
Spectral Response of Camelina (Camelina sativa (L.) Crantz) to Different Nitrogen Fertilization Regimes under Mediterranean Conditions

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup and Site Description
2.2. Crop Growth Cycle Monitoring
2.3. Non-Destructive Analysis and Destructive Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Indices | Formula |
|---|---|
| Normalized difference vegetation index (NDVI) [38] | NDVI = (NIR − RED)/(NIR + RED) |
| Ratio vegetation index (RVI) [39] | RVI = RED/NIR |
| Transformed normalized difference vegetation index [40] | TNDVI = (NIR − RED)/(NIR + RED) + 0.5]1/2 |
| Renormalize difference vegetation index (RDVI) [41] | RDVI = (NIR − RED)/(NIR + RED)1/2 |
| Improved modified chlorophyll absorption ratio index (MCARI2) [42] | MCARI 2 = 1.5 × [2.5 × (NIR − RED) − 1.3 × (NIR − GREEN)]/{[2NIR + 1]2 − [6NIR − 5 × (RED)1/2] − 0.5} |
| Modified red edge simple ratio index (MSR705) [43] | MSR 705 = (NIR/RED − 1)/(NIR/RED + 1)1/2 |
| Soil adjustment vegetation index (SAVI) [44] | SAVI = (NIR − RED) × (1 × L)/(NIR + RED + L); L = 0.5 |
2.4. Image Processing
2.5. Crop Production and Yield Component
2.6. Seed Quality
2.7. Statistical Analysis
3. Results
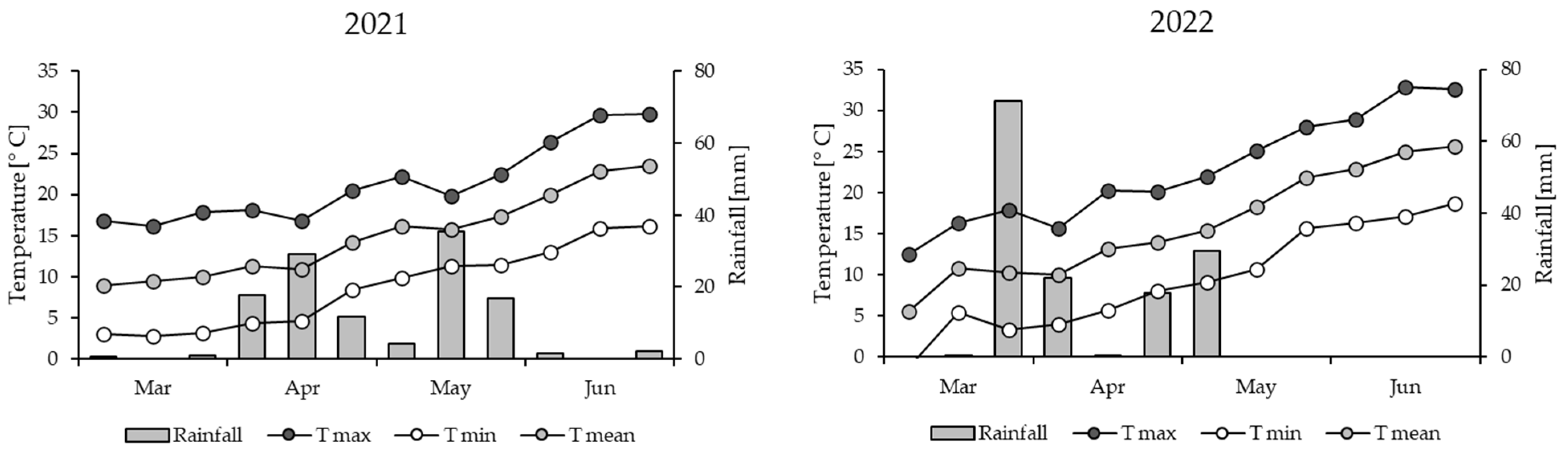
3.1. Meteorological Data and Crop Phenology
3.2. Vegetation Indices (VIs)
3.3. Dualex Detection and Destructive Samplings
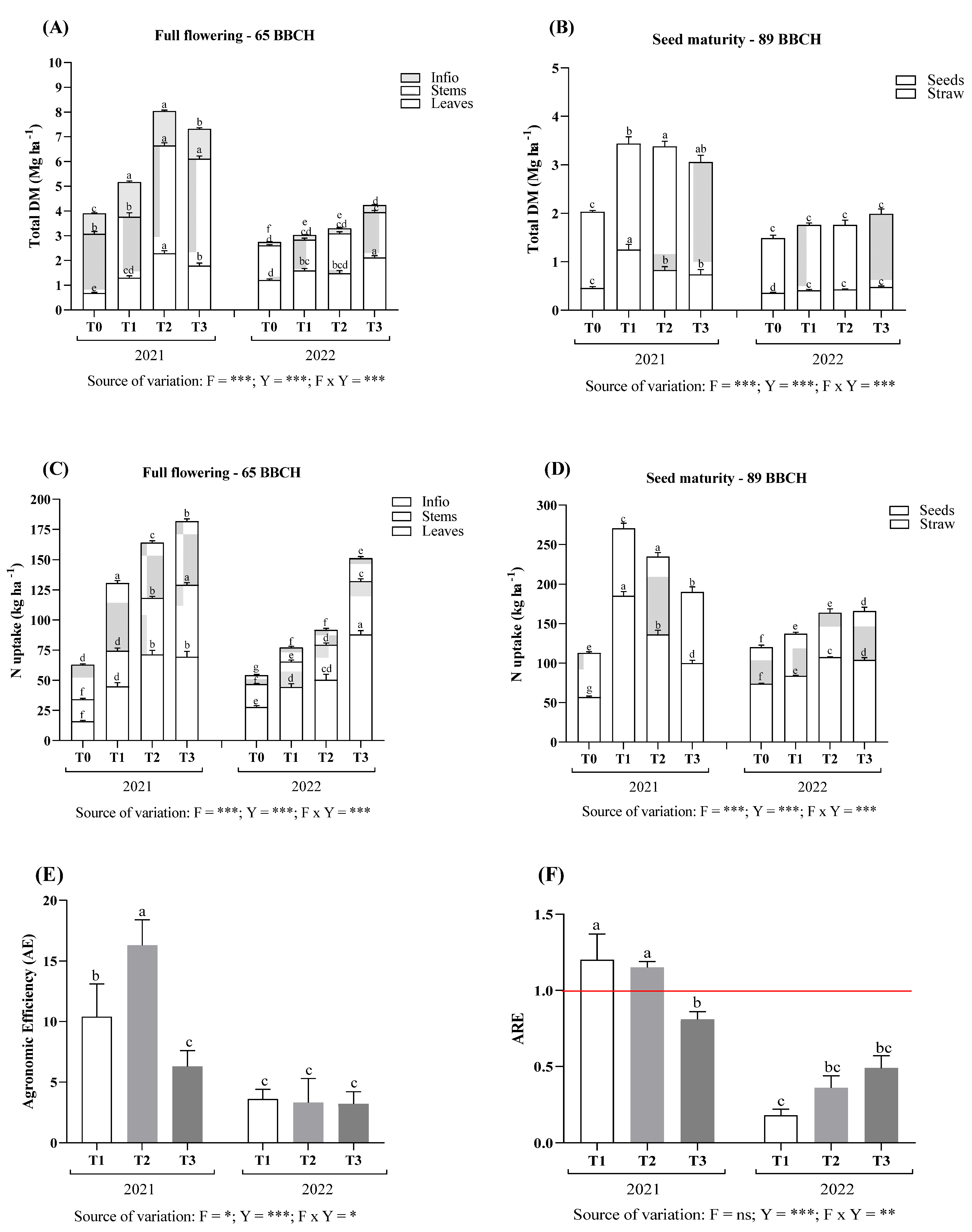
3.4. Dry Matter Accumulation and Nitrogen Uptake
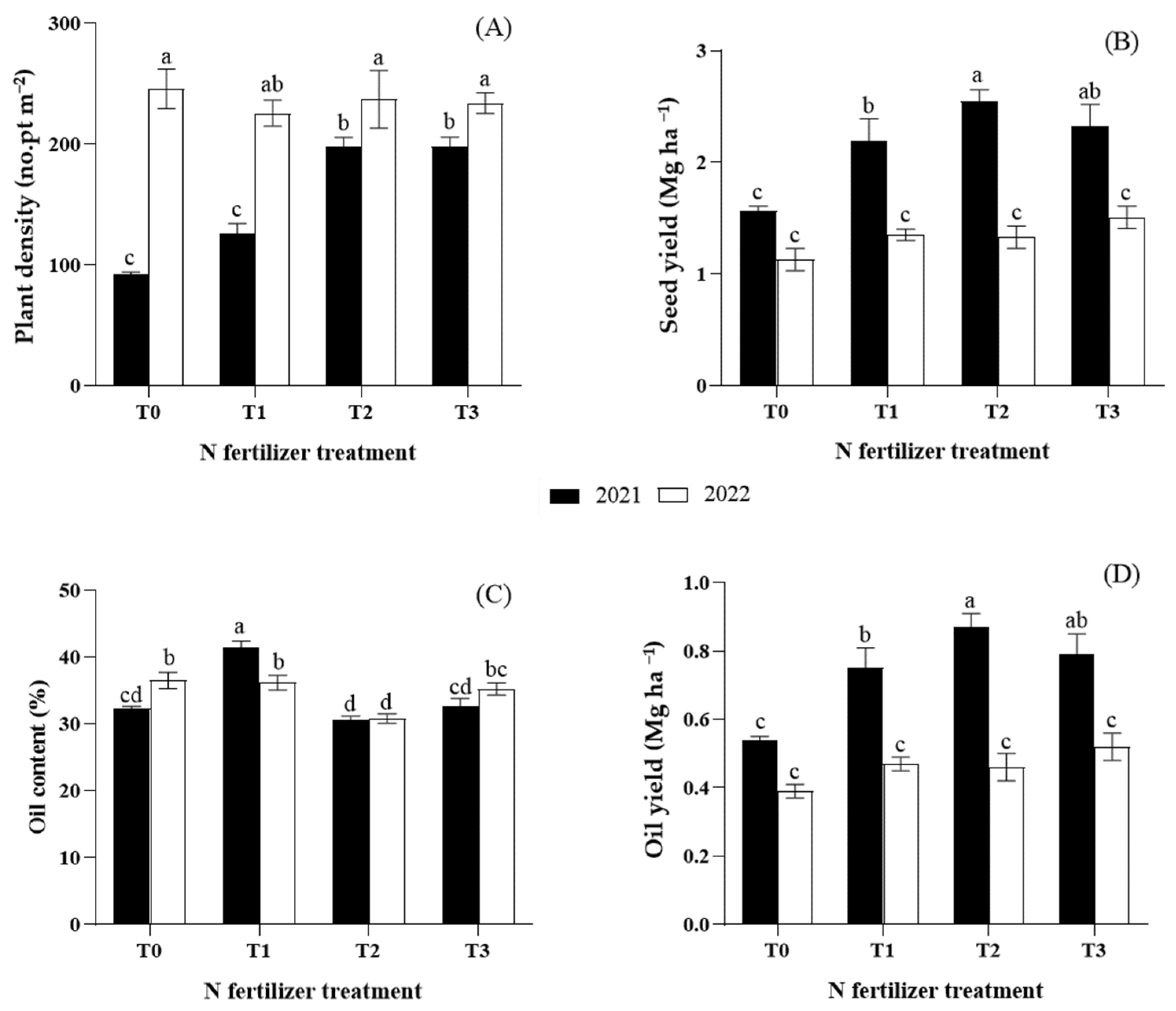
3.5. Yield, Yield Components, and Seed Quality
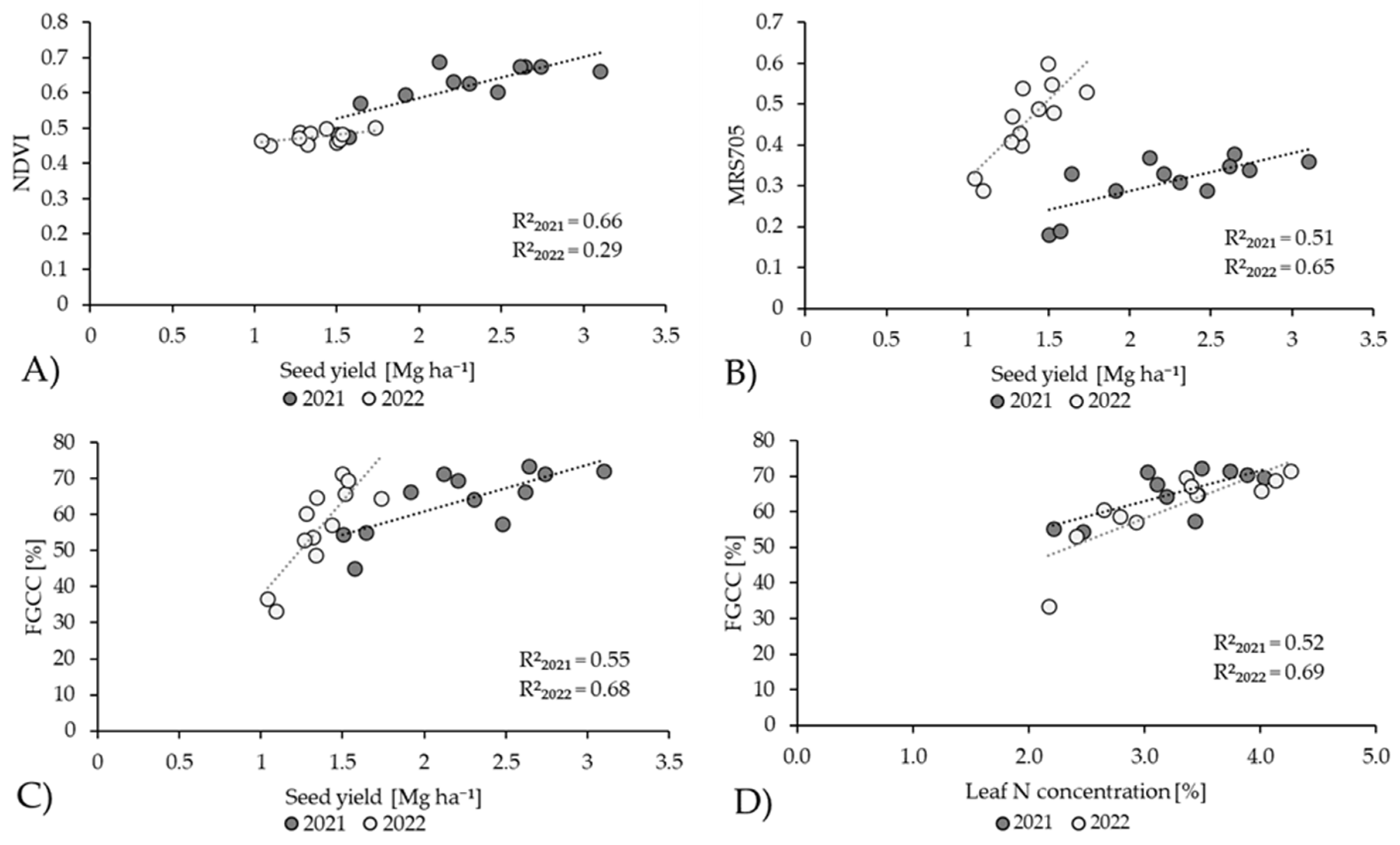
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khan, N.; Ray, R.L.; Sargani, G.R.; Ihtisham, M.; Khayyam, M.; Ismail, S. Current Progress and Future Prospects of Agriculture Technology: Gateway to Sustainable Agriculture. Sustainability 2021, 13, 4883. [Google Scholar] [CrossRef]
- Feng, W.; Guo, B.B.; Wang, Z.J.; He, L.; Song, X.; Wang, Y.H.; Guo, T.C. Measuring leaf nitrogen concentration in-winter wheat using double-peak spectral reflection remote sensing data. Field Crops Res. 2012, 159, 43–52. [Google Scholar] [CrossRef]
- Nasar, J.; Khan, W.; Khan, M.Z.; Gitari, H.I.; Gbolayori, H.F.; Moussa, A.A.; Mandozai, A.; Rizwan, N.; Anwari, G.; Maroof, S.M. Photosynthetic Activities and Photosynthetic Nitrogen Use Efficiency of Maize Crop Under Different Planting Patterns and Nitrogen Fertilization. J. Soil Sci. Plant Nutr. 2021, 21, 2274–2284. [Google Scholar] [CrossRef]
- Mahmud, K.; Panday, D.; Mergoum, A.; Missaoui, A. Nitrogen Losses and Potential Mitigation Strategies for a Sustainable Agroecosystem. Sustainability 2021, 13, 2400. [Google Scholar] [CrossRef]
- Li, Z.; Cui, S.; Zhang, Q.; Xu, G.; Feng, Q.; Chen, C.; Li, Y. Optimizing wheat yield, water, and nitrogen use efficiency with water and nitrogen inputs in China: A synthesis and life cycle assessment. Front. Plant Sci. 2022, 13, 930484. [Google Scholar] [CrossRef]
