
Phylogenetic Analyses and Transcriptional Survey Reveal the Characteristics, Evolution, and Expression Profile of NBS-Type Resistance Genes in Papaya
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Similarity Comparison of NLRs
2.2. Duplication Modes Identification and Chromosomal Localization of Papaya NLRs
2.3. Phylogenetic Analysis of NLRs in Papaya
2.4. Analysis of Conserved Motifs, Domains, and Gene Structures of NLRs in Papaya
2.5. Estimation of Divergent Times
2.6. Micro-Synteny Analysis of NLRs among Species
2.7. Expression Pattern and Co-Expression Analysis of NLRs in Papaya
2.8. Real-Time Quantitative PCR Analysis of NLRs in PRSV Infected Papaya
3. Results
3.1. A Set of Complete and Simplified NLRs in Papaya
3.2. Different Similarity Distribution of Three NLR Subclasses
3.3. Chromosome Distribution and Duplication Modes of NLRs in Papaya
3.4. Conserved Sequences, Gene Structures, and Phylogeny of NLRs in Papaya
3.5. Inheritance- and Insertion-Origin of NLRs in Papaya Genome
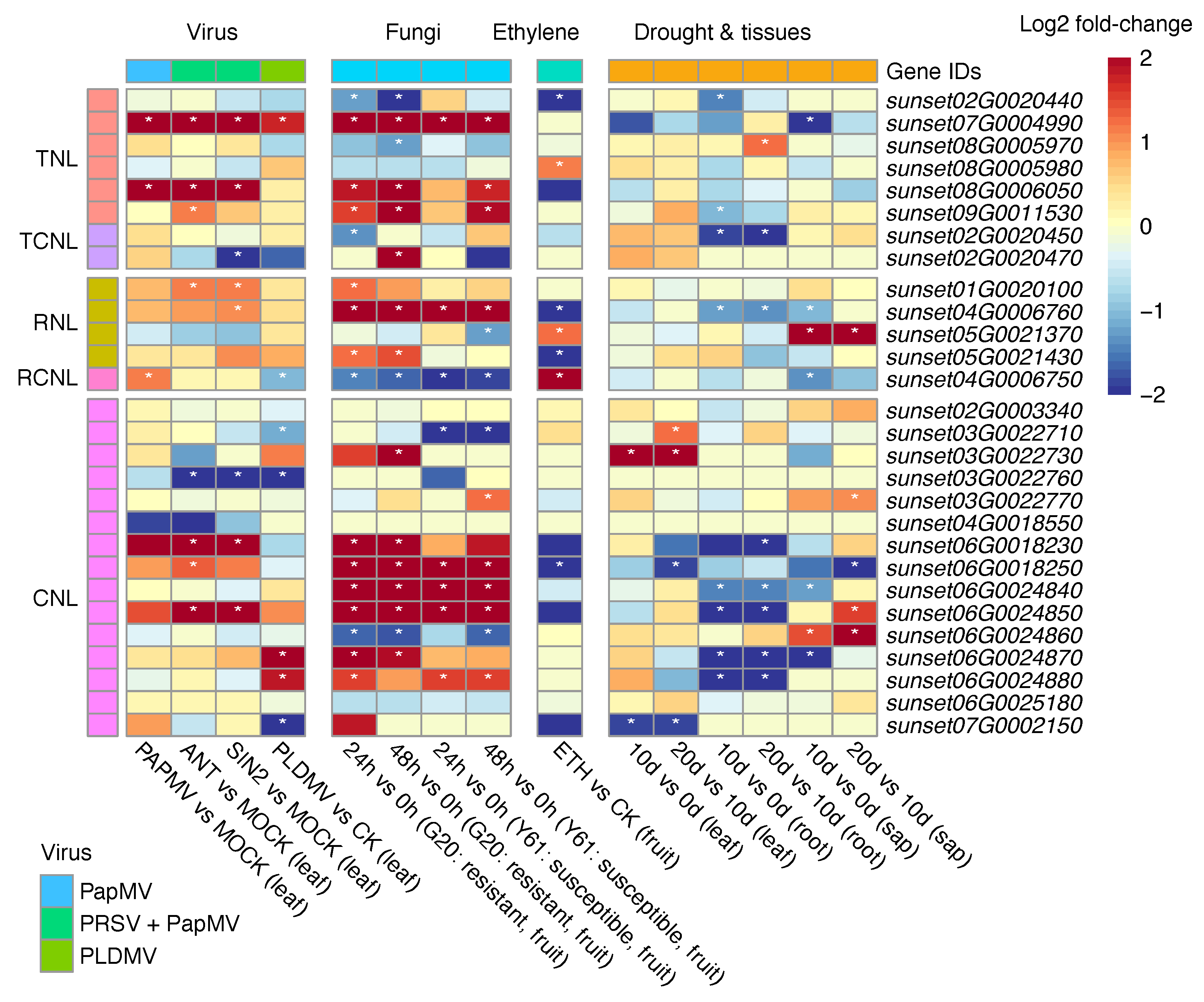
3.6. Expression of NLR Genes in Papaya under Biotic and Abiotic Stresses
3.7. Co-Expression Network Analysis of NLRs in Papaya Infected by Fungi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NLR | Nucleotide-binding site (NBS) leucine-rich repeat (LRR) receptor |
| ETI | Effector-triggered immunity |
| TIR | Toll/Interleukin-1 receptor |
| RPW8 | Resistance to Powdery Mildew Locus 8 |
| CC | Coiled-coil |
| Mya | Millions of years ago |
| Ks | Synonymous substitution rates |
| PRSV | Papaya ringspot virus |
| PAPMV | Papaya mosaic virus |
| PLDMV | Papaya leaf-distortion mosaic virus |
| WGCNA | Weighted gene co-expression network analysis |
| WGD | Whole-genome duplication |
| TD | Tandem duplication |
| PD | Proximal duplication |
| TRD | Transposed duplication |
| DSD | Dispersed duplication |
| PCA | Principal Component Analysis |
| FPKM | Fragments Per kb per Million reads |
| DEG | Differentially expressed genes |
Appendix A


References
