
Identifying QTLs Related to Grain Filling Using a Doubled Haploid Rice (Oryza sativa L.) Population
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Phenotypic Measurements
2.3. DNA Extraction and Molecular Markers Analysis
2.4. Linkage Mapping and QTL Analysis
2.5. Gene Ontology Search of qFG3, qFG5-1, and qFG5-2-Related Genes
3. Results
3.1. Differential Phenotypic Response of Parental Lines and Doubled Haploid Lines
3.2. Detected QTLs Associated with Grain Filling and Other Traits
3.3. Genes Harbored by qFG3, qFG5-1 and qFG5-2 QTLs Associated with Grain Filling in Rice
4. Discussion
4.1. Differential Grain-Filling Ratio between Parents and DH Lines
4.2. A Novel QTL Associated with Grain Filling Is Identified
4.3. QTL qFG3 Harbors Genes with Transcription Factor Activity
4.4. qFG3-Related Genes Are Associated with Abiotic Stress Response Mechanisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.H.; Dar, Z.A.; Dar, S.A. Breeding strategies for improving rice yield—A review. Agric. Sci. 2015, 6, 467. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Khush, G.S.; Virk, P.; Tang, Q.; Zou, Y. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 2008, 108, 32–38. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef]
- Okamura, M.; Arai-Sanoh, Y.; Yoshida, H.; Mukouyama, T.; Adachi, S.; Yabe, S.; Nakagawa, H.; Tsutsumi, K.; Taniguchi, Y.; Kobayashi, N. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crops Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- Zha, X.; Luo, X.; Qian, X.; He, G.; Yang, M.; Li, Y.; Yang, J. Over-expression of the rice LRK1 gene improves quantitative yield components. Plant Biotechnol. J. 2009, 7, 611–620. [Google Scholar] [CrossRef]
- Rao, Y.; Li, Y.; Qian, Q. Recent progress on molecular breeding of rice in China. Plant Cell Rep. 2014, 33, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, L.; Wang, M.; Xu, Y.Y.; Luo, W.; Liu, Y.J.; Xu, Z.H.; Li, J.; Chong, K. Engineering OsBAK1 gene as a molecular tool to improve rice architecture for high yield. Plant Biotechnol. J. 2009, 7, 791–806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cao, Z.; Zhou, Q.; Chen, J.; Xu, G.; Gu, J.; Liu, L.; Wang, Z.; Yang, J.; Zhang, H. Grain filling characteristics and their relations with endogenous hormones in large-and small-grain mutants of rice. PLoS ONE 2016, 11, e0165321. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J. Grain-filling problem in ‘super’rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Xie, H.; Wang, Y.; Lü, B.; Liang, J. The effect of sucrose and abscisic acid interaction on sucrose synthase and its relationship to grain filling of rice (Oryza sativa L.). J. Exp. Bot. 2009, 60, 2641–2652. [Google Scholar] [CrossRef] [Green Version]
- Su, J.-C. Starch synthesis and grain filling in rice. In Developments in Crop Science; Elsevier: Amsterdam, The Netherlands, 2000; Volume 26, pp. 107–124. [Google Scholar]
- Jiang, Z.; Chen, Q.; Chen, L.; Yang, H.; Zhu, M.; Ding, Y.; Li, W.; Liu, Z.; Jiang, Y.; Li, G. Efficiency of sucrose to starch metabolism is related to the initiation of inferior grain filling in large panicle rice. Front. Plant Sci. 2021, 12, 732867. [Google Scholar] [CrossRef] [PubMed]
- Yabe, S.; Yoshida, H.; Kajiya-Kanegae, H.; Yamasaki, M.; Iwata, H.; Ebana, K.; Hayashi, T.; Nakagawa, H. Description of grain weight distribution leading to genomic selection for grain-filling characteristics in rice. PLoS ONE 2018, 13, e0207627. [Google Scholar] [CrossRef] [Green Version]
