d. | 0.03 ± 0.01 a | 0.02 ± 0.01 b | |
| Stearic acid C18:0 | 0.22 ± 0.02 | 0.20 ± 0.01 c | 0.25 ± 0.01 a | 0.23 ± 0.01 b | 0.19 ± 0.01 c | |
| Arachidic acid C20:0 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Heneicosanoic acid C21:0 | n.d. | 0.20 ± 0.00 a | 0.20 ± 0.00 a | 0.20 ± 0.00 a | 0.20 ± 0.00 a | |
| Behenic acid C22:0 | 0.10 ± 0.01 | 0.90 ± 0.01 b | 0.11 ± 0.00 a | 0.11 ± 0.00 a | 0.09 ± 0.01 b | |
| Lignoceric acid C24:0 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Total | 3.65 | 3.82 | 4.50 | 4.01 | 3.69 | |
| Monounsaturated fatty acids (MUFA) | Palmitoleic acid C16:1 | 0.18 ± 0.02 | 0.33 ± 0.02 a | 0.30 ± 0.01 b | 0.30 ± 0.02 b | 0.24 ± 0.01 c |
| Oleic acid C18:1n9c | 0.78 ± 0.06 | 0.95 ± 0.07 b | 1.05 ± 0.02 a | 0.95 ± 0.06 b | 0.80 ± 0.03 c | |
| cis-11-Eicosenoic acid C20:1 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Erucic acid C22:1n9 | n.d. | 0.04 ± 0.00 d | 0.06 ± 0.01 b | 0.07 ± 0.01 a | 0.05 ± 0.01 c | |
| Nervonic acid C24:1 | n.d. | 0.20 ± 0.02 ab | 0.25 ± 0.01 a | 0.24 ± 0.03 a | 0.13 ± 0.10 c | |
| Total | 0.96 | 1.53 | 1.66 | 1.55 | 1.27 | |
| Polyunsaturated fatty acids (PUFA) | Linoleic acid C18:2n6c | 8.34 ± 0.34 | 11.18 ± 0.74 b | 12.66 ± 0.17 a | 12.15 ± 0.77 ab | 11.13 ± 0.44 b |
| α-linoleic acid C18:3n3 | 4.53 ± 0.41 | 5.17 ± 0.34 b | 5.63 ± 0.09 a | 4.90 ± 0.29 bc | 4.46 ± 0.17 c | |
| cis-11,14,17-Eicosatrienoic acid C20:3n3 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Total | 13.83 | 16.35 | 18.32 | 17.05 | 15.59 | |
| Total fatty acids | 18.44 | 21.69 | 24.48 | 22.67 | 20.55 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, P.; Wang, P.; Teng, Q.; Chen, T.; Tian, G.; Yao, C.; Yalimaimaiti, N.; Liu, Q. Postharvest Preservation of Flammulina velutipes with Isoamyl Isothiocyanate. Agronomy 2023, 13, 1771. https://doi.org/10.3390/agronomy13071771
Zhu P, Wang P, Teng Q, Chen T, Tian G, Yao C, Yalimaimaiti N, Liu Q. Postharvest Preservation of Flammulina velutipes with Isoamyl Isothiocyanate. Agronomy. 2023; 13(7):1771. https://doi.org/10.3390/agronomy13071771
Chicago/Turabian StyleZhu, Peilin, Peng Wang, Qian Teng, Tong Chen, Guoting Tian, Chunxin Yao, Nuerziya Yalimaimaiti, and Qinghong Liu. 2023. "Postharvest Preservation of Flammulina velutipes with Isoamyl Isothiocyanate" Agronomy 13, no. 7: 1771. https://doi.org/10.3390/agronomy13071771