- He, L.; Zhang, H.Y.; Zhang, Y.S.; Song, X.; Feng, W.; Kang, G.Z.; Wang, C.Y.; Guo, T.C. Estimating canopy leaf nitrogen concentration in winter wheat based on multi-angular hyperspectral remote sensing. Eur. J. Agron. 2016, 73, 170–185. [Google Scholar] [CrossRef]
- Esteves, E.; Locatelli, G.; Bou, N.A.; Ferrarezi, R.S. Sap Analysis: A Powerful Tool for Monitoring Plant Nutrition. Horticulturae 2021, 7, 426. [Google Scholar] [CrossRef]
- Guo, B.B.; Qi, S.L.; Heng, Y.R.; Duan, J.Z.; Zhang, H.Y.; Wu, Y.P.; Feng, W.; Xie, Y.X.; Zhu, Y.J. Remotely assessing leaf N uptake in winter wheat based on canopy hyperspectral red-edge absorption. Eur. J. Agron. 2017, 82, 113–124. [Google Scholar] [CrossRef]
- Luisa España-Boquera, M.; Cárdenas-Navarro, R.; López-Pérez, L.; Castellanos-Morales, V.; Lobit, P. Estimating the nitrogen concentration of strawberry plants from its spectral response. Commun. Soil Sci. Plant Anal. 2006, 37, 2447–2459. [Google Scholar] [CrossRef]
- Sishodia, R.P.; Ray, R.L.; Singh, S.K. Applications of Remote Sensing in Precision Agriculture: A Review. Remote Sens. 2020, 12, 3136. [Google Scholar] [CrossRef]
- Weiss, M.; Jacob, F.; Duveiller, G. Remote sensing for agricultural applications: A meta-review. Remote Sens. Environ. 2020, 236, 111402. [Google Scholar] [CrossRef]
- Mezera, J.; Lukas, V.; Horniaček, I.; Smutný, V.; Elbl, J. Comparison of Proximal and Remote Sensing for the Diagnosis of Crop Status in Site-Specific Crop Management. Sensors 2022, 22, 19. [Google Scholar] [CrossRef]
- Berger, K.; Verrelst, J.; Féret, J.B.; Wang, Z.; Wocher, M.; Strathmann, M.; Danner, M.; Mauser, W.; Hank, T. Crop nitrogen monitoring: Recent progress and principal developments in the context of imaging spectroscopy missions. Remote Sens. Environ. 2020, 242, 111758. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, D.; Cai, Z.; Wang, L.; Wang, J.; Sun, L.; Fan, X.; Shen, S.; Zhao, J. Spectral technology and multispectral imaging for estimating the photosynthetic pigments and SPAD of the Chinese cabbage based on machine learning. Comput. Electron. Agric. 2022, 195, 106814. [Google Scholar] [CrossRef]
- Kamarianakis, Z.; Panagiotakis, S. Design and Implementation of a Low-Cost Chlorophyll Content Meter. Sensors 2023, 23, 2699. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berjon, A.; Lopez-Lozano, R.; Miller, J.R.; Martin, P.; Cachorro, V.; Gonzalez, M.R.; de Frutos, A. Assessing vineyard condition with hyperspectral indices: Leaf and canopy reflectance simulation in a row-structured discontinuous canopy. Remote Sens. Environ. 2005, 99, 271–287. [Google Scholar] [CrossRef]
- Fang, H.; Liang, S. Reference Module in Earth Systems and Environmental Sciences. In Encyclopedia of Ecology; Elsevier Reference Collection; Elsevier: Amsterdam, The Netherlands, 2014; pp. 2139–2148. ISBN 978-0-12-409548-9. [Google Scholar]
- Liu, S.; Li, L.; Fan, H.; Guo, X.; Wang, S.; Lu, J. Real-time and multi-stage recommendations for nitrogen fertilizer topdressing rates in winter oilseed rape based on canopy hyperspectral data. Ind. Crops Prod. 2020, 154, 112699. [Google Scholar] [CrossRef]