- Isshiki, M.; Nakajima, M.; Satoh, H.; Shimamoto, K. dull: Rice mutants with tissue-specific effects on the splicing of the waxy pre-mRNA. Plant J. 2000, 23, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Ise, K.; Akama, Y.; Horisue, N.; Nakane, A.; Yokoo, M.; Ando, I.; Hata, T.; Suto, M.; Numaguchi, K.; Nemoto, H. Milky Queen, a new high-quality rice cultivar with low amylose content in endosperm. Bull. Natl. Inst. Crop Sci. 2001, 2, 39–61. [Google Scholar]
- He, P.; Li, S.; Qian, Q.; Ma, Y.; Li, J.; Wang, W.; Chen, Y.; Zhu, L. Genetic analysis of rice grain quality. Theor. Appl. Genet. 1999, 98, 502–508. [Google Scholar] [CrossRef]
- Han, Y.; Xu, M.; Liu, X.; Yan, C.; Korban, S.S.; Chen, X.; Gu, M. Genes coding for starch branching enzymes are major contributors to starch viscosity characteristics in waxy rice (Oryza sativa L.). Plant Sci. 2004, 166, 357–364. [Google Scholar] [CrossRef]
- Butardo, V.M., Jr.; Daygon, V.D.; Colgrave, M.L.; Campbell, P.M.; Resurreccion, A.; Cuevas, R.P.; Jobling, S.A.; Tetlow, I.; Rahman, S.; Morell, M. Biomolecular analyses of starch and starch granule proteins in the high-amylose rice mutant Goami 2. J. Agric. Food Chem. 2012, 60, 11576–11585. [Google Scholar] [CrossRef]
- Bao, J.; Corke, H.; Sun, M. Microsatellites, single nucleotide polymorphisms and a sequence tagged site in starch-synthesizing genes in relation to starch physicochemical properties in nonwaxy rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Aluko, G.; Martinez, C.; Tohme, J.; Castano, C.; Bergman, C.; Oard, J. QTL mapping of grain quality traits from the interspecific cross Oryza sativa × O. glaberrima. Theor. Appl. Genet. 2004, 109, 630–639. [Google Scholar] [CrossRef]
- Chen, L.; Deng, Y.; Zhu, H.; Hu, Y.; Jiang, Z.; Tang, S.; Wang, S.; Ding, Y. The initiation of inferior grain filling is affected by sugar translocation efficiency in large panicle rice. Rice 2019, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Akita, S. Improving yield potential in tropical rice. In Progress in Irrigated Rice Research; International Rice Research Institute: Manila, Philippines, 1989; pp. 41–73. [Google Scholar]
- Nagata, K.; Fukuta, Y.; Shimizu, H.; Yagi, T.; Terao, T. Quantitative trait loci for sink size and ripening traits in rice (Oryza sativa L.). Breed. Sci. 2002, 52, 259–273. [Google Scholar] [CrossRef] [Green Version]
- San, N.S.; Ootsuki, Y.; Adachi, S.; Yamamoto, T.; Ueda, T.; Tanabata, T.; Motobayashi, T.; Ookawa, T.; Hirasawa, T. A near-isogenic rice line carrying a QTL for larger leaf inclination angle yields heavier biomass and grain. Field Crops Res. 2018, 219, 131–138. [Google Scholar] [CrossRef]
- Yamamoto, T.; Suzuki, T.; Suzuki, K.; Adachi, S.; Sun, J.; Yano, M.; Ookawa, T.; Hirasawa, T. Characterization of a genomic region that maintains chlorophyll and nitrogen contents during ripening in a high-yielding stay-green rice cultivar. Field Crops Res. 2017, 206, 54–64. [Google Scholar] [CrossRef]
- Zhang, G.-H.; Li, S.-Y.; Wang, L.; Ye, W.-J.; Zeng, D.-L.; Rao, Y.-C.; Peng, Y.-L.; Hu, J.; Yang, Y.-L.; Xu, J. LSCHL4 from japonica cultivar, which is allelic to NAL1, increases yield of indica super rice 93-11. Mol. Plant 2014, 7, 1350–1364. [Google Scholar] [CrossRef] [Green Version]
- Ha, W.; Kim, H.; Choi, H.; Lim, S.; Seo, H.S.; Lim, M. Heritability Estimates of Sink and Source Characters by Fin-fill5 Correlation in Rice. Korean J. Crop Sci. 2002, 47, 151–156. [Google Scholar]
- Murray, M.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, K.-S.; Baek, J.; Cho, Y.-i.; Jeong, Y.-M.; Lee, Y.-Y.; Oh, J.; Won, Y.J.; Kang, D.-Y.; Oh, H.; Kim, S.L. Single nucleotide polymorphism (SNP) discovery and kompetitive allele-specific PCR (KASP) marker development with Korean japonica rice varieties. Plant Breed. Biotechnol. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Seo, J.; Lee, G.; Jin, Z.; Kim, B.; Chin, J.H.; Koh, H.-J. Development and application of indica–japonica SNP assays using the Fluidigm platform for rice genetic analysis and molecular breeding. Mol. Breed. 2020, 40, 39. [Google Scholar] [CrossRef] [Green Version]