- Ming, R.; Hou, S.; Feng, Y.; Yu, Q.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; VanBuren, R.; Liu, J.; Fang, J.; Zhang, X.; Liao, Z.; Wai, C.M.; Xu, X.; Chen, S.; Zhang, S.; et al. SunUp and Sunset genomes revealed impact of particle bombardment mediated transformation and domestication history in papaya. Nat. Genet. 2022, 54, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.H.; Lu, B.G.; Feng, L.; Yang, F.Y.; Geng, J.J.; Ming, R.; Chen, X.J. Isolation of ripening-related genes from ethylene/1-MCP treated papaya through RNA-seq. BMC Genom. 2017, 18, 671. [Google Scholar] [CrossRef]
- Chan-León, A.C.; Estrella-Maldonado, H.; Dubé, P.; Ortiz, G.F.; Espadas-Gil, F.; May, C.T.; Prado, J.R.; Desjardins, Y.; Santamaría, J.M. The high content of β-carotene present in orange-pulp fruits of Carica papaya L. is not correlated with a high expression of the CpLCY-β2 gene. Food Res. Int. 2017, 100, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sharma, A.; Niewiara, M.J.; Singh, R.; Ming, R.; Yu, Q. Papain-like cysteine proteases in Carica papaya: Lineage-specific gene duplication and expansion. BMC Genom. 2018, 19, 26. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, C.C.; Wang, Y.; Xu, J.; Wang, G.; Bai, X. Biological evaluations of decellularized extracellular matrix collagen microparticles prepared based on plant enzymes and aqueous two-phase method. Regen. Biomater. 2021, 8, rbab002. [Google Scholar] [CrossRef]
- Hafid, K.; John, J.; Sayah, T.M.; Domínguez, R.; Becila, S.; Lamri, M.; Dib, A.L.; Lorenzo, J.M.; Gagaoua, M. One-step recovery of latex papain from Carica Papaya using three phase partitioning and its use as milk-clotting and meat-tenderizing agent. Int. J. Biol. Macromol. 2020, 146, 798–810. [Google Scholar] [CrossRef]
- Lobato-Gómez, M.; Hewitt, S.; Capell, T.; Christou, P.; Dhingra, A.; Girón-Calva, P.S. Transgenic and genome-edited fruits: Background, constraints, benefits, and commercial opportunities. Hortic. Res. 2021, 8, 166. [Google Scholar] [CrossRef]
- Wu, Z.; Mo, C.; Zhang, S.; Li, H. Characterization of papaya ringspot virus isolates infecting transgenic papaya ‘Huanong No. 1’ in South China. Sci. Rep. 2018, 8, 8206. [Google Scholar] [CrossRef]