- Amiri, M.; Pourghasemi, H.R. Mapping the NDVI and monitoring of its changes using Google Earth Engine and Sentinel-2 images. In Computers in Earth and Environmental Sciences in: Artificial Intelligence and Advanced Technologies; Elsevier: Amsterdam, The Netherlands, 2022; pp. 127–136. [Google Scholar]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crop Prod. 2016, 94, 690–697. [Google Scholar] [CrossRef]
- Zanetti, F.; Alberghini, B.; Marjanović, A.; Jeromela, A.M.; Grahovac, N.; Rajković, D.; Kiprovski, B.; Monti, A. Camelina, an ancient oilseed crop actively contributing to the rural renaissance in Europe: A review. Agron. Sustain. Dev. 2021, 41, 2. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Stuper-Szablewska, K. The phytochemical quality of Camelina sativa seed and oil. Soil Plant Sci. 2019, 70, 39–47. [Google Scholar] [CrossRef]
- Oryschak, M.A.; Christianson, C.B.; Beltranena, E. Camelina sativa cake for broiler chickens: Effects of increasing dietary inclusion on clinical signs of toxicity, feed disappearance, and nutrient digestibility. Transl. Anim. Sci. 2020, 4, 1263–1277. [Google Scholar] [CrossRef]
- Juodka, R.; Nainienė, R.; Juškienė, V.; Juška, R.; Leikus, R.; Kadžienė, G.; Stankevičienė, D. Camelina (Camelina sativa (L.) Crantz) as Feedstuffs in Meat Type Poultry Diet: A Source of Protein and n-3 Fatty Acids. Animals 2022, 12, 295. [Google Scholar] [CrossRef]
- Cullere, M.; Singh, Y.; Pellattiero, E.; Berzuini, S.; Galasso, I.; Clemente, C.; Dalle Zotte, A. Effect of the dietary inclusion of Camelina sativa cake into quail diet on live performance, carcass traits and meat quality. Poult. Sci. 2023, 102, 102650. [Google Scholar] [CrossRef]
- Angelini, L.G.; Abou Chehade, L.; Foschi, L.; Tavarini, S. Performance and potentiality of camelina (Camelina sativa L. Crantz) genotypes in response to sowing date under Mediterranean Environment. Agronomy 2020, 10, 1929. [Google Scholar] [CrossRef]
- Vollmann, L.J.; Moritz, T.; Kargl, C.; Baumgartner, S.; Wagentristl, H. Agronomic evaluation of camelina genotypes selected for seed quality characteristics. Ind. Crop Prod. 2007, 26, 270–277. [Google Scholar] [CrossRef]
- Jankowsky, K.; Sokolski, M. Spring camelina: Effect of mineral fertilization on the energy efficiency of biomass production. Energy 2021, 220, 119731. [Google Scholar] [CrossRef]
- McLean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 199–224. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen total. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Agronomy Monograph 9; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Mehlich, A. Determination of cation- and anion-exchange properties of soils. Soil Sci. 1948, 66, 429–446. [Google Scholar] [CrossRef]
- Thomas, G.W. Exchangeable cations. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 159–165. [Google Scholar]
