

Metabolic, Nutritional and Morphophysiological Behavior of Eucalypt Genotypes Differing in Dieback Resistance in Field When Submitted to PEG-Induced Water Deficit
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Estimation of the Expected Osmotic Potential of the Substrate Solution
2.4. Morphological Variables
2.5. Physiological Parameters
2.6. Nutritional Variables
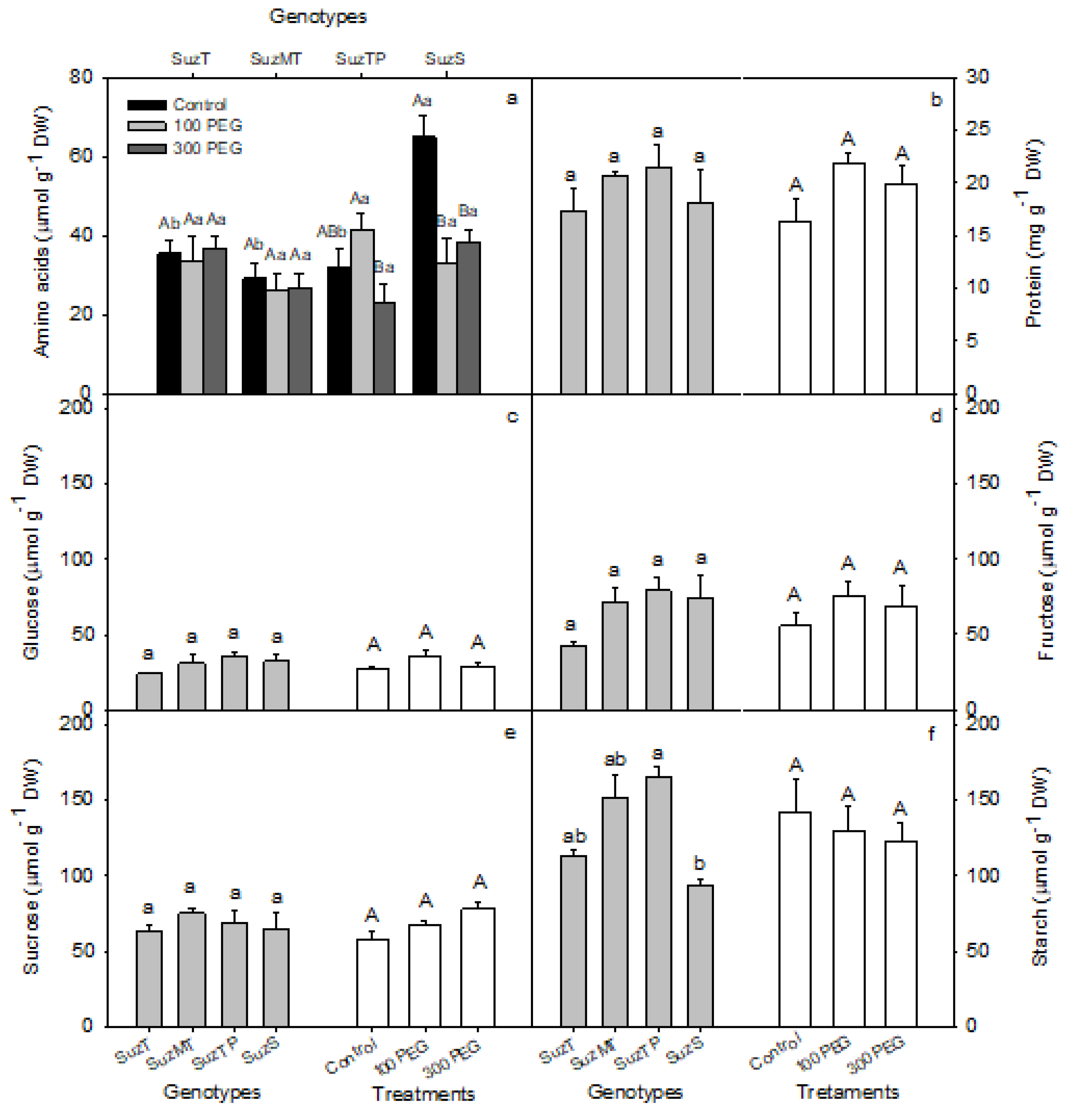
2.7. Biochemical Analyses
2.8. Metabolites Profile Determination
2.9. Statistical Analysis and Experimental Design
3. Results
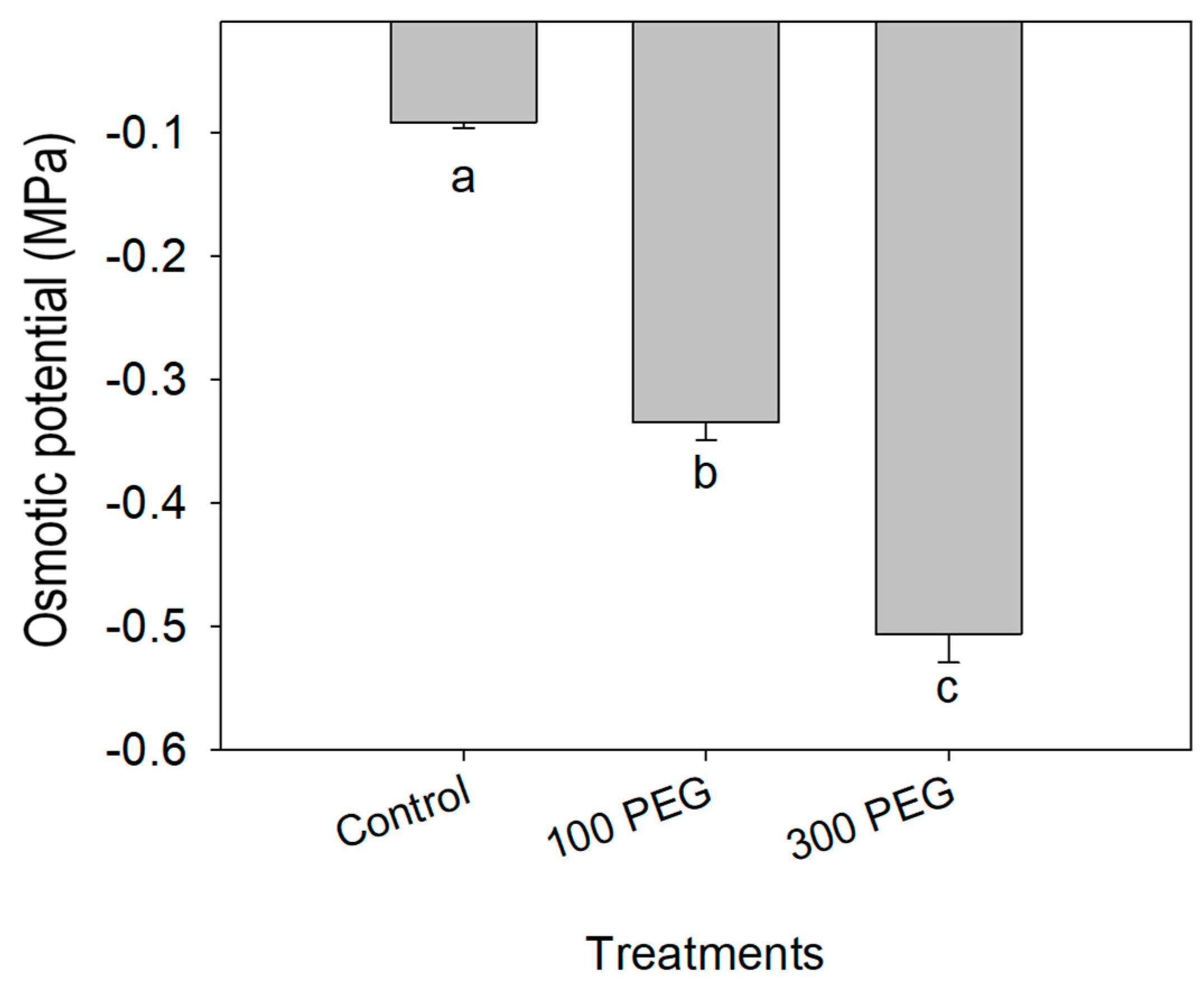
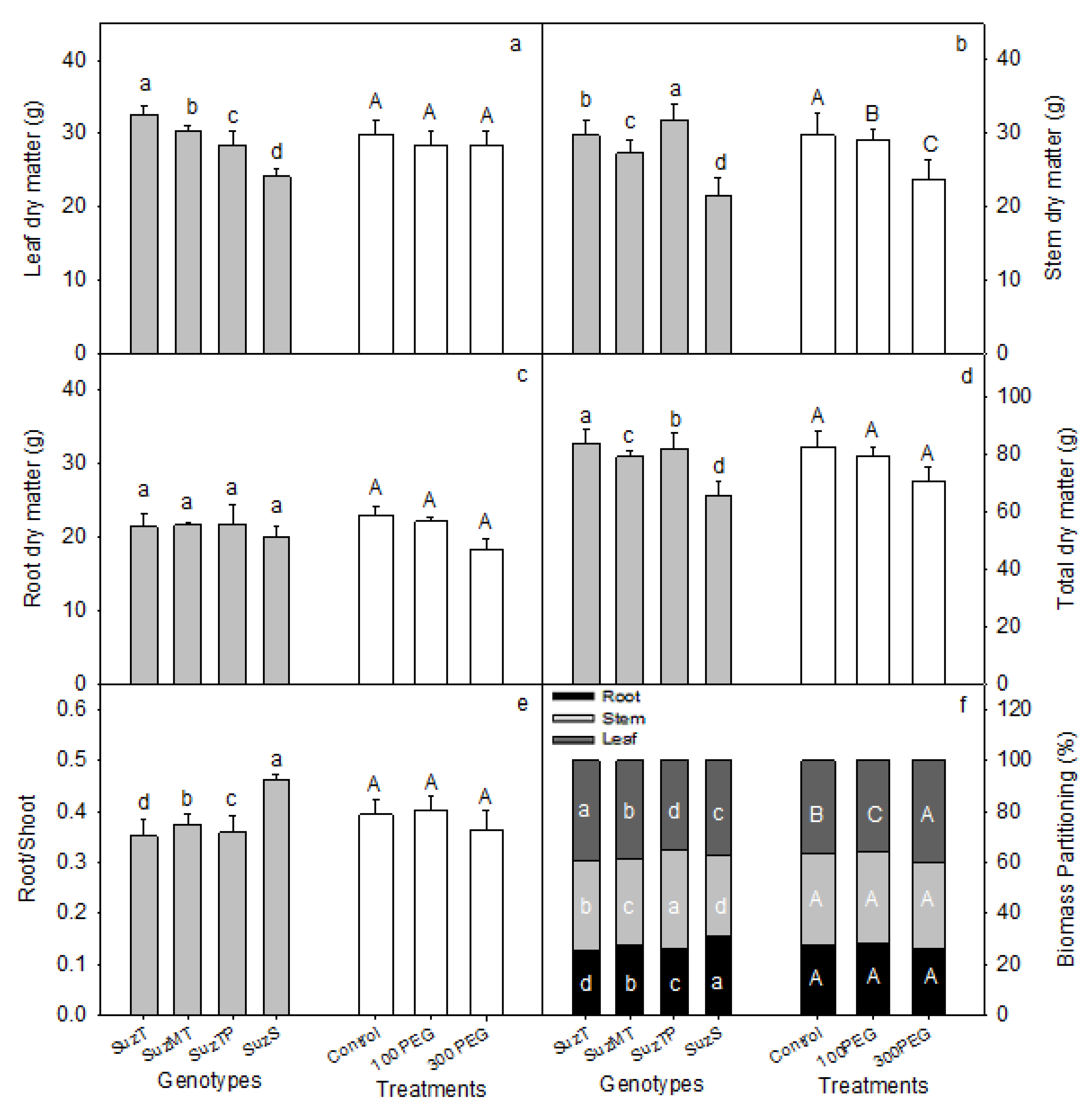
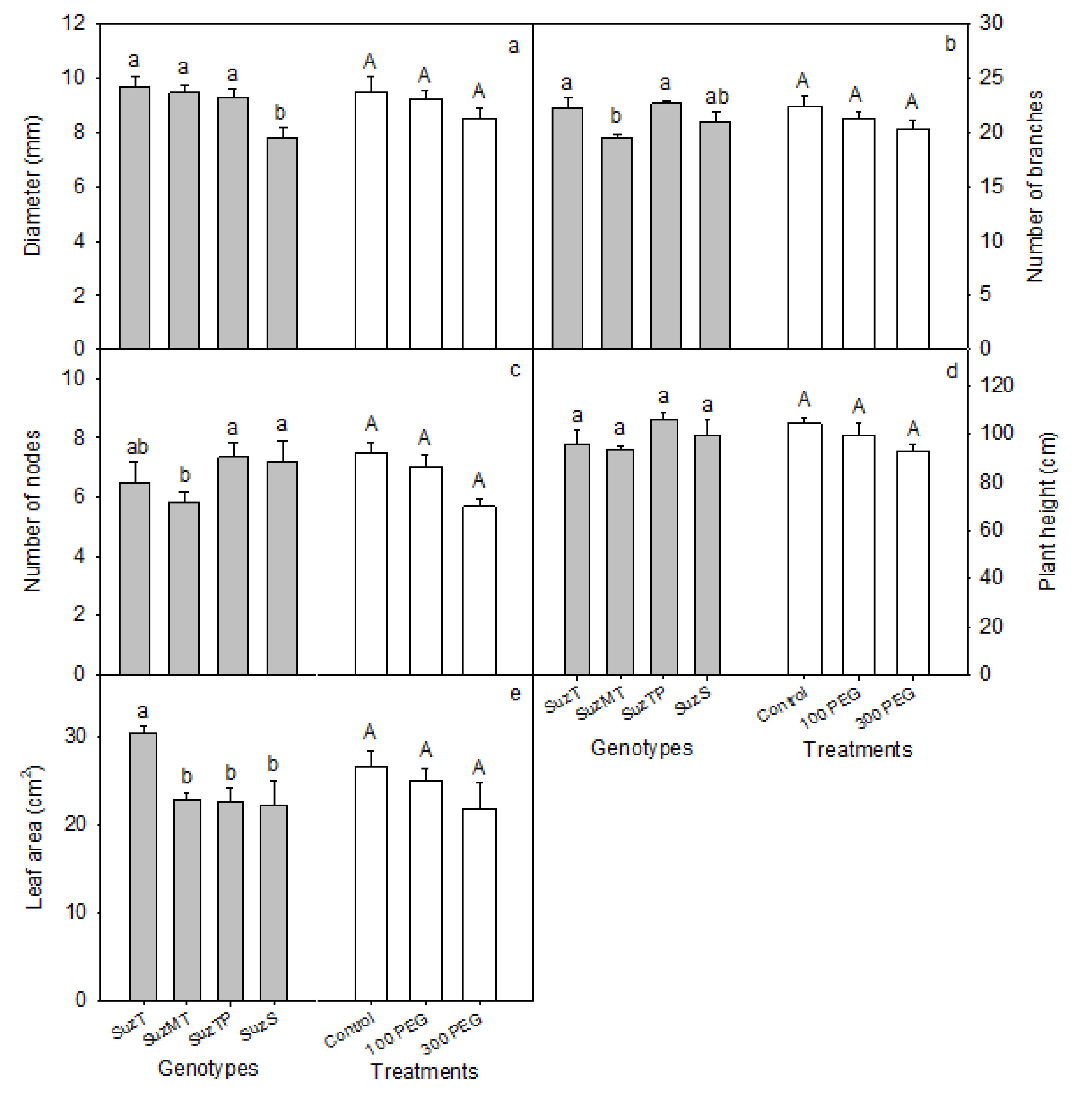
3.1. Osmotic Potential of Substrate Solution and Morphological Differences between Genotypes
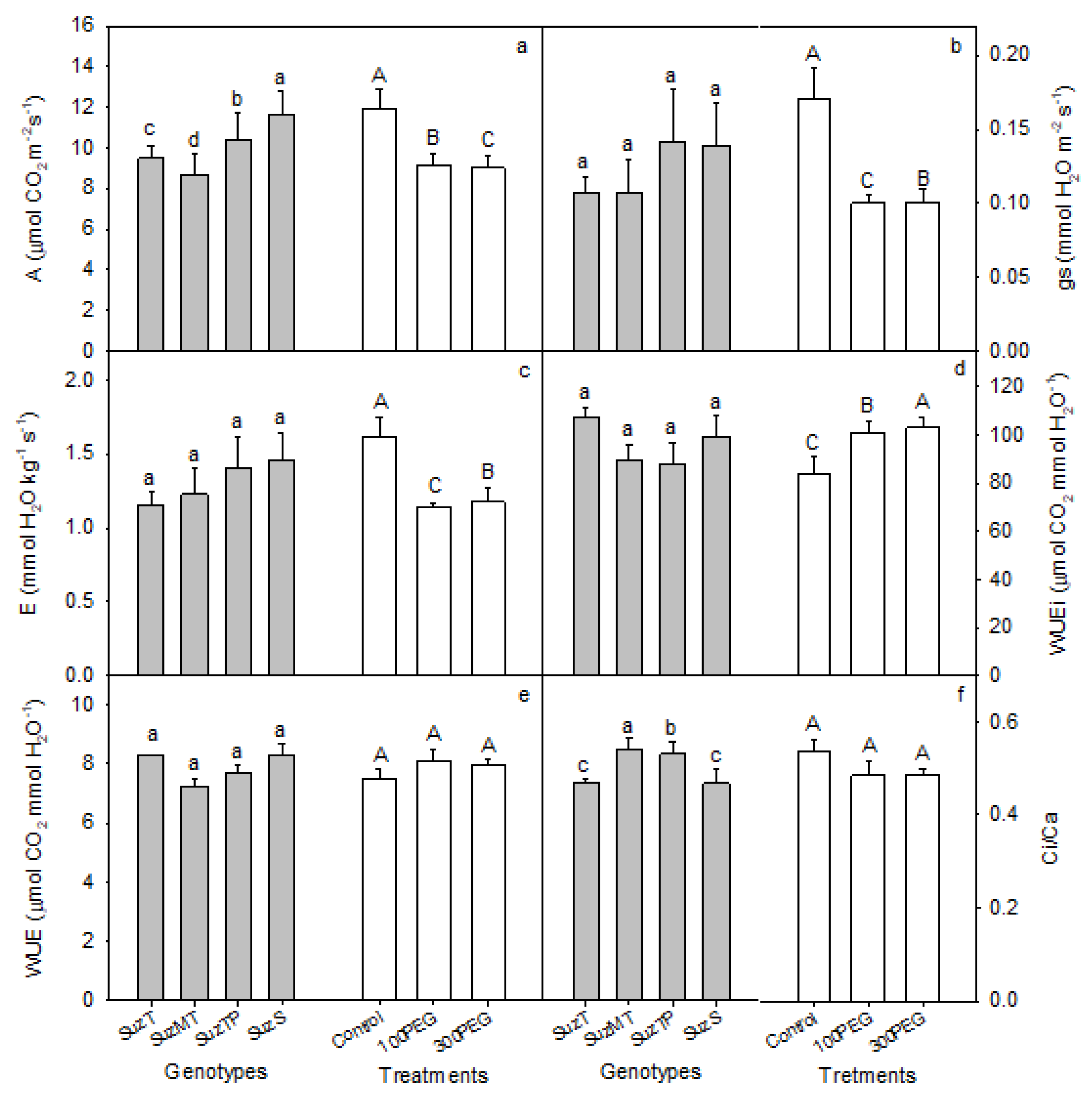
3.2. Gas Exchange Analysis
3.3. Nutritional Status of Genotypes with Distinct Dieback Tolerance
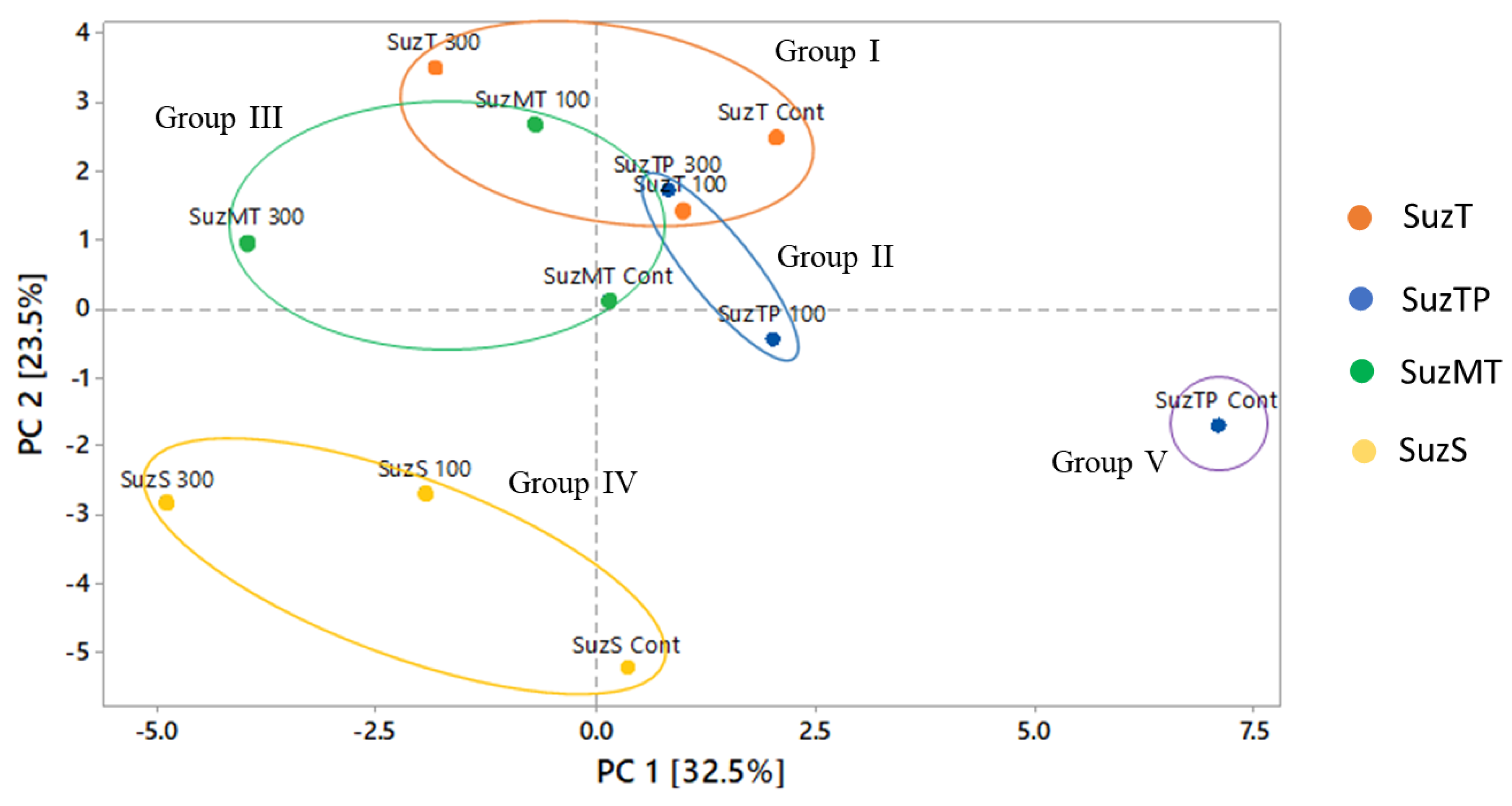
3.4. Multivariate Analysis
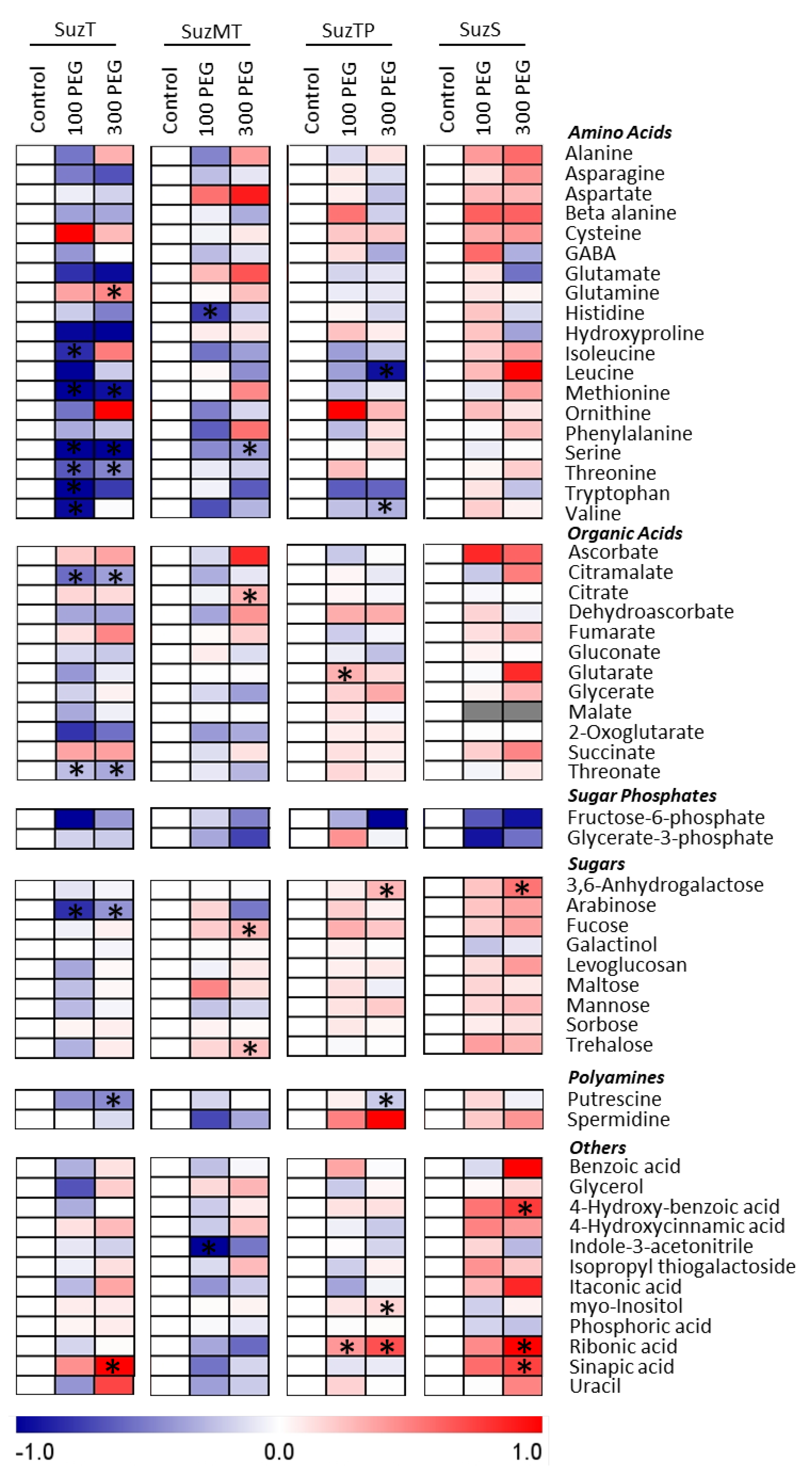
3.5. Effects of Water Deficit on Leaf Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jurskis, V. Eucalypt decline in Australia, and a general concept of tree decline and dieback. For. Ecol. Manag. 2005, 215, 1–20. [Google Scholar] [CrossRef]
- Dianese, J.G.; Haridasan, M.; Moraes, T.S.A. Tolerance to ‘Mal do Rio Doce’, a major disease of Eucalyptus in Brazil. Trop. Pest. Manag. 1984, 30, 247–252. [Google Scholar] [CrossRef]
- Leite, F.P.; Novais, R.F.; Silva, I.R.; Barros, N.F.; Neves, J.C.L.; Medeiros, A.G.B.; Ventrella, M.C.; Villani, E.M.A. Manganese accumulation and its relation to “eucalyptus shoot blight in the Vale do Rio Doce”. Rev. Bras. Ciênc. 2014, 38, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Fensham, R.J.; Holman, J.E. Temporal and spatial patterns in drought-related tree dieback in Australian Savanna. J. Appl. Ecol. 1999, 36, 1035–1050. [Google Scholar] [CrossRef] [Green Version]