- Wang, W.; Feng, B.; Zhou, J.M.; Tang, D. Plant immune signaling: Advancing on two frontiers. J. Integr. Plant Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef]
- Andolfo, G.; Dohm, J.C.; Himmelbauer, H. Prediction of NB-LRR resistance genes based on full-length sequence homology. Plant J. Cell Mol. Biol. 2022, 110, 1592–1602. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Lolle, S.; Stevens, D.; Coaker, G. Plant NLR-triggered immunity: From receptor activation to downstream signaling. Curr. Opin. Immunol. 2020, 62, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Delaux, P.M.; Schornack, S. Plant evolution driven by interactions with symbiotic and pathogenic microbes. Science 2021, 371, eaba6605. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Xue, J.Y.; Wang, Q.; Wang, B.; Chen, J.Q. Revisiting the Origin of Plant NBS-LRR Genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, Z.; Zhang, Y.M.; Li, Q.; Jiang, X.M.; Jiang, Z.; Tang, J.H.; Chen, D.; Wang, Q.; Chen, J.Q.; et al. An angiosperm NLR Atlas reveals that NLR gene reduction is associated with ecological specialization and signal transduction component deletion. Mol. Plant 2021, 14, 2015–2031. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.Q.; Xue, J.Y.; Wu, P.; Zhang, Y.M.; Wu, Y.; Hang, Y.Y.; Wang, B.; Chen, J.Q. Large-Scale Analyses of Angiosperm Nucleotide-Binding Site-Leucine-Rich Repeat Genes Reveal Three Anciently Diverged Classes with Distinct Evolutionary Patterns. Plant Physiol. 2016, 170, 2095–2109. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Jiang, X.M.; Shao, Z.Q. Genome-Wide Analysis of NLR Disease Resistance Genes in an Updated Reference Genome of Barley. Front. Genet. 2021, 12, 694682. [Google Scholar] [CrossRef]
- Porter, B.W.; Paidi, M.; Ming, R.; Alam, M.; Nishijima, W.T.; Zhu, Y.J. Genome-wide analysis of Carica papaya reveals a small NBS resistance gene family. Mol. Genet. Genom. MGG 2009, 281, 609–626. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Heal, R.; Wyler, M.; Schmid, M.W.; Jones, J.D.G. Concerted expansion and contraction of immune receptor gene repertoires in plant genomes. Nat. Plants 2022, 8, 1146–1152. