
Management of Eleusine indica (L.) Gaertn Resistance to Glyphosate Herbicide in Indonesia
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Herbicide Dose–Response Experiments
2.3. Dose–Response Experiments Statistical Analyses
2.4. DNA Isolation and EPSPS Gene Sequence
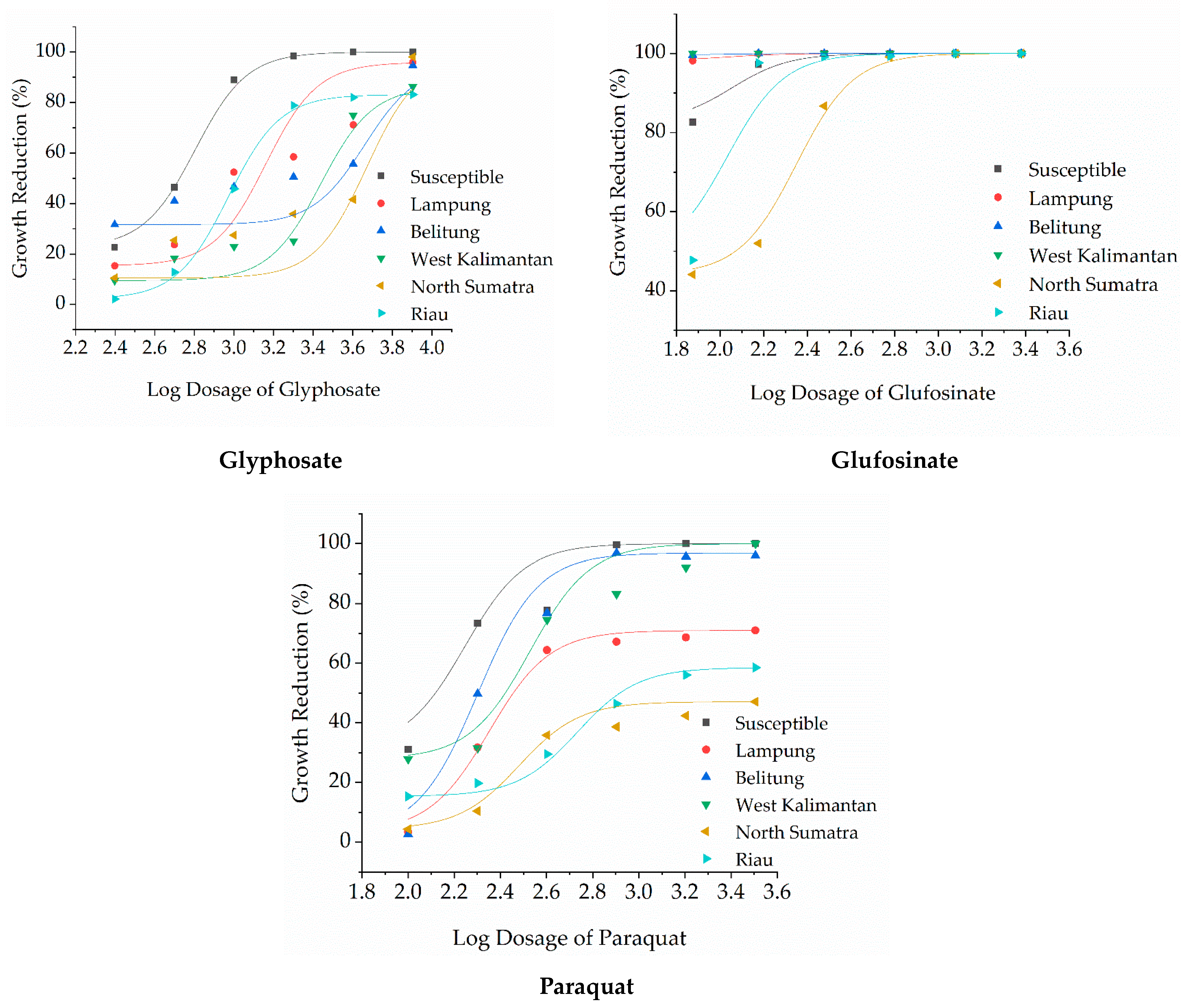
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dass, A.; Shekhawat, K.; Choudhary, A.K.; Sepat, S.; Rathore, S.S.; Mahajan, G.; Chauhan, B.S. Weed Management in Rice Using Crop Competition—A Review. Crop Prot. 2017, 95, 45–52. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Fundamentals of Weed Science, 5th ed.; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Clout, M.N.; Williams, P.A. Invasive Species Management: A Handbook of Techniques; Sutherland, W.J., Ed.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds; Distribution and Biology; Krieger Publishing Company: Malabar, FL, USA, 1991. [Google Scholar]
- Osuna, M.D.; Goulart, I.C.G.R.; Vidal, R.A.; Ruiz Santaella, J.P.; De Prado, R. Resistance to ACCase Inhibitors in Eleusine indica from Brazil Involves A Target Site Mutation. Planta Daninha 2012, 30, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Deng, W.; Li, X.; Yu, Q.; Bai, L.; Zheng, M. Target-Site and Non-Target-Site Based Resistance to the Herbicide Tribenuron-Methyl in Flixweed (Descurainia sophia L.). BMC Genom. 2016, 17, 551. [Google Scholar] [CrossRef] [Green Version]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef] [Green Version]
- Devi, E.L.; Kumar, S.; Basanta Singh, T.; Sharma, S.K.; Beemrote, A.; Devi, C.P.; Chongtham, S.K.; Singh, C.H.; Yumlembam, R.A.; Haribhushan, A.; et al. Adaptation Strategies and Defence Mechanisms of Plants during Environmental Stress. In Medicinal Plants and Environmental Challenges; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 359–413. [Google Scholar]
- Powles, S.B.; Preston, C. Herbicide Cross Resistance and Multiple Resistance in Plants; Herbicide Resistance Action Committee: Adelaide, South Australia, 1995; Volume 2, pp. 1–13. [Google Scholar]
