


Physiological and Histological Characterization of the ESB1 TILLING Mutant of Brassica rapa L.: Potential Use in Biofortification and Phytoremediation Programs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions, and Experimental Design
2.2. Plant Sampling
2.3. Determination of Photosynthetic Pigments Concentration
2.4. Fluorescence Parameters Analysis
2.5. Determination of Leaf Gas Exchange Parameters
2.6. Histological Staining of Root Suberin
2.7. Mineral Elements Concentration
2.8. Statistical Analysis
3. Results and Discussion
3.1. Plant Biomass and Root Morphology
3.2. Photosynthetic Pigments Concentration
3.3. Chl a Fluorescence
3.4. Leaf Gas Exchange Parameters
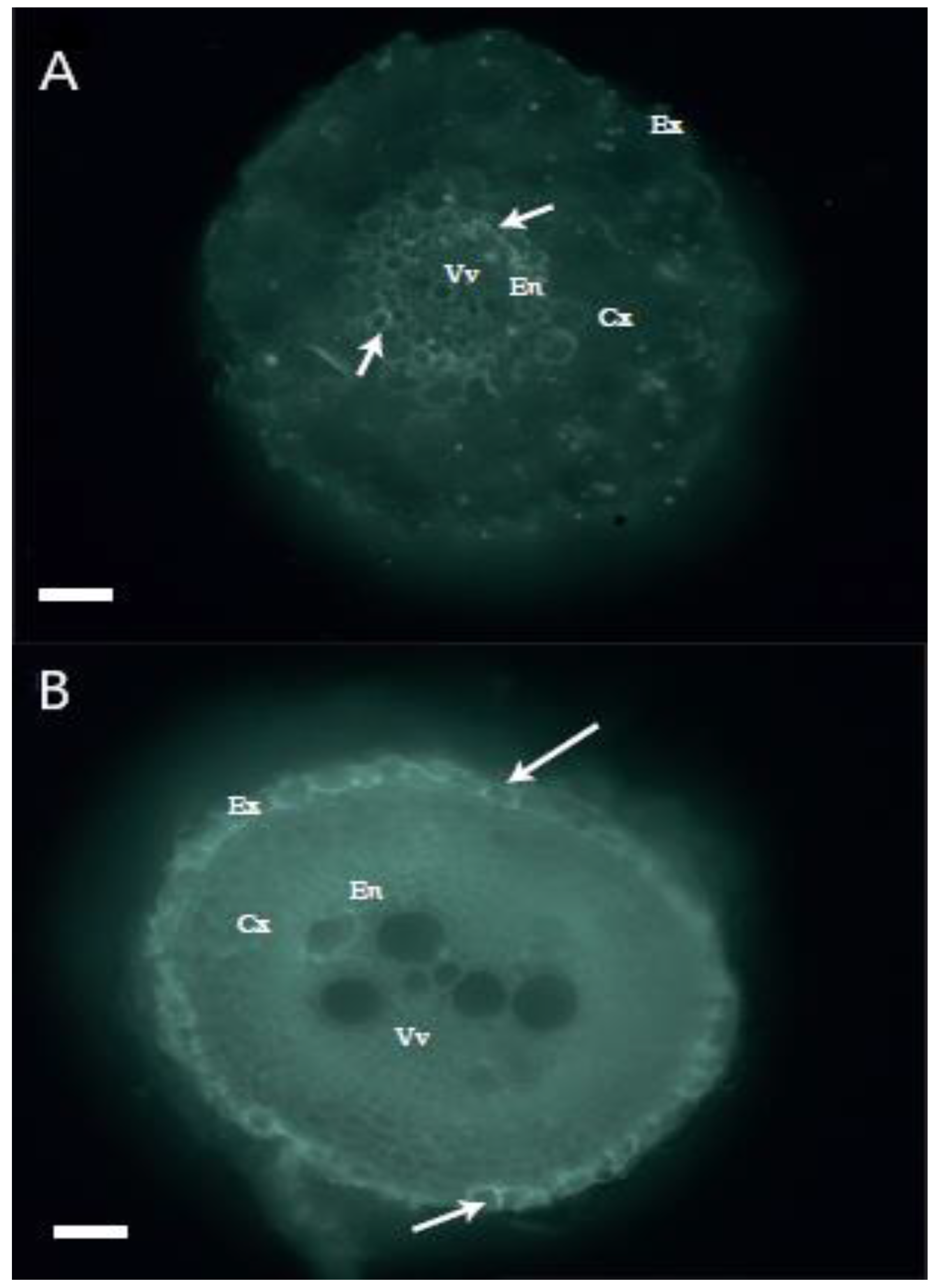
3.5. Histological Study of Root Suberin
3.6. Essential Mineral Nutrients
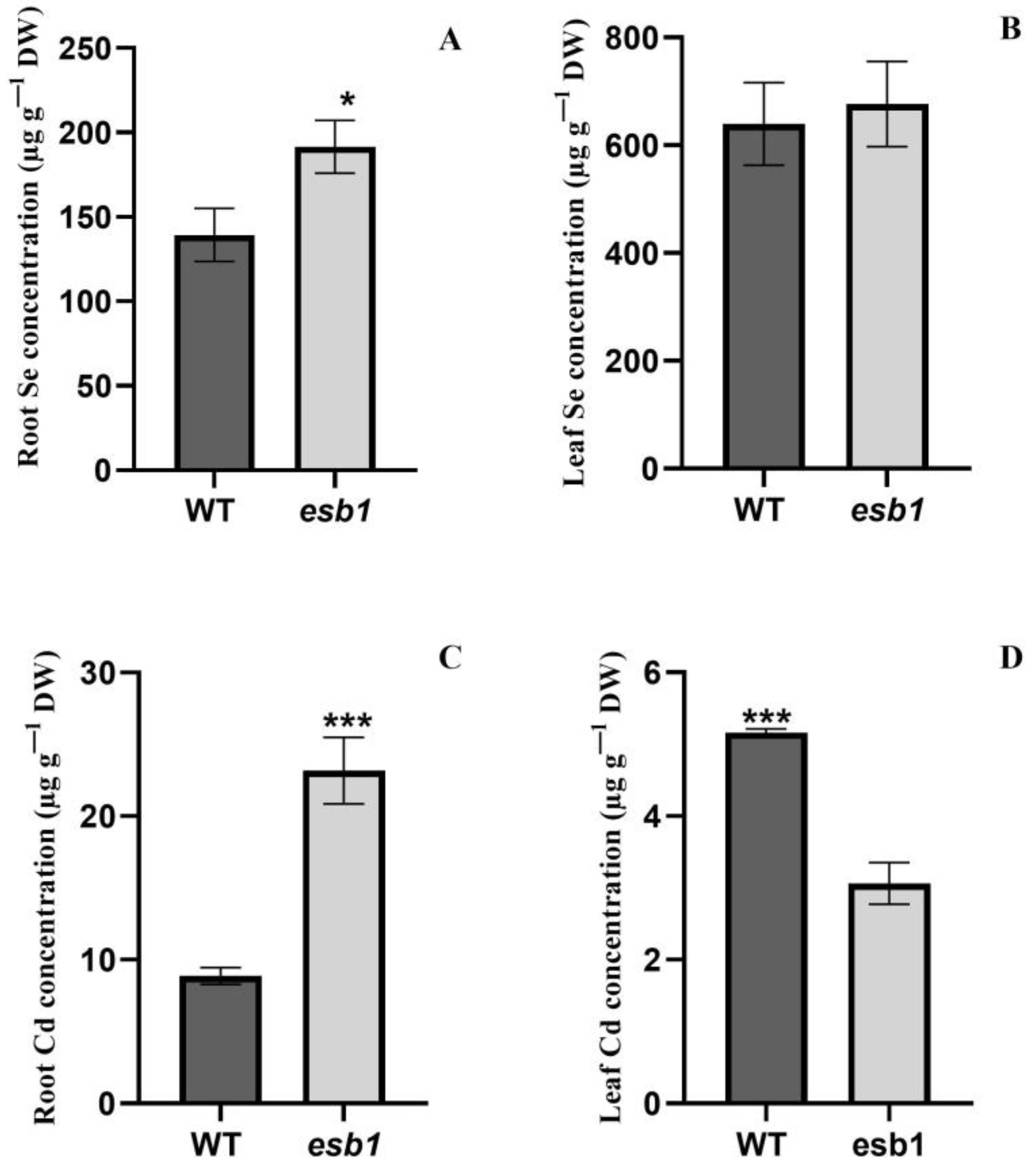
3.7. Se and Cd Concentration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramakrishna, P.; Barberon, M. Polarized Transport across Root Epithelia. Curr. Opin. Plant. Biol. 2019, 52, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Bachelier, J.B.; Razik, I.; Schauer, M.; Seago, J.L. Roots Structure and Development of Austrobaileya Scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms. Plants 2020, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Tam, N.F.Y.; Li, W.C.; Ye, Z. The Role of Root Apoplastic Barriers in Cadmium Translocation and Accumulation in Cultivars of Rice (Oryza Sativa L.) with Different Cd-Accumulating Characteristics. Environ. Pollut. 2020, 264, 114736. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in Plants: Uptake, Toxicity, and Its Interactions with Selenium Fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Chen, A.; Husted, S.; Salt, D.E.; Schjoerring, J.K.; Persson, D.P. The Intensity of Manganese Deficiency Strongly Affects Root Endodermal Suberization and Ion Homeostasis. Plant. Physiol. 2019, 181, 729–742. [Google Scholar] [CrossRef] [Green Version]
- Doblas, V.G.; Geldner, N.; Barberon, M. The Endodermis, a Tightly Controlled Barrier for Nutrients. Curr. Opin. Plant. Biol. 2017, 39, 136–143. [Google Scholar] [CrossRef]
- Barberon, M. The Endodermis as a Checkpoint for Nutrients. New Phytol. 2017, 213, 1604–1610. [Google Scholar] [CrossRef]