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, C.; Yang, H.; Kuang, R.; Huang, B.; Wei, Y. Identification of WRKY transcription factor genes in papaya and response of their expresion to Colletotrichum brevisporum infection. J. Northwest A F Univ. (Nat. Sci. Ed.) 2022, 50, 127–138+154. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, C.; Yang, H.; Kuang, R.; Liu, K.; Huang, B.; Wei, Y. Comparative transcriptomics and genomic analyses reveal differential gene expression related to Colletotrichum brevisporum resistance in papaya (Carica papaya L.). Front. Plant Sci. 2022, 13, 1038598. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Quan, X.; Jia, G.; Xiao, J.; Cloutier, S.; You, F.M. RGAugury: A pipeline for genome-wide prediction of resistance gene analogs (RGAs) in plants. BMC Genom. 2016, 17, 852. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Lupas, A.; Van Dyke, M.; Stock, J. Predicting coiled coils from protein sequences. Science 1991, 252, 1162–1164. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: An Expanded Resource for Species Divergence Times. Mol. Biol. Evol. 2022, 39, msac174. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.; Zhang, X.; Li, Z.; Zhao, Y.; Lohaus, R.; Chang, X.; Dong, W.; Ho, S.Y.W.; Liu, X.; et al. The water lily genome and the early evolution of flowering plants. Nature 2020, 577, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization-diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, W.; Zhang, T.; Gong, Z.; Zhao, H.; Han, G.Z. Out of Water: The Origin and Early Diversification of Plant R-Genes. Plant Physiol. 2018, 177, 82–89. [Google Scholar] [CrossRef]
- Nguyen, N.P.D.; Mirarab, S.; Kumar, K.; Warnow, T. Ultra-large alignments using phylogeny-aware profiles. Genome Biol. 2015, 16, 124. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PloS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Shimodaira, H.; Hasegawa, M. Multiple Comparisons of Log-Likelihoods with Applications to Phylogenetic Inference. Mol. Biol. Evol. 1999, 16, 1114. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, J.; Wu, J.; Zhang, H.; Liu, G.; Wang, X.; Dai, L. ParaAT: A parallel tool for constructing multiple protein-coding DNA alignments. Biochem. Biophys. Res. Commun. 