- Vrbničanin, S.; Pavlović, D.; Božić, D. Weed Resistance to Herbicides. In Herbicide Resistance in Weeds and Crop; Pacanoski, Z., Ed.; IntechOpen: London, UK, 2017; pp. 7–35. [Google Scholar]
- Duke, S.O. The History and Current Status of Glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Gherekhloo, J.; Fernández-Moreno, P.T.; Alcántara-De La Cruz, R.; Sánchez-González, E.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Pro-106-Ser Mutation and EPSPS Overexpression Acting Together Simultaneously in Glyphosate-Resistant Goosegrass (Eleusine indica). Sci. Rep. 2017, 7, 6702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, H.K.; Mendes, R.R.; Scoz, L.B.; Lopez Ovejero, R.F.; Constantin, J.; Gaines, T.A.; Westra, P.; Dayan, F.E.; Oliveira, R.S. Proline-106 EPSPS Mutation Imparting Glyphosate Resistance in Goosegrass (Eleusine indica) Emerges in South America. Weed Sci. 2019, 67, 48–56. [Google Scholar] [CrossRef]
- Petterson, D.E.; Thompson, C.R.; Shoup, D.E.; Jugulam, M. Herbicide Mode of Action; K-State Research and Extension; Kansas State University: Manhattan, KS, USA, 2015. [Google Scholar]
- Heap, I. The International Herbicide-Resistant Weed Database 2023. Available online: weedscience.org (accessed on 29 May 2023).
- Tampubolon, K.; Purba, E.; Basyuni, M.; Hanafiah, D.S. Glyphosate Resistance of Eleusine indica Populations from North Sumatra, Indonesia. Biodiversitas 2019, 20, 1910–1916. [Google Scholar] [CrossRef]
- Istriningsih; Dewi, Y.A.; Yulianti, A.; Hanifah, V.W.; Jamal, E.; Dadang; Sarwani, M.; Mardiharini, M.; Anugrah, I.S.; Darwis, V.; et al. Farmers’ Knowledge and Practice Regarding Good Agricultural Practices (GAP) on Safe Pesticide Usage in Indonesia. Heliyon 2022, 8, e08708. [Google Scholar] [CrossRef]
- Directorate of Fertilizers and Pesticides Sistem Informasi Pestisida. Available online: http://pestisida.id/simpes_app/ (accessed on 29 May 2023).