- Nelson, P.W.; Sommers, C.E. Total Carbon, organic Carbon and organic matter. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Martinelli, T.; Galasso, I. Phenological growth stages of Camelina sativa according to the extended BBCH scale. Ann. Appl. Biol. 2011, 158, 87–94. [Google Scholar] [CrossRef]
- Gesch, R.W. Influence of genotype and sowing date on camelina growth and yield in the north central U.S. Ind. Crop Prod. 2014, 54, 209–215. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef]
- Xie, Q.; Dash, J.; Huang, W.; Peng, D.; Qin, Q.; Mortimer, H.; Casa, R.; Pignatti, S.; Laneve, G.; Pasucci, S.; et al. Vegetation indices combining the red and red-edge spectral information for leaf area index retrieval. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 1482–1493. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.L.; Miller, L.D. Remote mapping of standing crop biomass for the estimation of the productivity of the short-grass prairie, Pawnee National Grasslands, Colorado. In Proceedings of the 8th International Symposium on Remote Sensing of Environment, Ann Arbor, MI, USA, 2–6 October 1972; pp. 1357–1381. [Google Scholar]
- Bannari, A.; Asalhi, H.; Teillet, P.M. Transformed difference vegetation index (TDVI) vor vegetation cover mapping. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Toronto, ON, Canada, 24–28 June 2002; Volume 5, pp. 3053–3055. [Google Scholar]
- Sun, H.Y.; Liu, C.M.; Zhang, X.Y.; Shen, Y.J.; Zhang, Y.Q. Effects of irrigation on water balance yield and WUE of winter wheat in the North China Plan. Agric. Water Manag. 2006, 85, 211–218. [Google Scholar] [CrossRef]
- Badura, G.P.; Bachmann, C.M.; Tyler, A.C.; Goldsmith, S.; Eon, R.S.; Lapszynsky, C.S. A novel approach for deriving LAI of salt marsh vegetation using structure from motion and multiangular spectra. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2019, 12, 599–613. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationship between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 331–354. [Google Scholar] [CrossRef]
- Mokhtari, A.; Noory, H.; Vazifedoust, M.; Bahrami, M. Estimating net irrigation requirement of winter theat using model and satellite based single and basal crop coefficient. Agric. Water Manag. 2018, 208, 95–106. [Google Scholar] [CrossRef]
- Lichtentahler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4. 3.1–F4. 3.8. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Lauenroth, W.K.; Roset, P.A. Estimating aboveground plant biomass using a photographic technique. Range Ecol. Manag. 2000, 53, 190–193. [Google Scholar] [CrossRef]
- Richardson, A.D.; Jenkis, J.P.; Braswell, B.H.; Hollinger, D.Y.; Ollinger, S.V.; Smith, M.L. Use of digital webcam images to track spring green-up in a deciduous broadleaf forest. Oecologia 2007, 152, 323–334. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.G.; Su, H.F.; Guo, W. Weed identification method based on probabilistic neural network in the corn seedlings field. In Proceedings of the 2010 International Conference on Machine Learning and Cybernetics, Haifa, Istrael, 11–14 July 2010; Volume 3, pp. 1528–1531. [Google Scholar]