2012, 419, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nielsen, R. Estimating synonymous and nonsynonymous substitution rates under realistic evolutionary models. Mol. Biol. Evol. 2000, 17, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, S.; Li, L.; Yang, T.; Dong, S.; Wei, T.; Wu, S.; Liu, Y.; Gong, Y.; Feng, X.; et al. The Cycas genome and the early evolution of seed plants. Nat. Plants 2022, 8, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Belfield, E.J.; Brown, C.; Ding, Z.J.; Chapman, L.; Luo, M.; Hinde, E.; Van Es, S.W.; Johnson, S.; Ning, Y.; Zheng, S.J.; et al. Thermal stress accelerates Arabidopsis thaliana mutation rate. Genome Res. 2021, 31, 40–50. [Google Scholar] [CrossRef]
- Vargas-Mejía, P.; Vega-Arreguín, J.; Chávez-Calvillo, G.; Ibarra-Laclette, E.; Silva-Rosales, L. Differential Accumulation of Innate- and Adaptive-Immune-Response-Derived Transcripts during Antagonism between Papaya Ringspot Virus and Papaya Mosaic Virus. Viruses 2020, 12, 230. [Google Scholar] [CrossRef]
- Gamboa-Tuz, S.D.; Pereira-Santana, A.; Zamora-Briseño, J.A.; Castano, E.; Espadas-Gil, F.; Ayala-Sumuano, J.T.; Keb-Llanes, M.Á.; Sanchez-Teyer, F.; Rodríguez-Zapata, L.C. Transcriptomics and co-expression networks reveal tissue-specific responses and regulatory hubs under mild and severe drought in papaya (Carica papaya L.). Sci. Rep. 2018, 8, 14539. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Anders, S.; McCarthy, D.J.; Chen, Y.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013, 8, 1765–1786. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Z.; Zhu, R.; Wang, F.; Cheng, Y.; Liu, Y. Three Differential Expression Analysis Methods for RNA Sequencing: Limma, EdgeR, DESeq2. J. Vis. Exp. 2021, 175, e62528. [Google Scholar] [CrossRef]
- Zhang, B.; Horvath, S. A general framework for weighted gene co-expression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Langfelder, P.; Luo, R.; Oldham, M.C.; Horvath, S. Is my network module preserved and reproducible? PLoS Comput. Biol. 2011, 7, e1001057. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sperry, A.O. Identification of a novel Leucine-rich repeat protein and candidate PP1 regulatory subunit expressed in developing spermatids. BMC Cell Biol. 2008, 9, 9. [Google Scholar] [CrossRef]