- Worrall, J.J.; Egeland, L.; Eager, T.; Mask, R.A.; Johnson, E.W.; Kemp, P.A.; Shepperd, W.D. Rapid mortality of Populus tremuloides in southwestern Colorado, USA. For. Ecol. Manag. 2008, 255, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Ma, Z.; Lei, X.; Zhu, Q.; Chen, H.; Wang, W.; Liu, S.; Li, W.; Fang, X.; Zhou, X. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increase climate-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, T.R.; Picoli, E.A.T.; Souza, G.A.; Condé, S.A.; Silva, N.M.; Lopes-Mattos, K.L.B.; Resende, M.D.V.; Zauza, E.A.V.; Oda, S. Phenotypic markers in early selection for tolerance to dieback in Eucalyptus. Ind. Crop. Prod. 2017, 107, 130–138. [Google Scholar] [CrossRef]
- Nogueira, R.J.M.C.; Moraes, J.A.P.V.; Burity, H.A.; Bezerra Neto, E. Alterações na resistência à difusão de vapor das folhas e relações hídricas em aceroleiras submetidas à déficit de água. Rev. Bras. Fisiol. Veg. 2001, 13, 75–87. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Maroco, J.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Groom, P.K.; Lamont, B.B.; Poot, P.; Prior, L.D.; Reich, P.B.; Schulze, E.D.; Veneklaas, E.J.; Westoby, M. Leaf trait relationships in Australian plant species. Funct. Plant Biol. 2004, 31, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Callister, A.N.; Arndt, S.K.; Adams, M.A. Comparison of four methods for measuring osmotic potential of tree leaves. Physiol. Plant. 2006, 127, 383–392. [Google Scholar] [CrossRef]
- Dias, L.P.R.; Gatiboni, L.P.; Miquelutti, D.J.; Brunetto, G.; Campos, D.J.P. Boron in preventing of shoot dieback on Eucalyptus benthamii grown in soil with different water regimes. Rev. Sci. Agrar. 2017, 18, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Picoli, E.A.T.; de Resende, M.D.V.; Oda, S. Come Hell or High Water: Breeding the Profile of Eucalyptus Tolerance to Abiotic Stress Focusing Water Deficit. In Plant Growth and Stress Physiology; Gupta, D.K., Palma, J.M., Eds.; Plant in Challenging Environments; Springer: Cham, Switzerland, 2021; Volume 3. [Google Scholar] [CrossRef]
- White, T.C.R. Weather, Eucalyptus Dieback in New England, and a General Hypothesis of the Cause of Dieback. Pac. Sci. 1986, 40, 58–78. Available online: http://hdl.handle.net/10125/1005 (accessed on 20 April 2023).