- Liang, L.; Schwartz, M.D.; Fei, S. Photographic assessment of temperate forest understory phenology in relation to springtime meteorological drivers. Int. J. Biometeorol. 2012, 56, 343–355. [Google Scholar] [CrossRef]
- Nelson, S.A.; Khorram, S. Image Processing and Data Analysis with ERDAS IMAGINE®; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- ISTA (The International Seed Testing Association). International Rules for Seed Testing; ISTA: Bassersdorf, Switzerland, 2015; Volume 215. [Google Scholar]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Fazadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Lukas, V.; Huňady, I.; Kintl, A.; Mezera, J.; Hammerschmiedt, T.; Sobotková, J.; Brtnický, M.; Elbl, J. Using UAV to Identify the Optimal Vegetation Index for Yield Prediction of Oil Seed Rape (Brassica napus L.) at the Flowering Stage. Remote Sens. 2022, 14, 4953. [Google Scholar] [CrossRef]
- Janousek, J.; Jambor, V.; Marcoň, P.; Dohnal, P.; Synková, H.; Fiala, P. Using UAV-based photogrammetry to obtain correlation between the vegetation indices and chemical analysis of agricultural crops. Remote Sens. 2021, 13, 1878. [Google Scholar] [CrossRef]
- Tucker, C.J.; Holben, B.N.; Elgin Jr, J.H.; Mcmurtrey III, J.E. Relationship of spectral data to grain yield variation. Photogramm. Eng. Remote Sens. 1980, 46, 657–666. [Google Scholar]
- Curran, P.J.; Dungan, J.L.; Peterson, D.L. Estimating the foliar biochemical concentration of leaves with reflectance spectrometry: Testing the Kokaly and Clark methodologies. Remote Sens. Environ. 2001, 76, 349–359. [Google Scholar] [CrossRef]
- Zhao, D.; Li, J.; Qi, J. Hyperspectral characteristic analysis of a developing cotton canopy under different nitrogen treatments. Agronomy 2004, 24, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Jáuregui, J.M.; Delbino, F.G.; Bonvini, M.I.B.; Berhongaray, G. Determining yield of forage crops using the Canopeo mobile phone app. J. N. Z. Grassl. 2019, 81, 41–46. [Google Scholar] [CrossRef]
- Chung, Y.S.; Choi, S.C.; Silva, R.R.; Kang, J.W.; Eom, J.H.; Kim, C. Case study: Estimation of sorghum biomass using digital image analysis with Canopeo. Biomass Bioenergy 2017, 105, 207–210. [Google Scholar] [CrossRef]
- Reed, V.; Arnall, D.B.; Finch, B.; Bigatao Souza, J.L. Predicting winter wheat grain yield using fractional green canopy cover (FGCC). Int. J. Agron. 2021, 2021, 1443191. [Google Scholar] [CrossRef]
- Goodwin, A.W.; Lindsey, L.E.; Harrison, S.K.; Paul, P.A. Estimating wheat yield with normalized difference vegetation index and fractional green canopy cover. Crop Forage Turfgrass Manag. 2018, 4, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, P.K.; Kandel, H.J. Using canopy measurements to predict soybean seed yield. Agronomy 2021, 13, 3260. [Google Scholar] [CrossRef]