- DeVaul, N.; Wang, R.; Sperry, A.O. PPP1R42, a PP1 binding protein, regulates centrosome dynamics in ARPE-19 cells. Biol. Cell 2013, 105, 359–371. [Google Scholar] [CrossRef]
- Ma, S.; Lapin, D.; Liu, L.; Sun, Y.; Song, W.; Zhang, X.; Logemann, E.; Yu, D.; Wang, J.; Jirschitzka, J.; et al. Direct pathogen-induced assembly of an NLR immune receptor complex to form a holoenzyme. Science 2020, 370, eabe3069. [Google Scholar] [CrossRef]
- Antunes Carvalho, F.; Renner, S.S. A dated phylogeny of the papaya family (Caricaceae) reveals the crop’s closest relatives and the family’s biogeographic history. Mol. Phylogenetics Evol. 2012, 65, 46–53. [Google Scholar] [CrossRef]
- Rockinger, A.; Sousa, A.; Carvalho, F.A.; Renner, S.S. Chromosome number reduction in the sister clade of Carica Papaya with concomitant genome size doubling. Am. J. Bot. 2016, 103, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef]
- Wang, J.; Song, W.; Chai, J. Structure, biochemical function, and signaling mechanism of plant NLRs. Mol. Plant 2023, 16, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Steuernagel, B.; Witek, K.; Krattinger, S.G.; Ramirez-Gonzalez, R.H.; Schoonbeek, H.J.; Yu, G.; Baggs, E.; Witek, A.I.; Yadav, I.; Krasileva, K.V.; et al. The NLR-Annotator Tool Enables Annotation of the Intracellular Immune Receptor Repertoire. Plant Physiol. 2020, 183, 468–482. [Google Scholar] [CrossRef]
- Toda, N.; Rustenholz, C.; Baud, A.; Le Paslier, M.C.; Amselem, J.; Merdinoglu, D.; Faivre-Rampant, P. NLGenomeSweeper: A Tool for Genome-Wide NBS-LRR Resistance Gene Identification. Genes 2020, 11, 333. [Google Scholar] [CrossRef]
- Friis, E.M.; Pedersen, K.R.; Crane, P.R. The emergence of core eudicots: New floral evidence from the earliest Late Cretaceous. Proc. Biol. Sci. 2016, 283, 20161325. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Fang, D.; Sahu, S.K.; Yang, S.; Guang, X.; Folk, R.; Smith, S.A.; Chanderbali, A.S.; Chen, S.; Liu, M.; et al. Chloranthus genome provides insights into the early diversification of angiosperms. Nat. Commun. 2021, 12, 6930. [Google Scholar] [CrossRef]