- Barbosa, I.C.R.; Rojas-Murcia, N.; Geldner, N. The Casparian Strip—One Ring to Bring Cell Biology to Lignification? Curr. Opin. Biotechnol. 2019, 56, 121–129. [Google Scholar] [CrossRef]
- Wang, P.; Calvo-Polanco, M.; Reyt, G.; Barberon, M.; Champeyroux, C.; Santoni, V.; Maurel, C.; Franke, R.B.; Ljung, K.; Novak, O.; et al. Surveillance of Cell Wall Diffusion Barrier Integrity Modulates Water and Solute Transport in Plants. Sci. Rep. 2019, 9, 4227. [Google Scholar] [CrossRef] [Green Version]
- Namyslov, J.; Bauriedlová, Z.; Janoušková, J.; Soukup, A.; Tylová, E. Exodermis and Endodermis Respond to Nutrient Deficiency in Nutrient-Specific and Localized Manner. Plants 2020, 9, 201. [Google Scholar] [CrossRef] [Green Version]
- Baxter, I.; Hosmani, P.S.; Rus, A.; Lahner, B.; Borevitz, J.O.; Muthukumar, B.; Mickelbart, M.V.; Schreiber, L.; Franke, R.B.; Salt, D.E. Root Suberin Forms an Extracellular Barrier That Affects Water Relations and Mineral Nutrition in Arabidopsis. PLoS Genet. 2009, 5, e1000492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Yeh, K.C. The Dual Benefit of a Dominant Mutation in Arabidopsis IRON DEFICIENCY TOLERANT1 for Iron Biofortification and Heavy Metal Phytoremediation. Plant. Biotechnol. J. 2020, 18, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Straeten, D.; Bhullar, N.K.; De Steur, H.; Gruissem, W.; MacKenzie, D.; Pfeiffer, W.; Qaim, M.; Slamet-Loedin, I.; Strobbe, S.; Tohme, J.; et al. Multiplying the Efficiency and Impact of Biofortification through Metabolic Engineering. Nat. Commun. 2020, 11, 5203. [Google Scholar] [CrossRef] [PubMed]
- Navarro-León, E.; Javier López-Moreno, F.; Ahmad, A.; Ruiz, J.M.; Blasco, B.; Navarro-León, E.; Ahmad, A.; Ruiz, J.M.; Blasco, B.; López-Moreno, F.J. Biofortification of Crops: Novel Insights and Approaches for Enhanced Nutrient Accumulation. In Conceptualizing Plant-Based Nutrition; Springer: Singapore, 2022; pp. 19–41. [Google Scholar] [CrossRef]
- Sarwar, N.; Akhtar, M.; Kamran, M.A.; Imran, M.; Riaz, M.A.; Kamran, K.; Hussain, S. Selenium Biofortification in Food Crops: Key Mechanisms and Future Perspectives. J. Food Compos. Anal. 2020, 93, 103615. [Google Scholar] [CrossRef]
