
Influence of Season and Organic Amendment on the Effectiveness of Different Biosolarization Treatments against Fusarium oxysporum f. sp. lactucae

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Location and Design of the Experiments
2.2. Production and Burial of the Inoculum with the Pathogen
2.3. Climate Variables Measured
2.4. Infectivity Trials in Lettuce
2.5. Microbiological Analysis in Soils
2.6. Statistical Analyses
3. Results
3.1. Evolution of Temperature and Oxygen Concentration during Biosolarization
3.2. Infectivity Trials
3.3. Soil Pathogen Quantification by qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FOLAC | Fusarium oxysporum f. sp. lactucae |
| SCM | Semi-composted manure |
References
- Gordon, T.R. Fusarium oxysporum and the Fusarium Wilt Syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Snyder, W.C.; Hansen, H.N. The species concept in Fusarium. Am. J. Bot. 1940, 27, 64–67. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. EPPO. 2023. Available online: https://gd.eppo.int/taxon/FUSALC (accessed on 1 July 2020).
- Rana, A.; Sahgal, M.; Johri, B.N. Fusarium oxysporum: Genomics, Diversity and Plant–Host Interaction. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S., Johri, B., Eds.; Springer: Singapore, 2017; pp. 159–199. [Google Scholar] [CrossRef]
- Guerrero, M.M.; Martínez, M.C.; León, M.; Armengol, J.; Monserrat, A. First Report of Fusarium Wilt of Lettuce Caused by Fusarium oxysporum f. sp. lactucae Race 1 in Spain. Plant Dis. 2019, 104, 1858. [Google Scholar] [CrossRef]
- Claerbout, J.; Van Poucke, K.; Mestdagh, H.; Delaere, I.; Vandevelde, I.; Venneman, S.; Decombel, A.; Bleyaert, P.; Neukermans, J.; Viaene, N.; et al. Fusarium isolates from Belgium causing wilt in lettuce show genetic and pathogenic diversity. Plant Pathol. 2023, 72, 593–609. [Google Scholar] [CrossRef]
- Gálvez, L.; Brizuela, A.M.; Garcés, I.; Cainarca, J.S.; Palmero, D. First Report of Fusarium oxysporum f. sp. lactucae Race 4 Causing Lettuce Wilt in Spain. Plant Dis. 2023; in press. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gilardi, G.; Gullino, M.L. Seed transmission of Fusarium oxysporum f.sp. lactucae. Phytoparasitica 2004, 32, 61–65. [Google Scholar] [CrossRef]