- Burgos, N.R. Whole-Plant and Seed Bioassays for Resistance Confirmation. Weed Sci. 2015, 63, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Seefeldt, S.S.; Jensen, J.E.; Feurst, E.P. Log-Logistic Analysis of Herbicide Dose-Response Relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- Widayat, D.; Sumekar, Y. The Effect of Butyl Cyhalophop Herbicide on Weeds, Growth and Yield of Rice Plants (Oryza sativa L.). Int. J. Vet. Sci. Agric. Res. 2019, 1, 2582–4112. [Google Scholar]
- Ahmad-Hamdani, M.S.; Owen, M.J.; Yu, Q.; Powles, S.B. ACCase-Inhibiting Herbicide-Resistant Avena spp. Populations from the Western Australian Grain Belt. Weed Technol. 2012, 26, 130–136. [Google Scholar] [CrossRef]
- Loddo, D.; Imperatore, G.; Milani, A.; Panozzo, S.; Farinati, S.; Sattin, M.; Zanin, G. Article First Report of Glyphosate-Resistant Biotype of Eleusine indica (L.) Gaertn. in Europe. Agronomy 2020, 10, 1692. [Google Scholar] [CrossRef]
- Yu, Q.; Jalaludin, A.; Han, H.; Chen, M.; Douglas Sammons, R.; Powles, S.B. Evolution of a Double Amino Acid Substitution in the 5-Enolpyruvylshikimate-3-Phosphate Synthase in Eleusine indica Conferring High-Level Glyphosate Resistance. Plant Physiol. 2015, 167, 1440–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Huang, H.; Zhang, C.; Wei, S.; Huang, Z.; Chen, J.; Wang, X. Mutations and Amplification of EPSPS Gene Confer Resistance to Glyphosate in Goosegrass (Eleusine indica). Planta 2015, 242, 859–868. [Google Scholar] [CrossRef]
- Page, E.R.; Grainger, C.M.; Laforest, M.; Nurse, R.E.; Rajcan, I.; Bae, J.; Tardif, F.J. Target and Non-Target Site Mechanisms Confer Resistance to Glyphosate in Canadian Accessions of Conyza canadensis. Weed Sci. 2018, 66, 234–245. [Google Scholar] [CrossRef]
- Li, J.; Peng, Q.; Han, H.; Nyporko, A.; Kulynych, T.; Yu, Q.; Powles, S. Glyphosate Resistance in Tridax procumbens via a Novel EPSPS Thr-102-Ser Substitution. J. Agric. Food Chem. 2018, 66, 7880–7888. [Google Scholar] [CrossRef]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Alvarez, C.E.; Tuesca, D.; Permingeat, H.R. A Novel Triple Amino Acid Substitution in the EPSPS Found in a High-Level Glyphosate-Resistant Amaranthus hybridus Population from Argentina. Pest Manag. Sci. 2019, 75, 1242–1251. [Google Scholar] [CrossRef]
- García, M.J.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Bracamonte, E.; Portugal, J.; Alcántara-De la Cruz, R.; De Prado, R. The Triple Amino Acid Substitution TAP-IVS in the EPSPS Gene Confers High Glyphosate Resistance to the Superweed Amaranthus hybridus. Int. J. Mol. Sci. 2019, 20, 2396. [Google Scholar] [CrossRef] [Green Version]
- Vargas, L.; Ulguim, A.R.; Agostinetto, D.; Margo, T.D.; Thurmer, L. Low Level Resistance of Goosegrass (Eleusine indica) to Glyphosate in Rio Grande Do Sul Brazil. Planta Daninha 2013, 31, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Jalaludin, A.; Yu, Q.; Powles, S.B. Multiple Resistance across Glufosinate, Glyphosate, Paraquat and ACCase-Inhibiting Herbicides in an Eleusine indica Population. Weed Res. 2015, 55, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Beckie, H.J.; Reboud, X. Selecting for Weed Resistance: Herbicide Rotation and Mixture. Weed Technol. 2009, 23, 363–370. [Google Scholar] [CrossRef]
- Diggle, A.J.; Neve, P.B.; Smithà, F.P. Herbicides Used in Combination Can Reduce The Probability of Herbicide Resistance in Finite Weed Populations. Weed Res. 2003, 43, 371–378. [Google Scholar] [CrossRef]
- Owen, M.D.K. Diverse Approaches to Herbicide-Resistant Weed Management. Weed Sci. 2016, 64, 570–584. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Province | District | Coordinate | Eleusine indica |
|---|---|---|---|
| North Sumatra | Langkat | 3°38′49.0″ N 98°19′44.3″ E | Susceptible |
| Lampung | Pesawaran | 5°10′07.0″ S 105°06′12.0″ E | Resistant |
| Belitung | East Belitung | 3°1′23″ S 107°52′23″ E | Resistant |