- Mattiello, E.M.; Ruiz, H.A.; Silva, I.R.; Barros, N.F.; Neves, J.C.L.; Behling, M. Transporte de boro no solo e sua absorção por eucalipto. Rev. Bras. Ciênc. 2009, 33, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Shenker, M.; Plessner, O.P.; Tel-Or, E. Manganese nutrition effects on tomato growth, chlorophyll concentration, and superoxide dismutase activity. J. Plant Physiol. 2004, 161, 197–202. [Google Scholar] [CrossRef]
- Silva, F.C.; Shvaleva, A.; Maroco, J.; Almeida, M.H. Responses to water stress in two Eucalyptus globulus clones differing in drought tolerance. Tree Physiol. 2004, 4, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Pita, P.; Cañas, I.; Soria, F.; Ruiz, F.; Toval, G. Use of physiological traits in tree breeding for improved yield in drought prone environments. The case of Eucalyptus globulus. For. Syst. 2005, 14, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Amrutha, S.; Parveen, A.B.M.; Muthupandi, M.; Sivakumar, V.; Nautiyal, R.; Dasgupta, M.G. Variation in morpho-physiological, biochemical and molecular responses of two Eucalyptus species under short-term water stress. Acta Bot. Croat. 2019, 78, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Correia, B.; Pintó-Marijuan, M.; Castro, B.B.; Brossa, R.; López-Carbonell, M.; Pinto, G. Hormonal dynamics during recovery from drought in two Eucalyptus globulus genotypes: From root to leaf. Plant Physiol. Biochem. 2014, 82, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Correia, B.; Pintó-Marijuan, M.; Neves, L.; Brossa, R.; Dias, M.C.; Costa, A.; Castro, B.B.; Araújo, C.; Santos, C.; Chaves, M.M.; et al. Water stress and recovery in the performance of two Eucalyptus globulus clones: Physiological and biochemical profiles. Physiol. Plant 2014, 150, 580–592. [Google Scholar] [CrossRef]
- Valdés, A.E.; Irar, S.; Majada, J.P.; Rodríguez, A.; Fernández, B.; Pagès, M. Drought tolerance acquisition in Eucalyptus globulus (Labill.): A research on plant morphology, physiology and proteomics. J. Proteom. 2013, 79, 263–276. [Google Scholar] [CrossRef]