- Gilardi, G.; Vasileiadou, A.; Garibaldi, A.; Gullino, M.L. Low temperatures favour Fusarium wilt development by race 4 of Fusarium oxysporum f. sp. lactucae. J. Plant. Pathol. 2021, 103, 973–979. [Google Scholar] [CrossRef]
- Gordon, T.R.; Koike, S.T. Management of Fusarium wilt of lettuce. Crop Prot. 2015, 73, 45–49. [Google Scholar] [CrossRef]
- Al-Ani, L.K.T.; Furtado, E.L. The effect of incompatible plant pathogens on the host plant. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 47–57. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Rosskopf, E.N.; Serrano-Pérez, P.; Hong, J.; Shrestha, U.; Rodríguez-Molina, M.D.C.; Martin, K.; Kokalis-Burelle, N.; Shennan, C.; Muramoto, J.; Butler, D. Anaerobic soil disinfestation and soilborne pest management. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Meghvansi, M., Varma, A., Eds.; Springer: Cham, Switzerland, 2015; pp. 277–305. [Google Scholar]
- Attia, M.S.; Elsayed, S.M.; Abdelaziz, A.M.; Ali, M.M. Potential impacts of Ascophyllum nodosum, Arthrospira platensis extracts and calcium phosphite as therapeutic nutrients for enhancing immune response in pepper plant against Fusarium wilt disease. Biomass Conv. Bioref. 2023. [Google Scholar] [CrossRef]
- Attia, M.S.; Salem, M.S.; Abdelaziz, A.M. Endophytic fungi Aspergillus spp. reduce fusarial wilt disease severity, enhance growth, metabolism and stimulate the plant defense system in pepper plants. Biomass Conv. Bioref. 2022. [Google Scholar] [CrossRef]
- Katan, J. Solar heating (solarization) of soil for control of soilborne pests. Ann. Rev. Phytopathol. 1981, 19, 211–236. [Google Scholar] [CrossRef]
- Stapleton, J.J. Soil solarization in various agricultural production systems. Crop Prot. 2000, 19, 837–841. [Google Scholar] [CrossRef]
- Srivastava, J.N.; Ghatak, A. Biofumigation: A control method for the soil-borne diseases. Int. J. Plant Prot. Phytopathol. 2017, 10, 453–460. [Google Scholar] [CrossRef]
- Coelho, L.; Mitchell, D.J.; Chellemi, D.O. Thermal inactivation of Phytophthora nicotianae. Phytopathology 2000, 90, 1089–1097. [Google Scholar] [CrossRef]
- Domínguez, P.; Miranda, L.; Soria, C.; de los Santos, B.; Chamorro, M.; Romero, F.; Daugovish, O.; López-Aranda, J.M.; Medina, J.J. Soil biosolarization for sustainable strawberry production. Agron. Sustain. Dev. 2014, 34, 821–829. [Google Scholar] [CrossRef]