- Schiavon, M.; Nardi, S.; dalla Vecchia, F.; Ertani, A. Selenium Biofortification in the 21st Century: Status and Challenges for Healthy Human Nutrition. Plant. Soil. 2020, 453, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Izydorczyk, G.; Ligas, B.; Mikula, K.; Witek-Krowiak, A.; Moustakas, K.; Chojnacka, K. Biofortification of Edible Plants with Selenium and Iodine—A Systematic Literature Review. Sci. Total. Environ. 2021, 754, 141983. [Google Scholar] [CrossRef]
- Jaskulak, M.; Grobelak, A.; Vandenbulcke, F. Modelling Assisted Phytoremediation of Soils Contaminated with Heavy Metals–Main Opportunities, Limitations, Decision Making and Future Prospects. Chemosphere 2020, 249, 126196. [Google Scholar] [CrossRef]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public. Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Z.; Song, W.; Hong, D.; Huang, L.; Li, Y. A Review on Cadmium Exposure in the Population and Intervention Strategies Against Cadmium Toxicity. Bull. Env. Contam. Toxicol. 2021, 106, 65–74. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zia ur Rehman, M.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium Phytoremediation Potential of Brassica Crop Species: A Review. Sci. Total. Environ. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef]
- Li, X.; Wu, Y.; Li, B.; Yang, Y.; Yang, Y. Selenium Accumulation Characteristics and Biofortification Potentiality in Turnip (Brassica rapa var. rapa) Supplied with Selenite or Selenate. Front. Plant. Sci. 2018, 8, 2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-León, E.; Oviedo-Silva, J.; Ruiz, J.M.; Blasco, B. Possible role of HMA4a TILLING mutants of Brassica rapa in cadmium phytoremediation programs. Ecotoxicol. Environ. Saf. 2019, 180, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Lochlainn, S.T.; Amoah, S.; Graham, N.S.; Alamer, K.; Rios, J.J.; Kurup, S.; Stoute, A.; Hammond, J.P.; Østergaard, L.; King, G.J.; et al. High Resolution Melt (HRM) Analysis Is an Efficient Tool to Genotype EMS Mutants in Complex Crop Genomes. Plant. Methods 2011, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, N.S.; Hammond, J.P.; Lysenko, A.; Mayes, S.; Lochlainn, S.