- Champolivier, L.; Merrien, A. Effects of water stress applied at different growth stages to Brassica napus L. var. oleifera on yield, yield components and seed quality. Eur. J. Agron. 1996, 5, 153–160. [Google Scholar]
- Leport, L.; Turner, N.C.; Davies, S.L.; Siddique, K.H.M. Variation in pod production and abortion among chickpea cultivation under terminal drought. Eur. J. Agron. 2006, 24, 236–246. [Google Scholar] [CrossRef]
- Ercoli, L.; Lulli, L.; Mariotti, M.; Masoni, A.; Arduini, I. Post-anthesis dry matter and nitrogen dynamics in durum wheat as affected by nitrogen supply and soil water availability. Eur. J. Agron. 2008, 28, 138–147. [Google Scholar] [CrossRef]
- Secchi, M.A.; Fernandez, J.A.; Stamm, M.J.; Durret, T.; Vara Prasad, P.V.; Messina, C.D.; Ciampitti, I.A. Effects of heat and drought on canola (Brassica napus L.) yield, oil, and protein: A meta-analysis. Field Crops Res. 2023, 293, 108848. [Google Scholar] [CrossRef]
- Ahmed, Z.; Liu, J.; Waraich, E.A.; Yan, Y.; Qi, Z.; Gui, D.; Zeng, F.; Tariq, A.; Shareed, M.; Iqbal, H.; et al. Differential physio-biochemical and yield responses of Camelina sativa L. under varying irrigation water regimes in semi-arid climatic conditions. PLoS ONE 2020, 15, e0242441. [Google Scholar] [CrossRef]
- Solis, A.; Vidal, I.; Paulino, L.; Johnson, B.L.; Berti, M.T. Camelina seed yield responses to nitrogen, sulphur, and phosphorus fertilizer in South Central Chile. Ind. Crops Prod. 2013, 44, 132–138. [Google Scholar] [CrossRef]
- Jankowsky, K.J.; Sokolski, M.; Kordan, B. Camelina: Yield and quality response to nitrogen and sulfur fertilization in Poland. Ind. Crops Prod. 2019, 141, 111776. [Google Scholar] [CrossRef]
- Bronson, K.F.; Hunsaker, D.J.; Thorp, K.R. Nitrogen fertilizer and irrigation effects on seed yield and oil in camelina. Agron. J. 2019, 111, 1712–1719. [Google Scholar] [CrossRef] [Green Version]
- Allen, B.L.; Lenssen, A.W.; Sainju, U.M.; Caesar-TonThat, T.; Evans, R.G. Nitrogen use in durum and selected Brassicaceae oilseed in two-year rotations. Agron. J. 2019, 106, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Afshar, R.K.; Lin, R.; Mohammed, Y.A.; Chen, C. Agronomic effects of urease and nitrification inhibitors on ammonia volatilization and nitrogen utilization in a dryland farming system: Field and laboratory investigation. J. Clean. Prod. 2018, 172, 4130–4139. [Google Scholar] [CrossRef]
- Mahli, S.S.; Johnson, E.N.; Hall, L.M.; May, W.E.; Phelps, S.; Nybo, B. Effects of nitrogen fertilizer application on seed yield, N uptake, and seed quality of Camelina sativa. Can. J. Soil Sci. 2014, 94, 35–47. [Google Scholar]
- Krzyżaniak, M.; Stolarski, M.J.; Tworkowski, J.; Puttick, D.; Eynck, C.; Załuski, D.; Kwiatkowski, J. Yield and seed composition of 10 spring camelina genotypes cultivated in the temperate climate of Central Europe. Ind. Crops Prod. 2019, 138, 111443. [Google Scholar] [CrossRef]




| Characteristics | Spring 2021 | Spring 2022 |
|---|---|---|
| Clay (<0.002 mm, %) | 15.1 | 14.6 |
| Silt (0.05–0.002 mm, %) | 35.9 | 42.0 |