- Guerrero, M.; Lacasa, C.M.; Martínez, V.; Martínez, M.C.; Larregla, S.; Lacasa, A. Soil biosolarization for Verticillium dahliae and Rhizoctonia solani control in artichoke crops in southeastern Spain. Span. J. Agric. 2019, 17, e1002. [Google Scholar] [CrossRef]
- Martínez, M.A.; Martínez, M.C.; Bielza, P.; Tello, J.; Lacasa, A. Effect of biofumigation with manure amendments and repeated biosolarization on Fusarium densities in pepper crops. J. Ind. Microbiol. Biotechnol. 2011, 38, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Cascone, G.; D’Emilio, A.; Polizzi, G.; Grillo, R. Effectiveness of greenhouse soil solarization with different plastic mulches in controlling corky root and rootknot on tomato plants. Acta Hortic. 2000, 532, 145–150. [Google Scholar] [CrossRef]
- Chellemi, D.O.; Mirusso, J. Optimizing soil disinfestation procedures for fresh market tomato and pepper production. Plant Dis. 2006, 90, 668–674. [Google Scholar] [CrossRef]
- Vitale, A.; Castello, I.; Cascone, G.; D’Emilio, A.; Mazzarella, R.; Polizzi, G. Reduction of corky root infections on greenhouse tomato crops by soil solarization in South Italy. Plant Dis. 2011, 95, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.; Katan, J.; Gamliel, A. Soil suppressiveness to Meloidogyne javanica as induced by organic amendments and solarization in greenhouse crops. Crop Prot. 2012, 39, 26–32. [Google Scholar] [CrossRef]
- Mauromicale, G.; Monaco, A.L.; Longo, A.M.G. Improved efficiency of soil solarization for growth and yield of greenhouse tomatoes. Agron. Sustain. Dev. 2010, 30, 753–761. [Google Scholar] [CrossRef]
- Garibaldi, A.; Tamietti, G. Attempts to use soil solarization in closed glasshouses in northern Italy for controlling corky root of tomato. Acta Hortic. 1984, 152, 237–244. [Google Scholar] [CrossRef]
- Oka, Y.; Shapira, N.; Fine, P. Control of root-knot nematodes in organic farming systems by organic amendments and soil solarization. Crop Prot. 2007, 26, 1556–1565. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; McSorley, R.; Stansly, P.A. Effects of long-term organic amendments and soil sanitation on weed and nematode populations in pepper and watermelon crops in Florida. Crop Prot. 2012, 41, 106–112. [Google Scholar] [CrossRef]