- Akond, Z.; Alam, M.J.; Hasan, M.N.; Uddin, M.S.; Alam, M.; Mollah, M.N.H. A comparison on some interval mapping approaches for QTL detection. Bioinformation 2019, 15, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.-W.; Kabange, N.R.; Phyo, Z.; Park, S.-Y.; Lee, S.-M.; Lee, J.-Y.; Shin, D.; Cho, J.H.; Park, D.-S.; Ko, J.-M. Combined linkage mapping and genome-wide association study identified QTLs associated with grain shape and weight in rice (Oryza sativa L.). Agronomy 2020, 10, 1532. [Google Scholar] [CrossRef]
- Zhu, X.; Liang, S.; Yin, J.; Yuan, C.; Wang, J.; Li, W.; He, M.; Wang, J.; Chen, W.; Ma, B. The DnaJ OsDjA7/8 is essential for chloroplast development in rice (Oryza sativa). Gene 2015, 574, 11–19. [Google Scholar] [CrossRef]
- Zhong, X.; Yang, J.; Shi, Y.; Wang, X.; Wang, G.L. The DnaJ protein OsDjA6 negatively regulates rice innate immunity to the blast fungus Magnaporthe oryzae. Mol. Plant Pathol. 2018, 19, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, N.; Ranjan, R.; Parida, S.K.; Agarwal, P.; Tyagi, A.K. Mediator subunit OsMED14_1 plays an important role in rice development. Plant J. 2020, 101, 1411–1429. [Google Scholar] [CrossRef]
- Lin, Z.; Li, X.; Shannon, L.M.; Yeh, C.-T.; Wang, M.L.; Bai, G.; Peng, Z.; Li, J.; Trick, H.N.; Clemente, T.E. Parallel domestication of the Shattering1 genes in cereals. Nat. Genet. 2012, 44, 720–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Hu, Y.; Zhao, Y.; Liu, H.; Zhou, D.-X. A WUSCHEL-LIKE HOMEOBOX gene represses a YABBY gene expression required for rice leaf development. Plant Physiol. 2007, 144, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, H.; Tang, J.; Li, Z.; Li, Z.; Chen, D.; Lin, W. A proteomic study on molecular mechanism of poor grain-filling of rice (Oryza sativa L.) inferior spikelets. PLoS ONE 2014, 9, e89140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Wang, H.; Bi, Y.; Yan, Y.; Gao, Y.; Xiong, X.; Wang, J.; Li, D.; Song, F. ONAC066, a stress-responsive NAC transcription activator, positively contributes to rice immunity against Magnaprothe oryzae through modulating expression of OsWRKY62 and three cytochrome P450 genes. Front. Plant Sci. 2021, 12, 749186. [Google Scholar] [CrossRef]
- Yang, W.; Gao, M.; Yin, X.; Liu, J.; Xu, Y.; Zeng, L.; Li, Q.; Zhang, S.; Wang, J.; Zhang, X. Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome p450. Mol. Plant 2013, 6, 1945–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, A.; Qian, Q.; Yin, H.; Liu, X.; Yin, C.; Lan, Y.; Tang, J.; Tang, Z.; Cao, S.; Wang, X. EUI1, encoding a putative cytochrome P450 monooxygenase, regulates internode elongation by modulating gibberellin responses in rice. Plant Cell Physiol. 2006, 47, 181–191. [Google Scholar] [CrossRef]
- Guo, F.; Endo, M.; Yamaguchi, T.; Uchino, A.; Sunohara, Y.; Matsumoto, H.; Iwakami, S. Investigation of clomazone-tolerance mechanism in a long-grain cultivar of rice. Pest Manag. Sci. 2021, 77, 2454–2461. [Google Scholar] [CrossRef] [PubMed]
- Fang, N.; Xu, R.; Huang, L.; Zhang, B.; Duan, P.; Li, N.; Luo, Y.; Li, Y. SMALL GRAIN 11 controls grain size, grain number and grain yield in rice. Rice 2016, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, G.; Tian, Y.; Song, Y.; Liang, W.; Zhang, D. A rice glutamyl-tRNA synthetase modulates early anther cell division and patterning. Plant Physiol. 2018, 177, 728–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, W.-L.; Chung, Y.-L.; Fang, J.-C.; Lu, C.-A. Novel interaction between CCR4 and CAF1 in rice CCR4–NOT deadenylase complex. Plant Mol. Biol. 2017, 93, 79–96. [Google Scholar] [CrossRef]