Ó.; Blasco, B.; Bowen, H.C.; Rawlings, C.J.; Rios, J.J.; Welham, S.; et al. Genetical and Comparative Genomics of Brassica under Altered CA Supply Identifies Arabidopsis CA-Transporter Orthologs. Plant. Cell 2014, 26, 2818–2830. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant. Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Vaculík, M.; Konlechner, C.; Langer, I.; Adlassnig, W.; Puschenreiter, M.; Lux, A.; Hauser, M.T. Root Anatomy and Element Distribution Vary between Two Salix Caprea Isolates with Different Cd Accumulation Capacities. Environ. Pollut. 2012, 163, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Wolf, B. A Comprehensive System of Leaf Analyses and Its Use for Diagnosing Crop Nutrient Status. Commun. Soil. Sci. Plant. Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: A study of a modified Berthelot reaction using salicylate and dichloroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Shikha, D.; Singh, P.K. In Situ Phytoremediation of Heavy Metal–Contaminated Soil and Groundwater: A Green Inventive Approach. Environ. Sci. Pollut. Res. 2021, 28, 4104–4124. [Google Scholar] [CrossRef]
- Ranathunge, K.; Schreiber, L. Water and Solute Permeabilities of Arabidopsis Roots in Relation to the Amount and Composition of Aliphatic Suberin. J. Exp. Bot. 2011, 62, 1961–1974. [Google Scholar] [CrossRef]
- Sharkey, T.D. Emerging Research in Plant Photosynthesis. Emerg. Top. Life Sci. 2020, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant. Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.M. Continuous Excitation Chlorophyll Fluorescence Parameters: A Review for Practitioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef]
- Kim, D.; Kwak, J.I.; An, Y.J. Physiological Response of Crop Plants to the Endocrine-Disrupting Chemical Nonylphenol in the Soil Environment. Environ. Pollut. 2019, 251, 573–580. [Google Scholar] [CrossRef]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of Stomatal Density and Morphology on Water-Use Efficiency in a Changing World. Front. Plant. Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- Kreszies, T.; Kreszies, V.; Ly, F.; Thangamani, P.D.; Shellakkutti, N.; Schreiber, L. Suberized Transport Barriers in Plant Roots: The Effect of Silicon. J. Exp. Bot. 2020, 71, 6799–6806. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Masood, S.; Ullah, N.; Abbasi, E.; Hussain, Z.; Ali, I. Molecular Basis of Iron Biofortification in Crop Plants; A Step towards Sustainability. Plant Breed. 2021, 140, 12–22. [Google Scholar] [CrossRef]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral Biofortification of Vegetables as a Tool to Improve Human Diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Wu, J.; Mock, H.P.; Giehl, R.F.H.; Pitann, B.; Mühling, K.H. Silicon Decreases Cadmium Concentrations by Modulating Root Endodermal Suberin Development in Wheat Plants. J. Hazard. Mater. 2019, 364, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; Al-Duaij, O.K. Phytoremediation of Heavy Metals: Mechanisms, Methods and Enhancements. Env. Chem. Lett. 2018, 16, 1339–1359. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Leaf DW (g plant−1) | Root DW (g plant−1) | Root Length (cm) | Root Volume (cm3) | |
|---|---|---|---|---|
| WT | 1.34 ± 0.17 | 0.11 ± 0.01 | 32.94 ± 1.06 | 1.81 ± 0.17 |
| esb1 | 1.46 ± 0.17 | 0.10 ± 0.02 | 39.70 ± 2.13 | 2.14 ± 0.31 |
| p-value | NS | NS | * | NS |
| LSD0.05 | 0.54 | 0.04 | 4.99 | 0.74 |
| Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Carotenoids (µg g−1 FW) | Ratio of Chl a/b | |
|---|---|---|---|---|
| WT | 0.367 ± 0.003 | 0.190 ± 0.005 | 45.42 ± 1.36 | 1.94 ± 0.05 |
| esb1 | 0.384 ± 0.004 | 0.184 ± 0.002 | 48.59 ± 1.63 | 2.13 ± 0.01 |
| p-value | ** | NS | NS | ** |
| LSD0.05 | 0.01 | 0.01 | 4.51 | 0.12 |
| Fv/Fm | RC/ABS | ΦEo | Ψo | PIABS | |
|---|---|---|---|---|---|
| WT | 0.839 ± 0.001 | 0.76 ± 0.01 | 0.50 ± 0.01 | 0.60 ± 0.03 | 6.17 ± 0.60 |
| esb1 | 0.844 ± 0.001 | 0.74 ± 0.02 | 0.51 ± 0.01 | 0.61 ± 0.03 | 5.64 ± 0.77 |
| p-value | ** | NS | NS | NS | NS |
| LSD0.05 | 0.003 | 0.059 | 0.028 | 0.032 | 0.692 |
| A (µmol m−2s−1) | gs (mol m−2s−1) | Ci (µmol mol−1) | E (mmol m−2s−1) | WUE | |
|---|---|---|---|---|---|
| WT | 29.03 ± 0.50 | 0.38 ± 0.02 | 309.00 ± 7.90 | 4.52 ± 0.15 | 6.43 ± 0.11 |
| esb1 | 30.69 ± 0.74 | 0.43 ± 0.02 | 312.82 ± 8.91 | 5.44 ± 0.21 | 5.49 ± 0.18 |
| p-value | NS | NS | NS | * | ** |
| LSD0.05 | 1.987 | 0.072 | 69.873 | 0.576 | 0.467 |