- Oka, Y. Mechanisms of nematode suppression by organic soil amendments—A review. Appl. Soil Ecol. 2010, 44, 101–115. [Google Scholar] [CrossRef]
- OJ. Directive 91/676/EEC of the Council of the European Communities of December 12 concerning the protection of waters against pollution caused by nitrates from agricultural sources. Off. J. Eur. Union L 1991, 375, 1–8. [Google Scholar]
- Gamliel, A.; Austerweil, M.; Kitzman, G. Non chemical approach to soilborne pest management with organic amendments. Crop Prot. 2000, 19, 847–853. [Google Scholar] [CrossRef]
- Momma, N.; Kobara, Y.; Momma, M. Fe2+ and Mn2+; potential agents to induce suppression of Fusarium oxysporum for biological soil disinfestation. J. Gen. Plant Pathol. 2011, 77, 331–335. [Google Scholar] [CrossRef]
- Katase, M.; Kubo, C.; Ushio, S.; Ootsuka, E.; Takeuchi, T.; Mizukub, T. Nematicidal activity of volatile fatty acids generated from wheat bran in reductive soil disinfestation. Jpn. J. Nematol. 2009, 39, 53–62. [Google Scholar] [CrossRef]
- Agtmaal, M.; Os, G.J.; Gera Hol, W.H.; Hundscheid, M.P.J.; Runia, W.T.; Hordijk, C.A.; De Boer, W. Legacy effects of anaerobic soil disinfestation on soil bacterial community composition and production of pathogen suppressing volatiles. Front. Microbiol. 2015, 6, 701. [Google Scholar] [CrossRef] [PubMed]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, J.; Bollen, G.J. Control of soilborne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Davison, E.M.; Tay, F.C.S. The effect of cinnamomi waterlogging on infection of Eucalyptus marginata seedlings by Phytophthora. New Phytol. 1987, 105, 585–594. [Google Scholar] [CrossRef]
- Butler, D.M.; Kokalis-Burelle, N.; Albano, J.P.; McCollum, T.G.; Muramoto, J.; Shennan, C.; Rosskopf, E.N. Anaerobic soil disinfestation (ASD) combined with soil solarization as a methyl bromide alternative: Vegetable crop performance and soil nutrient dynamics. Plant Soil 2014, 378, 365–381. [Google Scholar] [CrossRef]
- Rosskopf, E.; Di Gioia, F.; Hong, J.C.; Pisani, C.; Kokalis-Burelle, N. Organic amendments for pathogen and nematode control. Annu. Rev. Phytopathol. 2020, 58, 277–311. [Google Scholar] [CrossRef]
- Gandariasbeitia, M.; Ojinaga, M.; Orbegozo, E.; Mendarte, S.; Larregla, S. Winter biodisinfestation with Brassica green manure is a promising management strategy for Phytophthora capsici control of protected pepper crops in humid temperate climate regions of northern Spain. Span. J. Agric. Res. Arch. 2019, 17, e1005. [Google Scholar] [CrossRef]