- Ren, D.; Rao, Y.; Yu, H.; Xu, Q.; Cui, Y.; Xia, S.; Yu, X.; Liu, H.; Hu, H.; Xue, D. MORE FLORET1 encodes a MYB transcription factor that regulates spikelet development in rice. Plant Physiol. 2020, 184, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.-L.; Chu, R.-Z.; Zhang, Y.; Luo, J.; Su, Y.-Y.; Xie, L.-J.; Zhang, H.-S.; Wang, J.-F.; Bao, Y.-M. OsJAMyb, a R2R3-type MYB transcription factor, enhanced blast resistance in transgenic rice. Physiol. Mol. Plant Pathol. 2015, 92, 154–160. [Google Scholar] [CrossRef]
- Bao, Y.-M.; Sun, S.-J.; Li, M.; Li, L.; Cao, W.-L.; Luo, J.; Tang, H.-J.; Huang, J.; Wang, Z.-F.; Wang, J.-F. Overexpression of the Qc-SNARE gene OsSYP71 enhances tolerance to oxidative stress and resistance to rice blast in rice (Oryza sativa L.). Gene 2012, 504, 238–244. [Google Scholar] [CrossRef]
- Sikdar, M.; Kim, J. Isolation of a gene encoding 3-isopropylmalate dehydrogenase from rice. Russ. J. Plant Physiol. 2011, 58, 190–196. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, S.; Qian, Q.; Zeng, D.; Zhang, M.; Guo, L.; Liu, X.; Zhang, B.; Deng, L.; Liu, X. BC10, a DUF266-containing and Golgi-located type II membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.). Plant J. 2009, 57, 446–462. [Google Scholar] [CrossRef]
- Li, K.; Chen, Y.; Luo, Y.; Huang, F.; Zhao, C.; Cheng, F.; Xiang, X.; Pan, G. A 22-bp deletion in OsPLS3 gene encoding a DUF266-containing protein is implicated in rice leaf senescence. Plant Mol. Biol. 2018, 98, 19–32. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.-F.; Liang, S.; Jones, R.L.; Lu, Y.-T. Molecular and biochemical characterization of a calcium/calmodulin-binding protein kinase from rice. Biochem. J. 2002, 368, 145–157. [Google Scholar] [CrossRef]
- Koo, S.C.; Choi, M.S.; Chun, H.J.; Shin, D.B.; Park, B.S.; Kim, Y.H.; Park, H.-M.; Seo, H.S.; Song, J.T.; Kang, K.Y. The calmodulin-binding transcription factor OsCBT suppresses defense responses to pathogens in rice. Mol. Cells 2009, 27, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Lee, S.H.; Kim, J.K.; Chun, H.J.; Choi, M.S.; Chung, W.S.; Moon, B.C.; Kang, C.H.; Park, C.Y.; Yoo, J.H. Mlo, a modulator of plant defense and cell death, is a novel calmodulin-binding protein: Isolation and characterization of a rice Mlo homologue. J. Biol. Chem. 2002, 277, 19304–19314. [Google Scholar] [CrossRef] [Green Version]
- Ricachenevsky, F.K.; Sperotto, R.A.; Menguer, P.K.; Fett, J.P. Identification of Fe-excess-induced genes in rice shoots reveals a WRKY transcription factor responsive to Fe, drought and senescence. Mol. Biol. Rep. 2010, 37, 3735–3745. [Google Scholar] [CrossRef]
- Fu, S.; Fu, L.; Zhang, X.; Huang, J.; Yang, G.; Wang, Z.; Liu, Y.-G.; Zhang, G.; Wu, D.; Xia, J. OsC2DP, a novel C2 domain-containing protein is required for salt tolerance in rice. Plant Cell Physiol. 2019, 60, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, S.; Qin, J.; Sun, C.; Liu, F. The lipid transfer protein Os LTPL 159 is involved in cold tolerance at the early seedling stage in rice. Plant Biotechnol. J. 2020, 18, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, W.; Lu, Z.; Ouyang, Y.; Yao, J. A lipid transfer protein, OsLTPL36, is essential for seed development and seed quality in rice. Plant Sci. 2015, 239, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, D.; Zhang, H.; Wang, X.; Zheng, Z.; Song, F. Molecular characterization of rice OsBIANK1, encoding a plasma membrane-anchored ankyrin repeat protein, and its inducible expression in defense responses. Mol. Biol. Rep. 2010, 37, 653–660. [Google Scholar] [CrossRef]