- Warren, C.R.; Aranda, I.; Cano, F.J. Metabolomics demonstrates divergent responses of two Eucalyptus species to water stress. Metabolomics 2012, 8, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.N.; Picoli, E.A.T.; Souza, G.A.; Farag, M.A.; Scotti, M.T.; Barbosa Filho, J.M.; da Silva, M.S.; Tavares, J.F. Phenolics metabolismo provides a tool for screening drought tolerant Eucalyptus grandis hybrids. Austr. J. Crop Sci. 2017, 11, 1016–1025. [Google Scholar] [CrossRef]
- Correia, B.; Valledor, L.; Hancock, R.D.; Renaut, J.; Pascual, J.; Soares, A.M.V.M.; Pinto, G. Integrated proteomics and metabolomics to unlock global and clonal responses of Eucalyptus globulus recovery from water deficit. Metabolomics 2016, 12, 141. [Google Scholar] [CrossRef]
- Adams, M.A.; Richter, A.; Hill, A.; Colmer, T. Salt tolerance in Eucalyptus spp.: Identity and response of putative osmolytes. Plant Cell Environ. 2005, 28, 772–787. [Google Scholar] [CrossRef]
- Merchant, A.; Tausz, M.; Arndt, S.K.; Adams, M.A. Cyclitols and carbohydrates in leaves and roots of 13 Eucalyptus species suggest contrasting physiological responses to water deficit. Plant Cell Environ. 2006, 29, 2017–2029. [Google Scholar] [CrossRef]
- Warren, C.R.; Bleby, T.; Adams, M.A. Changes in gas exchange versus leaf solutes as a means to cope with summer drought in Eucalyptus marginata. Oecologia 2007, 154, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, T.R.; Picoli, E.A.d.T.; Pereira, W.L.; Condé, S.A.; Resende, R.T.; de Resende, M.D.V.; da Costa, W.G.; Cruz, C.D.; Zauza, E.A.V. Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones. Agronomy 2023, 13, 937. [Google Scholar] [CrossRef]
- Szijarto, L.; van de Voort, F.R. Determination of added water and bovine milk to caprine milk. J. Dairy Sci. 1983, 66, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Baer, R.J.; Baldwin, K.A. Freezing Points of Bulking Agents Used in Manufacture of Low-Calorie Frozen Desserts. J. Dairy Sci. 1984, 67, 2860–2862. [Google Scholar] [CrossRef]