- Ojinaga, M.; Gandariasbeitia, M.; Orbegozo, E.; Ortíz, A.; Guerrero, M.M.; Lacasa, C.M.; Arizmendi, J.; Otaño, A.; Camino, C.; Enbeita, G.; et al. Biodisinfestation for Meloidogyne and Verticillium control in commercial protected crops in the Basque Country Atlantic area (northern Spain). Acta Hortic. 2020, 1270, 327–336. [Google Scholar] [CrossRef]
- Tsao, P.H.; Oster, J.J. Relation of Ammonia and Nitrous Acid to Suppression of Phytophthora in Soils Amended with Nitrogenous Organic Substances. Phytopathology 1981, 171, 53–59. [Google Scholar] [CrossRef]
- Tenuta, M.; Lazarovits, G. Ammonia and Nitrous Acid from Nitrogenous Amendments Kill the Microsclerotia of Verticillium dahliae. Phytopathology 2002, 92, 255–264. [Google Scholar] [CrossRef]
- Markakis, E.A.; Tjamos, S.E.; Chatzipavlidis, I.; Antoniou, P.P.; Paplomatas, E.J. Evaluation of compost amendments for control of vascular wilt diseases. J. Phytopathol. 2008, 156, 622–627. [Google Scholar] [CrossRef]
- Guerrero, M.M.; Lacasa, C.M.; Martínez, V.; Martínez, M.C.; Monserrat, A.; Larregla, S. Low temperature biodisinfection effectiveness for Phytophthora capsici control of protected sweet pepper crops in the Southeast of Spain. Front. Sustain. Food Syst. 2021, 5, 659290. [Google Scholar] [CrossRef]
- Etxeberria, A.; Mendarte, S.; Larregla, S. Thermal inactivation of Phytophthora capsici oospores. Rev. Iberoam. Micol. 2011, 28, 83–90. [Google Scholar] [CrossRef]
- Lacasa, C.M.; Martínez, V.; Hernández, A.; Ros, C.; Lacasa, A.; Guerrero, M.M. Surival reduction of Phytophthora capsici oospores and P. nicotianae chlamydospores with Brassica green manures combined with solarization. Sci. Hortic. 2015, 197, 607–618. [Google Scholar] [CrossRef]



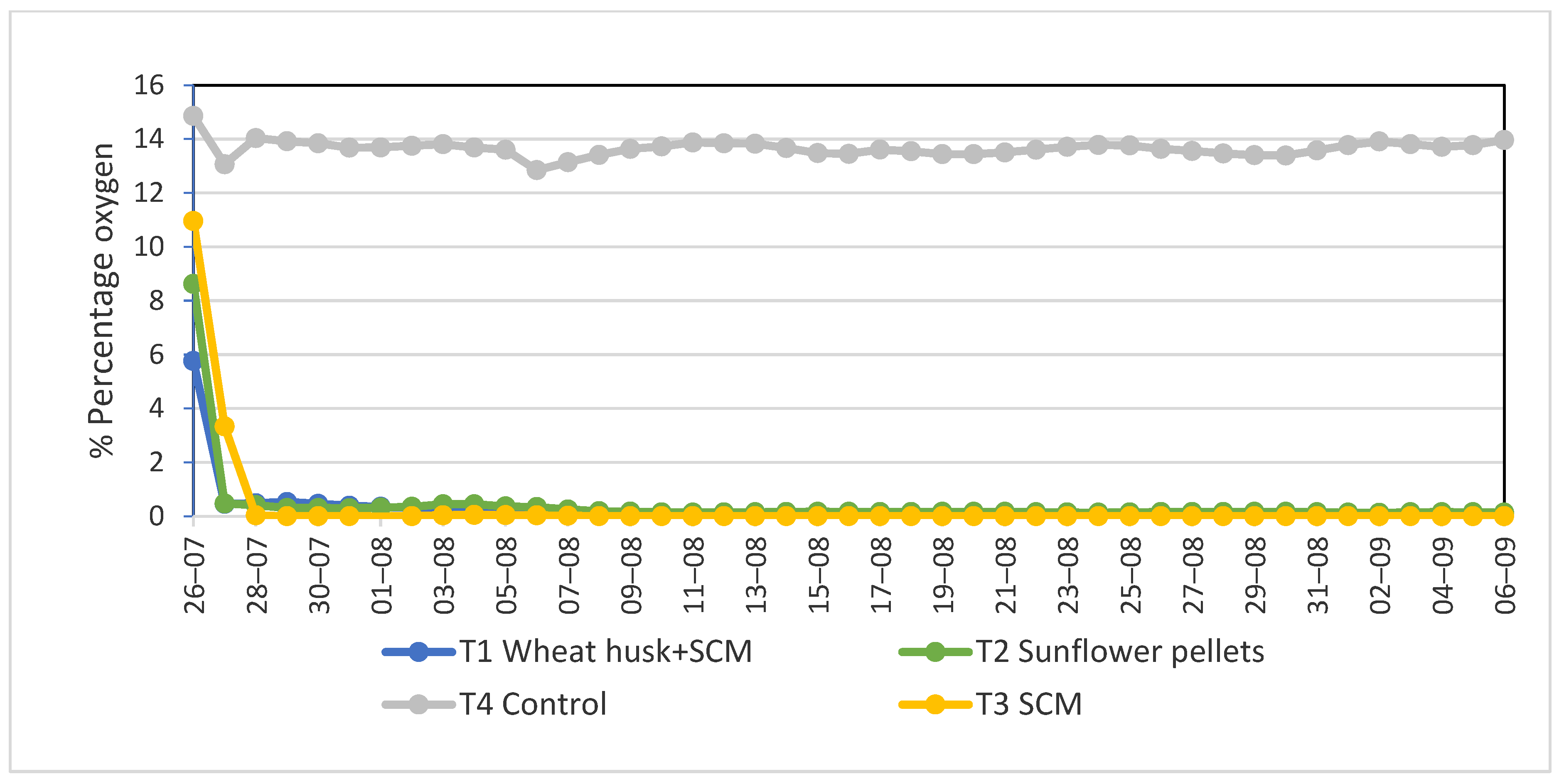{kind=link}
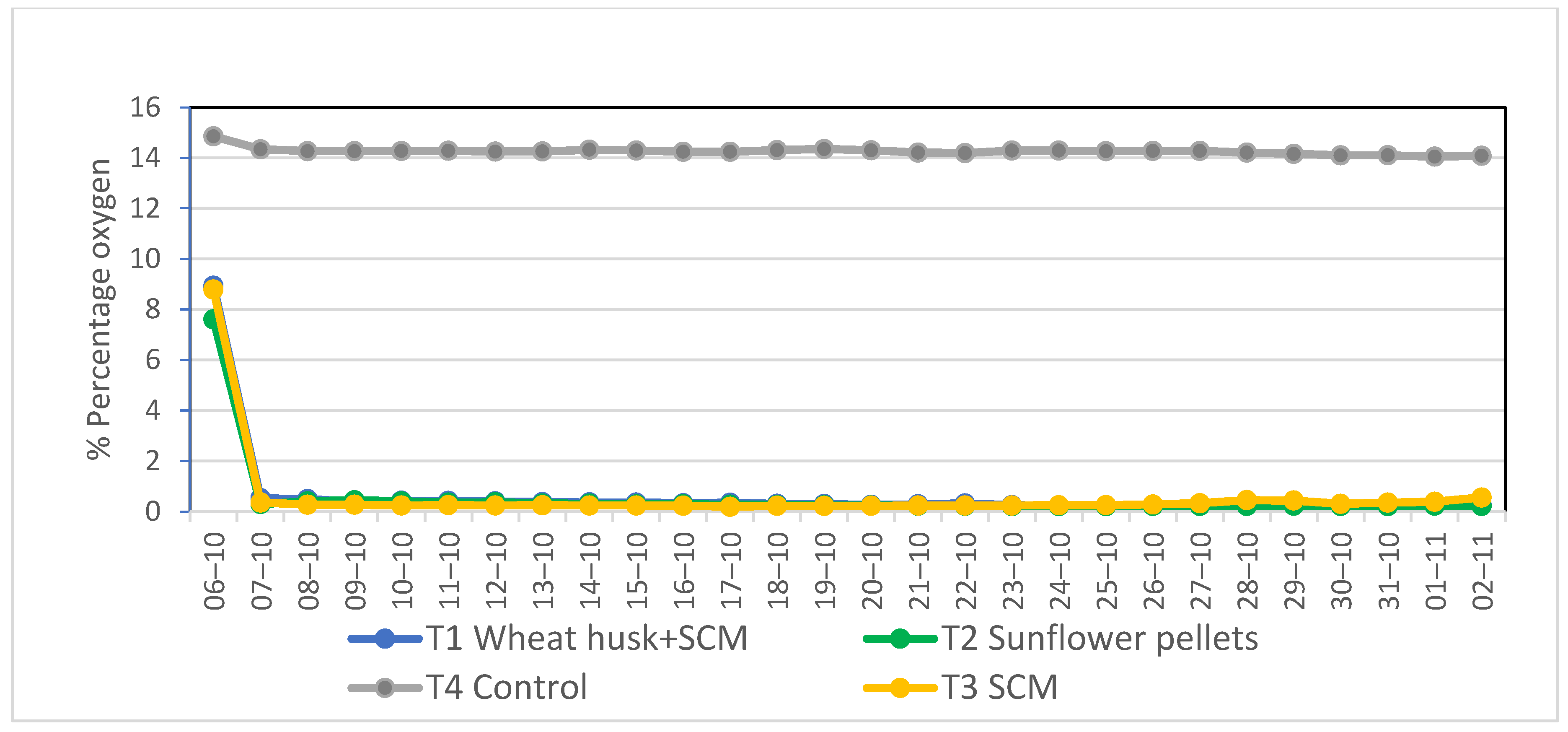{kind=link}
| Treatments | Depths | Season | <38 °C | >38–40 °C | 40–42 °C | >42 °C |
|---|---|---|---|---|---|---|
| T1: Wheat husk + SCM | 15 cm | Sumer | 147 | 186 | 236 | 469 |
| 30 cm | 96.5 | 176.5 | 312 | 453 | ||
| T2: Sunflower pellets | 15 cm | 68 | 71 | 199 | 700 | |
| 30 cm | 712 | 99 | 150 | 77 | ||
| T3: SCM | 15 cm | 130 | 197 | 215 | 496 | |