- Pooja, S.; Sweta, K.; Mohanapriya, A.; Sudandiradoss, C.; Siva, R.; Gothandam, K.M.; Babu, S. Homotypic clustering of OsMYB4 binding site motifs in promoters of the rice genome and cellular-level implications on sheath blight disease resistance. Gene 2015, 561, 209–218. [Google Scholar] [CrossRef]
- Takai, T.; Fukuta, Y.; Shiraiwa, T.; Horie, T. Time-related mapping of quantitative trait loci controlling grain-filling in rice (Oryza sativa L.). J. Exp. Bot. 2005, 56, 2107–2118. [Google Scholar] [CrossRef] [Green Version]
- Sekhar, S.; Kumar, J.; Mohanty, S.; Mohanty, N.; Panda, R.S.; Das, S.; Shaw, B.P.; Behera, L. Identification of novel QTLs for grain fertility and associated traits to decipher poor grain filling of basal spikelets in dense panicle rice. Sci. Rep. 2021, 11, 13617. [Google Scholar] [CrossRef] [PubMed]
- Song, X.-J.; Huang, W.; Shi, M.; Zhu, M.-Z.; Lin, H.-X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Huang, J.; Lan, H.; Wang, C.; Yin, C.; Wu, Y.; Tang, H.; Qian, Q.; Li, J. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc. Natl. Acad. Sci. USA 2012, 109, 21534–21539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Sun, X.; Chen, Y.; Huang, Y.; Tong, C.; Bao, J. Rapid identification of major QTLs associated with rice grain weight and their utilization. PLoS ONE 2015, 10, e0122206. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Jiao, G.; Lin, H.; Sheng, Z.; Shao, G.; Xie, L.; Tang, S.; Xu, Q.; Hu, P. GRAIN INCOMPLETE FILLING 2 regulates grain filling and starch synthesis during rice caryopsis development. J. Integr. Plant Biol. 2017, 59, 134–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Fu, Y.; Zhu, Z.; Tan, L.; Liu, F.; Sun, X.; Sun, X.; Sun, C. GS 6, a member of the GRAS gene family, negatively regulates grain size in rice. J. Integr. Plant Biol. 2013, 55, 938–949. [Google Scholar] [CrossRef]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kanegae, H.; Konishi, S.; Yano, M. Deletion in a gene associated with grain size increased yields during rice domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef]
- Qi, P.; Lin, Y.-S.; Song, X.-J.; Shen, J.-B.; Huang, W.; Shan, J.-X.; Zhu, M.-Z.; Jiang, L.; Gao, J.-P.; Lin, H.-X. The novel quantitative trait locus GL3. 1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res. 2012, 22, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Sun, S.; Yao, J.; Wang, C.; Yu, S.; Xu, C.; Li, X.; Zhang, Q. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 19579–19584. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fan, C.; Xing, Y.; Jiang, Y.; Luo, L.; Sun, L.; Shao, D.; Xu, C.; Li, X.; Xiao, J. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, A.; Liu, X.; Chen, J. Grain size associated genes and the molecular regulatory mechanism in rice. Int. J. Mol. Sci. 2022, 23, 3169. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.-I.; Onishi, A. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef]
- Breviario, D.; Gianì, S.; Morello, L. Multiple tubulins: Evolutionary aspects and biological implications. Plant J. 2013, 75, 202–218. [Google Scholar] [CrossRef]
- Wasteneys, G.O. Microtubule organization in the green kingdom: Chaos or self-order? J. Cell Sci. 2002, 115, 1345–1354. [Google Scholar] [CrossRef]
- Mathur, J.; Hülskamp, M. Microtubules and microfilaments in cell morphogenesis in higher plants. Curr. Biol. 2002, 12, R669–R676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T. Microtubules in plants. Arab. Book Am. Soc. Plant Biol. 2015, 13, e0179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segami, S.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Small and round seed 5 gene encodes alpha-tubulin regulating seed cell elongation in rice. Rice 2012, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Jan, A.; Komatsu, S. Functional analysis of OsTUB8, an anther-specific β-tubulin in rice. Plant Sci. 2007, 172, 832–838. [Google Scholar] [CrossRef]