- Henriques, G.S.; Rosado, G.P. Formulação de dietas enterais artesanais e determinação da osmolalidade pelo método crioscópico. Rev. Nutr. 1999, 12, 225–232. [Google Scholar] [CrossRef] [Green Version]
- George, E.F.; Hall, M.A.; De Klerk, G.J. Plant growth regulators I: Introduction; auxins, their analogues and inhibitors. In Plant Propagation by Tissue Culture; Springer: Dordrecht, The Netherlands, 2008; pp. 115–174. [Google Scholar]
- Sarruge, J.R.; Haag, H.P. Análise Química em Plantas; Departamento de Química, ESALQ/USP: Piracicaba, Brazil, 1974; 56p. [Google Scholar]
- Braga, J.M.; Defelipo, B.V. Determinação espectrofotométrica do fósforo em extrato de solo e plantas. Rev. Ceres 1974, 1, 73–85. [Google Scholar]
- Alvarez, V.V.H.; Dias, L.E.; Ribeiro, E.S., Jr.; Fonseca, C.A. Métodos de Análises de Enxofre em Solos e Plantas; Editora UFV: Viçosa, Brazil, 2001; 131p. [Google Scholar]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatographymass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Cross, J.M.; von Korff, M.; Altmann, T.; Bartzetko, L.; Sulpice, R.; Gibon, Y.; Palacios, N.; Stitt, M. Variation of enzyme activities and metabolite levels in 24 Arabidopsis accessions growing in carbon-limited conditions. Plant Physiol. 2006, 142, 1574–1588. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fernie, A.R.; Roscher, A.; Ratcliffe, R.G.; Kruger, N.J. Fructose 2,6-bisphosphate activates pyrophosphate: Fructose-6-phosphate 1-phosphotransferase and increases triose phosphate to hexose phosphate cycling heterotrophic cells. Planta 2001, 212, 250–263. [Google Scholar] [CrossRef]
- Cuadros-Inostroza, A.; Caldana, C.; Redestig, H.; Kusano, M.; Lisec, J.; Peña-Cortés, H.; Willmitzer, L.; Hannah, M.A. (TargetSearch—A Bioconductor package for the efficient preprocessing of GC-MS metabolite profiling data. BMC Bioinform. 2009, 10, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernie, A.R.; Aharoni, A.; Willmitzer, L.; Stitt, M.; Tohge, T.; Kopka, J.; Caroll, A.J.; Saito, K.; Fraser, P.D.; Deluca, V. Recommendations for reporting metabolite data. Plant Cell 2011, 23, 2477–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, C.D. GENES—A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Nepomuceno, A.L.; Oosterhuis, D.M.; Stewart, J.M. Physiological responses of cotton leaves and roots to water deficit induced by polyethylene glycol. Environ. Exp. Bot. 1998, 40, 29–41. [Google Scholar] [CrossRef]