| Sand (2–0.05 mm, %) | 48.9 | 43.4 |
| pH (H2O 1:2.5 soil:water suspension; McLean method) | 8.2 | 8.1 |
| Total Nitrogen (Kjeldahl method, g kg−1) | 0.9 | 1.2 |
| Organic matter (Walkley–Black method, %) | 2.1 | 2.8 |
| Available phosphorus (Olsen method, mg kg−1) | 3.4 | 8.0 |
| Exchangeable potassium (Thomas method, mg kg−1) | 65.0 | 146.0 |
| EC (μS cm−1) | 53.7 | 78.0 |
| CEC (Method BaCl2, pH 8.1, meq 100 g−1) | 13.7 | 8.4 |
| Growing Season | Cycle Length (Days) | GDD (°C d) | Cumulative Rainfall (mm) | |||
|---|---|---|---|---|---|---|
| Vegetative † | Reproductive †† | Vegetative | Reproductive | Vegetative | Reproductive | |
| Spring 2021 | 75 | 34 | 633.3 | 561.8 | 99.4 | 18.5 |
| Spring 2022 | 66 | 39 | 531.8 | 710.1 | 140.8 | 0 |
| Vegetation Indices (Vis) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Main Effects | NDVI | RVI | TNDVI | WvVi | Mcari2 | MSR705 | SAVI | FGGC |
| Y | *** | *** | *** | *** | ns | *** | *** | * |
| F | *** | ns | ns | ns | ns | *** | ns | *** |
| Y x F | ** | ns | ns | ns | ns | ns | ns | ns |
| Main Effects | Factor Level | Dualex | Plant Leaf Analysis | Soil Analysis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chls (μg cm−2) | Anth (Abs Units) | Flav (Abs Units) | NBI | Chl_a (μg cm−2) | Chl_b (μg cm−2) | Chl_a + b (μg cm−2) | Chl_a/b | Car (μg cm−2) | M (%) | NO3− | ||
| Year (Y) | 2021 | 24.9 ± 0.4 | 0.117 ± 0.002 | 1.15 ± 0.03 b | 21.8 ± 0.4 a | 9.6 ± 0.3 b | 4.3 ± 0.1 b | 13.8 ± 0.5 b | 2.3 ± 0.01 | 1.2 ± 0.01 b | 12.7 ± 0.2 b | 23.9 ± 4.7 b |
| 2022 | 24.1 ± 0.4 | 0.110 ± 0.003 | 1.29 ± 0.05 a | 19.1 ± 0.9 b | 17.0 ± 0.5 a | 7.3 ± 0.2 a | 24.4 ± 0.7 a | 2.3 ± 0.1 | 3.7 ± 0.1 a | 15.3 ± 0.2 a | 41.6 ± 7.5 a | |
| N fertilizer thesis (F) | T0 | 23.3 ± 0.7 | 0.114 ± 0.002 | 1.24 ± 0.10 | 19.4 ± 1.7 | 12.4 ± 1.7 b | 5.0 ± 0.7 b | 17.4 ± 2.3 b | 2.5 ± 0.1 a | 2.5 ± 0.6 | 14.1 ± 0.6 | 22.1 ± 6.1 |
| T1 | 25.1 ± 0.3 | 0.113 ± 0.005 | 1.18 ± 0.04 | 21.4 ± 0.9 | 14.7 ± 2.2 a | 6.4 ± 0.8 a | 21.1 ± 3.0 a | 2.3 ± 0.1 b | 2.7 ± 0.7 | 14.5 ± 0.7 | 40.0 ± 13.3 | |
| T2 | 24.8 ± 0.5 | 0.115 ± 0.004 | 1.29 ± 0.05 | 19.5 ± 1.1 | 13.2 ± 1.1 ab | 6.0 ± 0.6 a | 19.2 ± 1.7 ab | 2.2 ± 0.1 b | 2.3 ± 0.5 | 14.1 ± 0.6 | 21.6 ± 4.3 | |
| T3 | 24.9 ± 0.6 | 0.113 ± 0.003 | 1.16 ± 0.02 | 21.5 ± 0.3 | 12.9 ± 1.9 b | 5.8 ± 0.7 b | 18.7 ± 2.6 b | 2.2 ± 0.1 b | 2.3 ± 0.5 | 13.3 ± 0.6 | 47.3 ± 8.5 | |
| Significance | ||||||||||||
| Y | ns | ns | ** | ** | *** | *** | *** | ns | *** | *** | * | |
| F | ns | ns | ns | ns | ** | *** | ** | ** | ns | ns | ns | |
| Y × F | ns | ns | ** | ns | *** | ns | *** | ** | * | ns | ns | |
| Agronomic and Seed Quality Features | Main Effects | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year (Y) | N Fertilizer Regimes (F) | Significance | |||||||
| 2021 | 2022 | T0 | T1 | T2 | T3 | Y | F | YxF | |
| Plant height (cm) | 77.1 ± 1.4 a | 64.1 ± 2.0 b | 64.3 ± 4.2 b | 67.6 ± 3.2 b | 75.5 ± 2.6 a | 74.5 ± 2.8 a | *** | *** | ns |