| 30 cm | 290.5 | 424.5 | 323 | 0 | ||
| T4: Control | 15 cm | 703 | 144 | 131 | 60 | |
| 30 cm | 641 | 351 | 44 | 0 | ||
| T1: Wheat husk+ SCM | 15 cm | Autumn | 1008 | 3 | 0 | 0 |
| 30 cm | 1011 | 0 | 0 | 0 | ||
| T2: Sunflower pellets | 15 cm | 1006 | 5 | 0 | 0 | |
| 30 cm | 1011 | 0 | 0 | 0 | ||
| T3: SCM | 15 cm | 1011 | 0 | 0 | 0 | |
| 30 cm | 1011 | 0 | 0 | 0 | ||
| T4: Control | 15 cm | 1011 | 0 | 0 | 0 | |
| 30 cm | 1011 | 0 | 0 | 0 |
| Treatments | Percentage of Affected Plants | |||
|---|---|---|---|---|
| Season | Summer | Autumn | ||
| Depth | 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm |
| T1: Wheat husks + SCM | 0 a | 0 a | 0 a | 10 a |
| T2: Sunflower pellets | 0 a | 0 a | 0 a | 0 a |
| T3: SCM | 0 a | 0 a | 10 a | 20 a |
| T4: Control | 40 b | 40 b | 100 b | 100 b |
| Treatments | N° Sequences/g Soil | |||
|---|---|---|---|---|
| Season | Summer | Autumn | ||
| Depth | 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm |
| T1: Wheat husk + SCM | 0 a | 0 a | 22 ns | 145 a |
| T2: Sunflower pellets | 0 a | 0 a | 0 | 0 a |
| T3: SCM | 0 a | 0 a | 1248 | 507 a |
| T4: Control | 2349 b | 1428 b | 2255 | 4871 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero, M.d.M.; Lacasa, C.M.; Martínez, V.; Monserrat, A.; López-Pérez, J.A.; Ortega, R.; Nieto, J.C.; Miralles, I.; Larregla, S. Influence of Season and Organic Amendment on the Effectiveness of Different Biosolarization Treatments against Fusarium oxysporum f. sp. lactucae. Agronomy 2023, 13, 1498. https://doi.org/10.3390/agronomy13061498
Guerrero MdM, Lacasa CM, Martínez V, Monserrat A, López-Pérez JA, Ortega R, Nieto JC, Miralles I, Larregla S. Influence of Season and Organic Amendment on the Effectiveness of Different Biosolarization Treatments against Fusarium oxysporum f. sp. lactucae. Agronomy. 2023; 13(6):1498. https://doi.org/10.3390/agronomy13061498
Chicago/Turabian StyleGuerrero, María del Mar, Carmen Mª Lacasa, Victoriano Martínez, Antonio Monserrat, José Antonio López-Pérez, Raúl Ortega, José Carlos Nieto, Isabel Miralles, and Santiago Larregla. 2023. "Influence of Season and Organic Amendment on the Effectiveness of Different Biosolarization Treatments against Fusarium oxysporum f. sp. lactucae" Agronomy 13, no. 6: 1498. https://doi.org/10.3390/agronomy13061498