- Khalid, K.A.; da Silva, J.A.T.; Cai, W. Water deficit and polyethylene glycol 6000 affects morphological and biochemical characters of Pelargonium odoratissimum (L.). Sci. Hortic. 2010, 125, 159–166. [Google Scholar] [CrossRef]
- Ji, H.; Li, X. ABA mediates PEG-mediated premature differentiation of root apical meristem in plants. Plant Signal. Behav. 2014, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Basal, O.; Szabó, A.; Veres, S. Physiology of soybean as affected by PEG-induced drought stress. Curr. Plant Biol. 2020, 22, 100135. [Google Scholar] [CrossRef]
- Lawlor, D.W. Absorption of polyethylene glycols by plant and their effects on plant growth. New Phytol. 1970, 96, 501–513. [Google Scholar] [CrossRef]
- Slama, I.; Ghnaya, T.; Hessini, K.; Messedi, D.; Savouré, A.; Abdelly, C. Comparative study of the effects of mannitol and PEG osmotic stress on growth and solute accumulation in Sesuvium portulacastrum. Environ. Exp. Bot. 2007, 61, 10–17. [Google Scholar] [CrossRef]
- Condé, S.A.; Picoli, E.A.T.; Corrêa, T.R.; Dias, L.A.S.; Lourenço, R.D.S.; Silva, F.C.S.; Pereira, W.L.; Zauza, E.A.V. Biomarkers for early selection in eucalyptus tolerant to dieback associated with water deficit. Rev. Bras. Cienc. Agrar. 2020, 15, 7515. [Google Scholar] [CrossRef]
- Razouk, R.; Hssaini, L.; Alghoum, M.; Adiba, A.; Hamdani, A. Phenotyping Olive Cultivars for Drought Tolerance Using Leaf Macro-Characteristics. Horticulturae 2022, 8, 939. [Google Scholar] [CrossRef]
- Andrade-Bueno, I.G.; Picoli, E.A.T.; dos Santos Isaias, R.M.; Lopes-Mattos, K.L.B.; Cruz, C.D.; Kuki, K.N.; Zauza, E.A.V. Wood anatomy of field grown eucalypt genotypes exhibiting differential dieback and water deficit tolerance. Curr. Plant Biol. 2020, 22, 100186. [Google Scholar] [CrossRef]