- Li, Z.; Defoort, J.; Tasdighian, S.; Maere, S.; Van de Peer, Y.; De Smet, R. Gene Duplicability of Core Genes Is Highly Consistent across All Angiosperms. Plant Cell 2016, 28, 326–344. [Google Scholar] [CrossRef]
- Glenfield, C.; Innan, H. Gene Duplication and Gene Fusion Are Important Drivers of Tumourigenesis during Cancer Evolution. Genes 2021, 12, 1376. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Yang, F.S.; Nie, S.; Liu, H.; Shi, T.L.; Tian, X.C.; Zhou, S.S.; Bao, Y.T.; Jia, K.H.; Guo, J.F.; Zhao, W.; et al. Chromosome-level genome assembly of a parent species of widely cultivated azaleas. Nat. Commun. 2020, 11, 5269. [Google Scholar] [CrossRef]
- Wang, K.; de la Torre, D.; Robertson, W.E.; Chin, J.W. Programmed chromosome fission and fusion enable precise large-scale genome rearrangement and assembly. Science 2019, 365, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Tamborski, J.; Krasileva, K.V. Evolution of Plant NLRs: From Natural History to Precise Modifications. Annu. Rev. Plant Biol. 2020, 71, 355–378. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hirayama, C.; Nakamura, M.; Tateishi, K.; Tamura, Y.; Hattori, M.; Kohno, K. Papain protects papaya trees from herbivorous insects: Role of cysteine proteases in latex. Plant J. Cell Mol. Biol. 2004, 37, 370–378. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
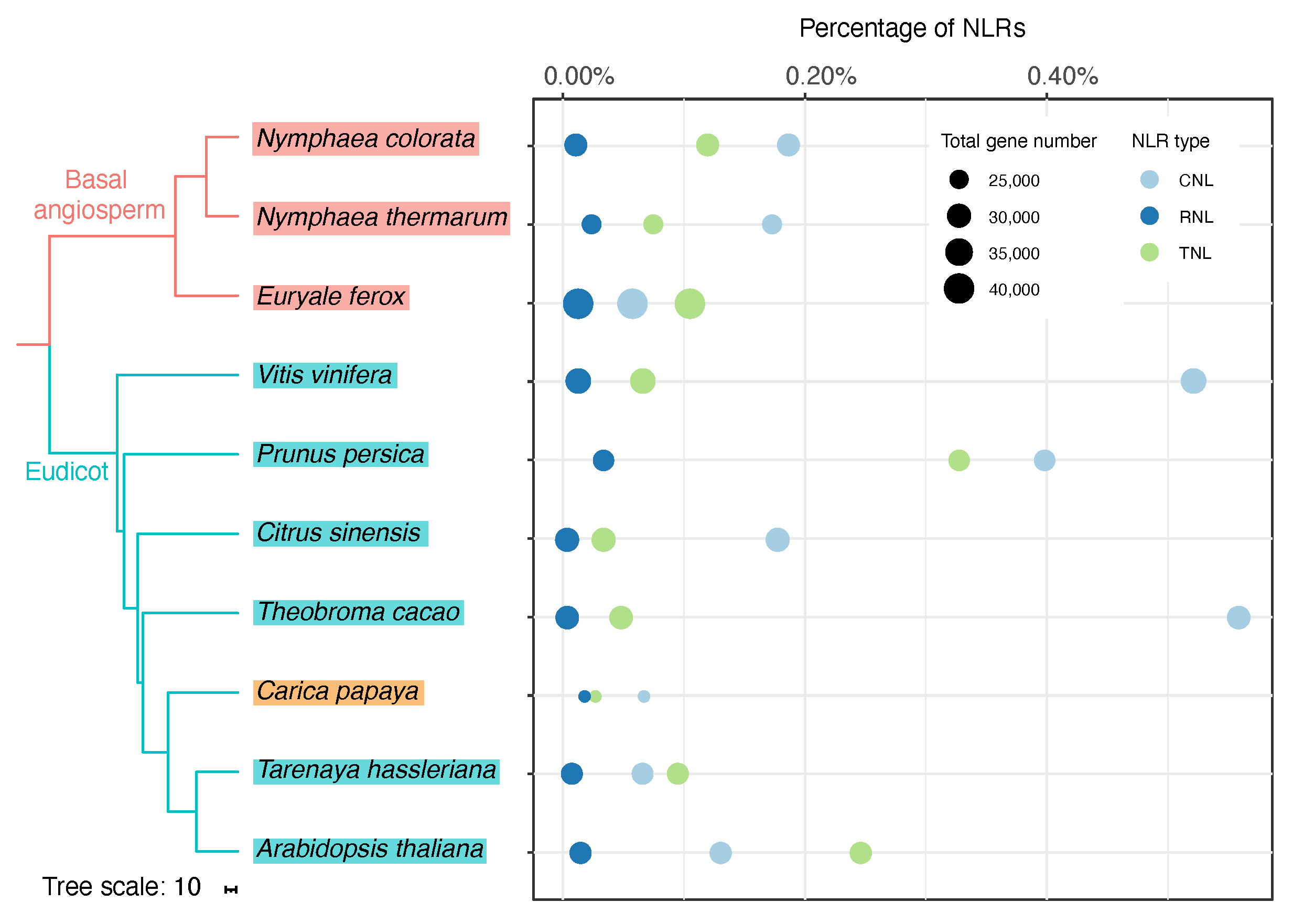
| Species | Total | NLR | NBS | CN | TN | RN | NL | TCN | RCN | CNL | TNL | RNL | TCNL | RCNL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nymphaea colorata | 28,438 | 255 | 63 | 17 | 12 | 2 | 57 | 0 | 1 | 53 | 34 | 3 | 9 | 4 |
| Nymphaea thermarum | 25,461 | 157 | 21 | 15 | 10 | 2 | 31 | 1 | 1 | 44 | 19 | 6 | 3 | 4 |
| Euryale ferox | 40,049 | 134 | 8 | 9 | 8 | 0 | 13 | 1 | 0 | 23 | 42 | 5 | 16 | 9 |
| Vitis vinifera | 31,845 | 439 | 52 | 41 | 8 | 2 | 126 | 0 | 1 | 166 | 21 | 4 | 7 | 11 |
| Prunus persica | 26,873 | 374 | 24 | 7 | 17 | 0 | 105 | 3 | 1 | 107 | 88 | 9 | 11 | 2 |