| West Kalimantan | Ketapang | 1°28′28.1″ S 110°12′23.4″ E | Resistant |
| North Sumatra | Langkat | 3°38′49.046″ N 98°19′34.458″ E | Resistant |
| Riau | Pelalawan | 0°9′22.439″ S 102°12′51.997″ E | Resistant |
| Herbicide | Biotype of Eleusine indica | b2 | r2 | GR50 (g a.i. ha−1) | Resistance Index | Level of Resistance [22] |
|---|---|---|---|---|---|---|
| Glyphosate | Susceptible | 2.34 | 0.98 | 483.4 | - | S |
| Lampung | 1.07 | 0.95 | 1240.0 | 2.6 | L | |
| Belitung | 0.62 | 0.76 | 1163.4 | 2.4 | L | |
| West Kalimantan | 1.48 | 0.90 | 2573.3 | 5.3 | L | |
| North Sumatra | 1.06 | 0.78 | 2711.0 | 5.6 | L | |
| Riau | 1.75 | 0.94 | 1178.1 | 2.4 | L | |
| Paraquat | Susceptible | 1.86 | 0.95 | 141.0 | - | S |
| Lampung | 0.86 | 0.77 | 469.6 | 3.3 | L | |
| Belitung | 2.60 | 0.97 | 222.7 | 1.6 | S | |
| West Kalimantan | 1.47 | 0.95 | 247.7 | 1.8 | S | |
| North Sumatra | 0.59 | 0.80 | 2679.2 | 19.0 | H | |
| Riau | 0.65 | 0.95 | 1357.7 | 9.6 | M | |
| Glufosinate | Susceptible | 2.94 | 0.99 | 44.1 | - | S |
| Lampung | - | - | - | - | S | |
| Belitung | - | - | - | - | S | |
| West Kalimantan | - | - | - | - | S | |
| North Sumatra | 1.50 | 0.93 | 105.4 | 2.4 | L | |
| Riau | 5.48 | 0.99 | 76.3 | 1.7 | S | |
| Propaquizafop | Susceptible | - | - | - | - | S |
| Lampung | - | - | - | - | S | |
| Belitung | - | - | - | - | S | |
| West Kalimantan | - | - | - | - | S | |
| North Sumatra | - | - | - | - | S | |
| Riau | - | - | - | - | S | |
| Ametryn | Susceptible | - | - | - | - | S |
| Lampung | - | - | - | - | S | |
| Belitung | - | - | - | - | S | |
| West Kalimantan | - | - | - | - | S | |
| North Sumatra | - | - | - | - | S | |
| Riau | - | - | - | - | S | |
| Sulfentrazone | Susceptible | - | - | - | - | S |
| Lampung | - | - | - | - | S | |
| Belitung | - | - | - | - | S | |
| West Kalimantan | - | - | - | - | S | |
| North Sumatra | - | - | - | - | S | |
| Riau | - | - | - | - | S |
| Susceptible Biotype Reference [13], Similar to GenBank Accession AY157642 | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ACT Threonine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 CCA Proline |
|---|---|---|---|---|---|---|---|
| Susceptible | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ACT Threonine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 CCA Proline |
| Lampung | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ATT Isoleucine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 TCA Serine |
| Belitung | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ATT Isoleucine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 TCA Serine |
| West Kalimantan | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ATT Isoleucine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 TCA Serine |
| North Sumatra | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ATT Isoleucine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 TCA Serine |
| Riau | Position: Sequence: Amino Acid: | 101 GGA Glycine | 102 ATT Isoleucine | 103 GCA Alanine | 104 ATG Methionine | 105 CGA Arginine | 106 TCA Serine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurniadie, D.; Widianto, R.; Umiyati, U.; Widayat, D.; Nasahi, C.; Budiawan, A. Management of Eleusine indica (L.) Gaertn Resistance to Glyphosate Herbicide in Indonesia. Agronomy 2023, 13, 1649. https://doi.org/10.3390/agronomy13061649
Kurniadie D, Widianto R, Umiyati U, Widayat D, Nasahi C, Budiawan A. Management of Eleusine indica (L.) Gaertn Resistance to Glyphosate Herbicide in Indonesia. Agronomy. 2023; 13(6):1649. https://doi.org/10.3390/agronomy13061649
Chicago/Turabian StyleKurniadie, Denny, Ryan Widianto, Uum Umiyati, Dedi Widayat, Ceppy Nasahi, and Ari Budiawan. 2023. "Management of Eleusine indica (L.) Gaertn Resistance to Glyphosate Herbicide in Indonesia" Agronomy 13, no. 6: 1649. https://doi.org/10.3390/agronomy13061649