- Condé, S.A.; Picoli, E.A.T.; Corrêa, T.R.; Lourenço, R.D.S. Marcadores anatômicos do pecíolo e a tolerância à seca de ponteiros e ao déficit hídrico em eucalipto. Nativa 2020, 8, 591–596. [Google Scholar] [CrossRef]
- Sala, A.; Piper, F.; Hoch, G. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol. 2010, 186, 274–281. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A Central Regulator of Plant Growth and Development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Fujiwara, T. Boron transport in plants: Co-ordinated regulation of transporters. Ann. Bot. 2010, 105, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Wadt, P.G.S.; Novais, R.F.; Alvarez, V.; Fonseca, S.; Barros, N.F. Valores de referência para macronutrientes em eucalipto obtidos pelos métodos DRIS e chance matemática. Rev. Bras. Ciênc. Solo 1998, 22, 685–692. [Google Scholar] [CrossRef]
- Kurihara, C.H.; Maeda, S.; Alvarez, V.H.V. Interpretação de Resultados de Análise Foliar; Embrapa: CPAO: Dourados, Brazil, 2005; 44p, ((Embrapa: CPAO. Documentos, 74), Colombo, PR, Brazil). [Google Scholar]
- Wadt, P.G.S.; Novais, R.F. Influência da idade da árvore na interpretação do estado nutricional de Eucalyptus grandis, pelos métodos do nível crítico e do DRIS. In Proceedings of the IUFRO Conference on Silviculture and Improvement for Eucalypt, EMBRAPA; Centro nacional de Pesquisa de Florestas, Colombo, Brazil; 1997; pp. 262–268. [Google Scholar]
- Herbert, M.A. Fertilizers and eucalypt plantations in South Africa. In Nutrition of Eucalypts; Attiwill, P.M., Adams, M.A., Eds.; CSIRO: Collingwood, Australia, 1996; pp. 303–325. [Google Scholar]
- Marschner, H. Mineral Nutrition in Higher Plants, 2nd ed.; Academic Press: London, UK, 1995; p. 889. [Google Scholar]
- Malavolta, E.; Vitti, G.C.; Oliveira, A.S. Avaliação do Estado Nutricional das Plantas: Princípios e Aplicações, 2nd ed.; Potafós: Piracicaba, Brazil, 1997; 319p. [Google Scholar]
- Sakya, A.T.; Dell, B.; Huan, L. Boron requirements for Eucalyptus globulus seedlings. Plant Soil 2002, 246, 87–95. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | SuzT | SuzMT | SuzTP | SuzS |