| N | P | K | S | Ca | Mg | Fe | B | Cu | Mn | Zn | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | 30.40 ± 2.87 | 17.21 ± 1.23 | 31.22 ± 2.24 | 3.73 ± 0.26 | 40.12 ± 5.44 | 16.74 ± 2.39 | 131,333.42 ± 470.48 | 30.59 ± 4.06 | 46.67 ± 9.50 | 422.27 ± 8.22 | 45.11 ± 0.64 |
| esb1 | 56.93 ± 4.64 | 18.48 ± 0.38 | 29.62 ± 3.26 | 4.83 ± 0.32 | 49.79 ± 3.47 | 25.02 ± 1.30 | 167,769.11 ± 415.46 | 27.67 ± 3.30 | 57.86 ± 5.12 | 500.45 ± 17.85 | 56.71 ± 1.94 |
| p-value | *** | NS | NS | NS | NS | * | ** | NS | NS | * | ** |
| LSD0.05 | 11.57 | 3.59 | 10.99 | 1.16 | 17.92 | 7.54 | 1742.68 | 14.54 | 29.97 | 54.56 | 5.68 |
| N | P | K | S | Ca | Mg | Fe | B | Cu | Mn | Zn | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | 60.97 ± 3.79 | 7.32 ± 0.03 | 62.57 ± 4.77 | 34.97 ± 2.53 | 47.39 ± 3.82 | 9.68 ± 0.85 | 508.55 ± 35.64 | 236.87 ± 11.91 | 9.17 ± 0.36 | 221.77 ± 11.13 | 28.91 ± 3.45 |
| esb1 | 60.89 ± 2.52 | 7.75 ± 0.15 | 58.88 ± 2.37 | 35.50 ± 2.59 | 48.13 ± 2.82 | 10.14 ± 0.58 | 840.08 ± 7.85 | 300.68 ± 11.74 | 14.07 ± 2.03 | 234.41 ± 10.57 | 20.96 ± 0.44 |
| p-value | NS | * | NS | NS | NS | NS | *** | * | ** | NS | NS |
| LSD0.05 | 9.66 | 0.42 | 14.79 | 10.06 | 13.19 | 2.86 | 101.322 | 46.44 | 1.92 | 42.61 | 9.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atero-Calvo, S.; Rios, J.J.; Navarro-León, E.; Ruiz, J.M.; Blasco, B. Physiological and Histological Characterization of the ESB1 TILLING Mutant of Brassica rapa L.: Potential Use in Biofortification and Phytoremediation Programs. Agronomy 2023, 13, 1642. https://doi.org/10.3390/agronomy13061642
Atero-Calvo S, Rios JJ, Navarro-León E, Ruiz JM, Blasco B. Physiological and Histological Characterization of the ESB1 TILLING Mutant of Brassica rapa L.: Potential Use in Biofortification and Phytoremediation Programs. Agronomy. 2023; 13(6):1642. https://doi.org/10.3390/agronomy13061642
Chicago/Turabian StyleAtero-Calvo, Santiago, Juan José Rios, Eloy Navarro-León, Juan M. Ruiz, and Begoña Blasco. 2023. "Physiological and Histological Characterization of the ESB1 TILLING Mutant of Brassica rapa L.: Potential Use in Biofortification and Phytoremediation Programs" Agronomy 13, no. 6: 1642. https://doi.org/10.3390/agronomy13061642