| Branching (no. stems plant−1) | 6.8 ± 0.3 a | 6.0 ± 0.3 b | 5.9 ± 0.3 bc | 6.7 ± 0.2 b | 5.3 ± 0.2 c | 7.8 ± 0.3 a | ** | *** | ns |
| Plant density (plants m−2) | 153.5 ± 14.2 b | 235.5 ± 7.2 a | 168.9 ± 35.1 b | 175.9 ± 23.0 b | 217.4 ± 14.3 a | 215.9 ± 9.5 a | *** | ** | *** |
| no. Silique plant−1 | 224.2 ± 13.3 a | 132.0 ± 3.7 b | 193.3 ± 30.7 a | 180.4 ± 20.2 ab | 141.4 ± 18.8 b | 197.1 ± 23.1 a | *** | * | ns |
| no. Seeds silique−1 | 8.2 ± 0.2 a | 7.4 ± 0.1 b | 8.0 ± 0.4 | 7.6 ± 0.2 | 8.1 ± 0.2 | 7.6 ± 0.4 | * | ns | ns |
| Seed yield (Mg ha−1 DW) | 2.2 ± 0.1 a | 1.3 ± 0.1 b | 1.4 ± 0.1 b | 1.8 ± 0.2 a | 1.9 ± 0.3 a | 1.9 ± 0.2 a | *** | *** | * |
| TSW (g) | 1.1 ± 0.02 a | 1.1 ± 0.02 b | 1.0 ± 0.02 | 1.1 ± 0.03 | 1.1 ± 0.02 | 1.1 ± 0.02 | * | ns | ns |
| Oil content (% dry weight) | 34.2 ± 1.6 | 34.7 ± 0.97 | 34.4 ± 1.66 b | 38.8 ± 1.66 a | 30.7 ± 0.46 c | 33.9 ± 1.05 bc | ns | ** | *** |
| Oil yield (Mg ha−1) | 0.7 ± 0.04 a | 0.5 ± 0.02 b | 0.5 ± 0.04 b | 0.7 ± 0.08 a | 0.6 ± 0.09 a | 0.7 ± 0.07 a | *** | *** | * |
| Crude protein (%) | 23.7 ± 0.3 b | 25.2 ± 0.4 a | 23.3 ± 0.3 b | 24.8 ± 0.3 ab | 24.4 ± 0.3 ab | 25.3 ± 0.5 a | * | ** | ns |
| Parameters | Growing Season | VIs by UAV Images | Dualex | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NDVI | RVI | TNDVI | WvVi | MCARI2 | MSR705 | SAVI | FGCC | Chls | ||
| Seed yield | 2021 | 0.81 *** | −0.69 ** | 0.73 ** | 0.67 ** | 0.43 ns | 0.72 ** | 0.64 * | 0.74 ** | 0.50 * |
| 2022 | 0.53 * | 0.25 ns | −0.12 ns | −0.18 ns | −0.03 ns | 0.80 *** | −0.19 ns | 0.82 *** | 0.29 ns | |
| Leaf Chl content | 2021 | 0.51 * | −0.37 ns | 0.43 ns | 0.36 ns | 0.59 * | 0.35 ns | 0.32 ns | 0.37 ns | 0.51 *** |
| 2022 | 0.49 ns | −0.02 ns | 0.07 ns | 0.08 ns | 0.47 ns | 0.22 ns | 0.05 ns | 0.13 ns | 0.17 ns | |
| Leaf N% | 2021 | 0.68 * | −0.56 ns | 0.58 ns | 0.61 ns | 0.24 ns | 0.50 ns | 0.43 ns | 0.72 * | 0.92 *** |
| 2022 | −0.07 ns | −0.18 ns | 0.44 ns | 0.28 ns | 0.14 ns | 0.90 ** | 0.23 ns | 0.83 ** | 0.03 ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemente, C.; Ercolini, L.; Rossi, A.; Foschi, L.; Grossi, N.; Angelini, L.G.; Tavarini, S.; Silvestri, N. Spectral Response of Camelina (Camelina sativa (L.) Crantz) to Different Nitrogen Fertilization Regimes under Mediterranean Conditions. Agronomy 2023, 13, 1539. https://doi.org/10.3390/agronomy13061539
Clemente C, Ercolini L, Rossi A, Foschi L, Grossi N, Angelini LG, Tavarini S, Silvestri N. Spectral Response of Camelina (Camelina sativa (L.) Crantz) to Different Nitrogen Fertilization Regimes under Mediterranean Conditions. Agronomy. 2023; 13(6):1539. https://doi.org/10.3390/agronomy13061539
Chicago/Turabian StyleClemente, Clarissa, Leonardo Ercolini, Alessandro Rossi, Lara Foschi, Nicola Grossi, Luciana G. Angelini, Silvia Tavarini, and Nicola Silvestri. 2023. "Spectral Response of Camelina (Camelina sativa (L.) Crantz) to Different Nitrogen Fertilization Regimes under Mediterranean Conditions" Agronomy 13, no. 6: 1539. https://doi.org/10.3390/agronomy13061539