|---|---|---|---|---|
| N | 1.22 ± 0.06 ab | 1.13 ± 0.08 b | 1.14 ± 0.06 b | 1.50 ± 0.09 a |
| P | 0.23 ± 0.01 a | 0.24 ± 0.01 a | 0.29 ± 0.01 a | 0.24 ± 0.01 a |
| K | 1.07 ± 0.02 ab | 0.83 ± 0.03 b | 0.98 ± 0.05 ab | 1.13 ± 0.06 a |
| Ca | 1.08 ± 0.03 a | 0.88 ± 0.04 ab | 0.95 ± 0.02 ab | 0.85 ± 0.05 b |
| Mg | 0.39 ± 0.01 a | 0.31 ± 0.01 b | 0.39 ± 0.01 a | 0.35 ± 0.01 ab |
| Zn | 22.9 ± 1.42 b | 21.93 ± 0.99 b | 33.55 ± 1.16 a | 19.81 ± 0.87 b |
| Fe | 173.28 ± 11.94 a | 95.39 ± 14.44 a | 177.22 ± 21.95 a | 118.07 ± 7.37 a |
| Mn | 242.52 ± 23.78 a | 172.01 ± 23.86 a | 204.50 ± 6.34 a | 193.81 ± 8.31 a |
| B | 20.84 ± 0.54 b | 19.17 ± 1.45 b | 24.27 ± 1.60 ab | 26.61 ±1.69 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caetano-Madeira, D.D.; Omena-Garcia, R.P.; Elerati, T.L.; da Silva Lopes, C.B.; Corrêa, T.R.; de Souza, G.A.; Oliveira, L.A.; Cruz, C.D.; Bhering, L.L.; Nunes-Nesi, A.; et al. Metabolic, Nutritional and Morphophysiological Behavior of Eucalypt Genotypes Differing in Dieback Resistance in Field When Submitted to PEG-Induced Water Deficit. Agronomy 2023, 13, 1261. https://doi.org/10.3390/agronomy13051261
Caetano-Madeira DD, Omena-Garcia RP, Elerati TL, da Silva Lopes CB, Corrêa TR, de Souza GA, Oliveira LA, Cruz CD, Bhering LL, Nunes-Nesi A, et al. Metabolic, Nutritional and Morphophysiological Behavior of Eucalypt Genotypes Differing in Dieback Resistance in Field When Submitted to PEG-Induced Water Deficit. Agronomy. 2023; 13(5):1261. https://doi.org/10.3390/agronomy13051261
Chicago/Turabian StyleCaetano-Madeira, Débora Durso, Rebeca Patricia Omena-Garcia, Tamiris Lopes Elerati, Camila Batista da Silva Lopes, Thais Roseli Corrêa, Genaína Aparecida de Souza, Leonardo Araujo Oliveira, Cosme Damião Cruz, Leonardo Lopes Bhering, Adriano Nunes-Nesi, and et al. 2023. "Metabolic, Nutritional and Morphophysiological Behavior of Eucalypt Genotypes Differing in Dieback Resistance in Field When Submitted to PEG-Induced Water Deficit" Agronomy 13, no. 5: 1261. https://doi.org/10.3390/agronomy13051261