| Citrus sinensis | 24,456 | 112 | 7 | 12 | 2 | 0 | 25 | 0 | 1 | 53 | 10 | 1 | 0 | 1 |
| Theobroma cacao | 29,181 | 275 | 15 | 13 | 3 | 0 | 63 | 0 | 0 | 163 | 14 | 1 | 0 | 3 |
| Carica papaya | 22,416 | 59 | 10 | 3 | 1 | 0 | 16 | 1 | 0 | 15 | 6 | 4 | 2 | 1 |
| Tarenaya hassleriana | 27,396 | 76 | 7 | 1 | 4 | 0 | 15 | 0 | 0 | 18 | 26 | 2 | 3 | 0 |
| Arabidopsis thaliana | 27,628 | 166 | 4 | 1 | 14 | 1 | 25 | 3 | 1 | 36 | 68 | 4 | 9 | 0 |
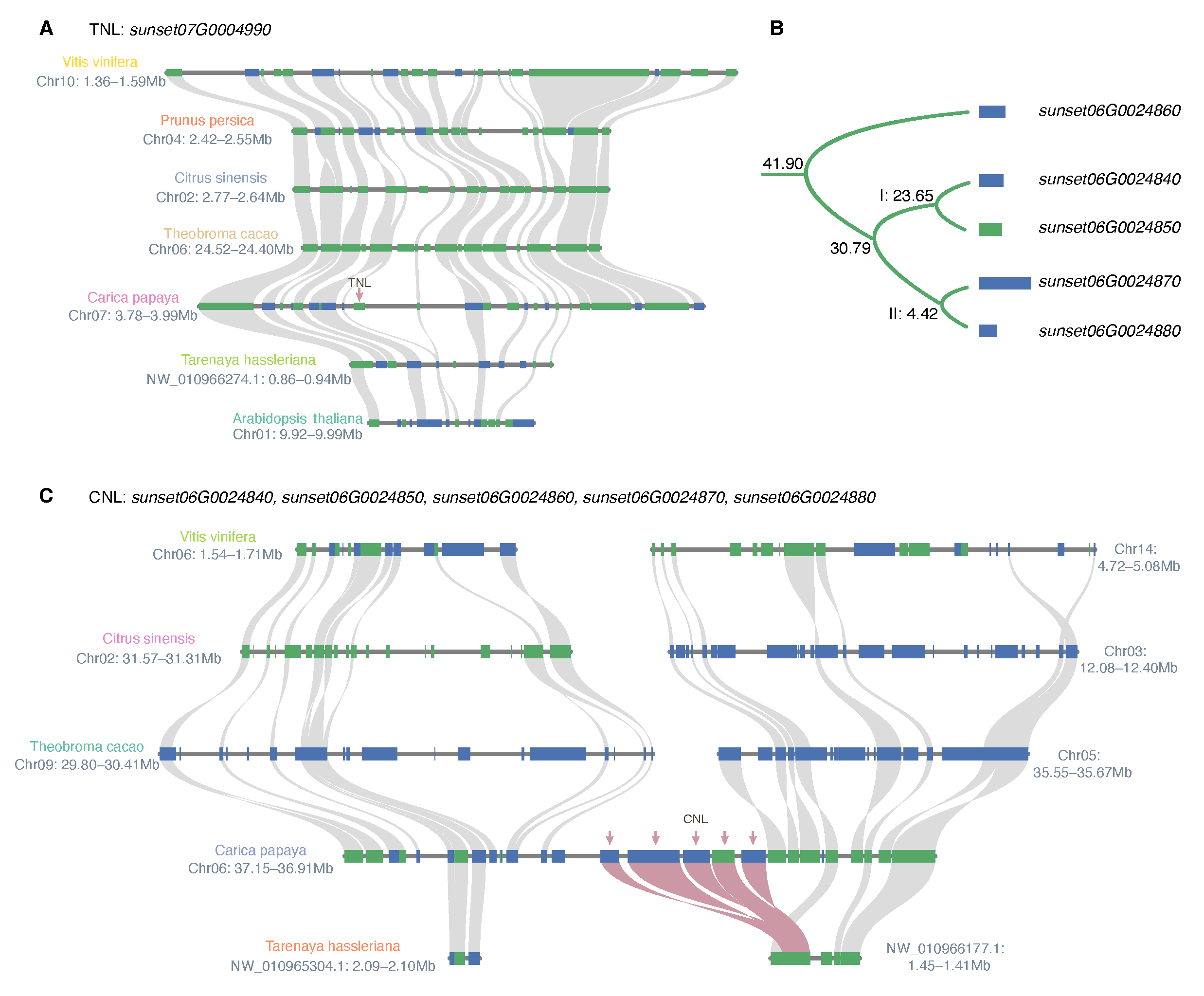
| Duplicate 1 | Type 1 | Duplicate 2 | Type 2 | E-Value | Duplication Mode | Ks | Divergent Time (Mya) |
|---|---|---|---|---|---|---|---|
| sunset06G0024860 | CNL | sunset06G0024870 | CNL | tandem | 1.01 | 41.90 | |
| sunset06G0024840 | CNL | sunset06G0024860 | CNL | 0 | dispersed | 0.93 | 38.81 |
| sunset06G0024860 | CNL | sunset06G0024880 | CNL | dispersed | 0.87 | 36.19 | |
| sunset06G0024850 | CNL | sunset06G0024860 | CNL | tandem | 0.87 | 36.06 | |
| sunset06G0024850 | CNL | sunset06G0024870 | CNL | 0 | dispersed | 0.74 | 30.79 |
| sunset06G0024840 | CNL | sunset06G0024850 | CNL | tandem | 0.57 | 23.65 | |
| sunset06G0024870 | CNL | sunset06G0024880 | CNL | 0 | tandem | 0.11 | 4.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Wang, Y.; Xiong, A.; Zhao, H.; Jia, R.; Li, M.; An, H.; Ji, C.; Guo, A. Phylogenetic Analyses and Transcriptional Survey Reveal the Characteristics, Evolution, and Expression Profile of NBS-Type Resistance Genes in Papaya. Agronomy 2023, 13, 970. https://doi.org/10.3390/agronomy13040970
Jiang Q, Wang Y, Xiong A, Zhao H, Jia R, Li M, An H, Ji C, Guo A. Phylogenetic Analyses and Transcriptional Survey Reveal the Characteristics, Evolution, and Expression Profile of NBS-Type Resistance Genes in Papaya. Agronomy. 2023; 13(4):970. https://doi.org/10.3390/agronomy13040970
Chicago/Turabian StyleJiang, Qian, Yu Wang, Aisheng Xiong, Hui Zhao, Ruizong Jia, Mengyao Li, Huaming An, Changmian Ji, and Anping Guo. 2023. "Phylogenetic Analyses and Transcriptional Survey Reveal the Characteristics, Evolution, and Expression Profile of NBS-Type Resistance Genes in Papaya" Agronomy 13, no. 4: 970. https://doi.org/10.3390/agronomy13040970