- Blume, Y.B.; Lloyd, C.W.; Yemets, A.I. Plant tubulin phosphorylation and its role in cell cycle progression. In The Plant Cytoskeleton: A Key Tool for Agro-Biotechnology; Springer: Dordrecht, The Netherlands, 2008; pp. 145–159. [Google Scholar]
- Chen, J.; Cao, F.; Li, H.; Shan, S.; Tao, Z.; Lei, T.; Liu, Y.; Xiao, Z.; Zou, Y.; Huang, M. Genotypic variation in the grain photosynthetic contribution to grain filling in rice. J. Plant Physiol. 2020, 253, 153269. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.K.; Kitching, R.; Vary, C.; Kola, I.; Seth, A. Isolation of target gene promoter/enhancer sequences by whole genome PCR method. In Transcription Factor Protocols; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–11. [Google Scholar]
- Tanaka, W.; Toriba, T.; Hirano, H.Y. Three TOB 1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice. New Phytol. 2017, 215, 825–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasid, S.; Simontacchi, M.; Bartoli, C.G.; Puntarulo, S. Chloroplasts as a nitric oxide cellular source. Effect of reactive nitrogen species on chloroplastic lipids and proteins. Plant Physiol. 2006, 142, 1246–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, K.-J.; Turkan, I.; Krieger-Liszkay, A. Redox-and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacza, Z.; Pankotai, E.; Busija, D.W. Mitochondrial nitric oxide synthase: Current concepts and controversies. Front. Biosci. 2009, 14, 4436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandalio, L.; Romero-Puertas, M. Peroxisomes sense and respond to environmental cues by regulating ROS and RNS signalling networks. Ann. Bot. 2015, 116, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Hayashi, M.; Mano, S.; Nishimura, M.; Barroso, J.B. Peroxisomes are required for in vivo nitric oxide accumulation in the cytosol following salinity stress of Arabidopsis plants. Plant Physiol. 2009, 151, 2083–2094. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Traits | Cultivation Seasons | |||||||
|---|---|---|---|---|---|---|---|---|
| Early 2018 | Normal 2019 | Normal 2021 | Late 2018 | |||||
| 93-11 | Milyang 352 | 93-11 | Milyang 352 | 93-11 | Milyang 352 | 93-11 | Milyang 352 | |
| Panicle number per plant | 14 ± 1.5 * | 10 ± 2.6 | 11 ± 0.3 ns | 10 ± 1.1 | 8 ± 1.8 ns | 9 ± 0.7 | 12 ± 0.2 ns | 11 ± 0.9 |
| Spikelet number per panicle | 173 ± 6.2 ** | 120 ± 3.5 | 119 ± 2.6 ** | 100 ± 4.4 | 157 ± 4.0 ** | 220 ± 5.3 | 136 ± 3.6 ** | 173 ± 3.5 |
| Grain filling (%) | 86.1 ± 0.4 ** | 57.5 ± 1.7 | 93.4 ± 2.6 ** | 78.0 ± 8.9 | 91.7 ± 8.2 ** | 66.1 ± 8.7 | 74.4 ± 7.2 ** | 61.1 ± 4.6 |
| Thousand-grain weight (g) | 19.6 ± 0.9 ** | 23.1 ± 0.9 | 25.2 ± 1.0 ** | 23.2 ± 1.2 | 22.7 ± 1.1 ** | 24.2 ± 1.6 | 20.8 ± 0.8 ns | 23.3 ± 1.7 |
| Traits | Cultivation Seasons | ||||
|---|---|---|---|---|---|
| Early 2018 | Normal 2019 | Normal 2021 | Late 2018 | ||
| Panicle number per plant | Average ± SD | 13 ± 2.3 ab | 12 ± 2.3 b | 10 ± 1.8 a | 13 ± 2.3 c |
| Range | 9–22 | 7–20 | 6–16 | 8–21 | |
| Spikelet number per panicle | Average ± SD | 183 ± 40.2 a | 144 ± 28.9 c | 173 ± 31.4 c | 149 ± 26.4 c |
| Range | 92–283 | 22–216 | 105–273 | 96–220 | |
| Grain filling (%) | Average ± SD | 58.7 ± 16.7 b | 75.3 ± 13.9 b | 62.4 ± 16.4 b | 50.8 ± 26.5 c |
| Range | 4.7–86.1 | 15.7–94.1 | 10.8–91.7 | 1.0–77.6 | |
| Thousand-grain weight (g) | Average ± SD | 21.7 ± 2.2 b | 24.4 ± 2.5 ab | 23.3 ± 2.4 ab | 21.7 ± 2.7 ab |
| Range | 18.1–26.9 | 19.1–31.0 | 19.0–29.4 | 16.7–28.9 | |
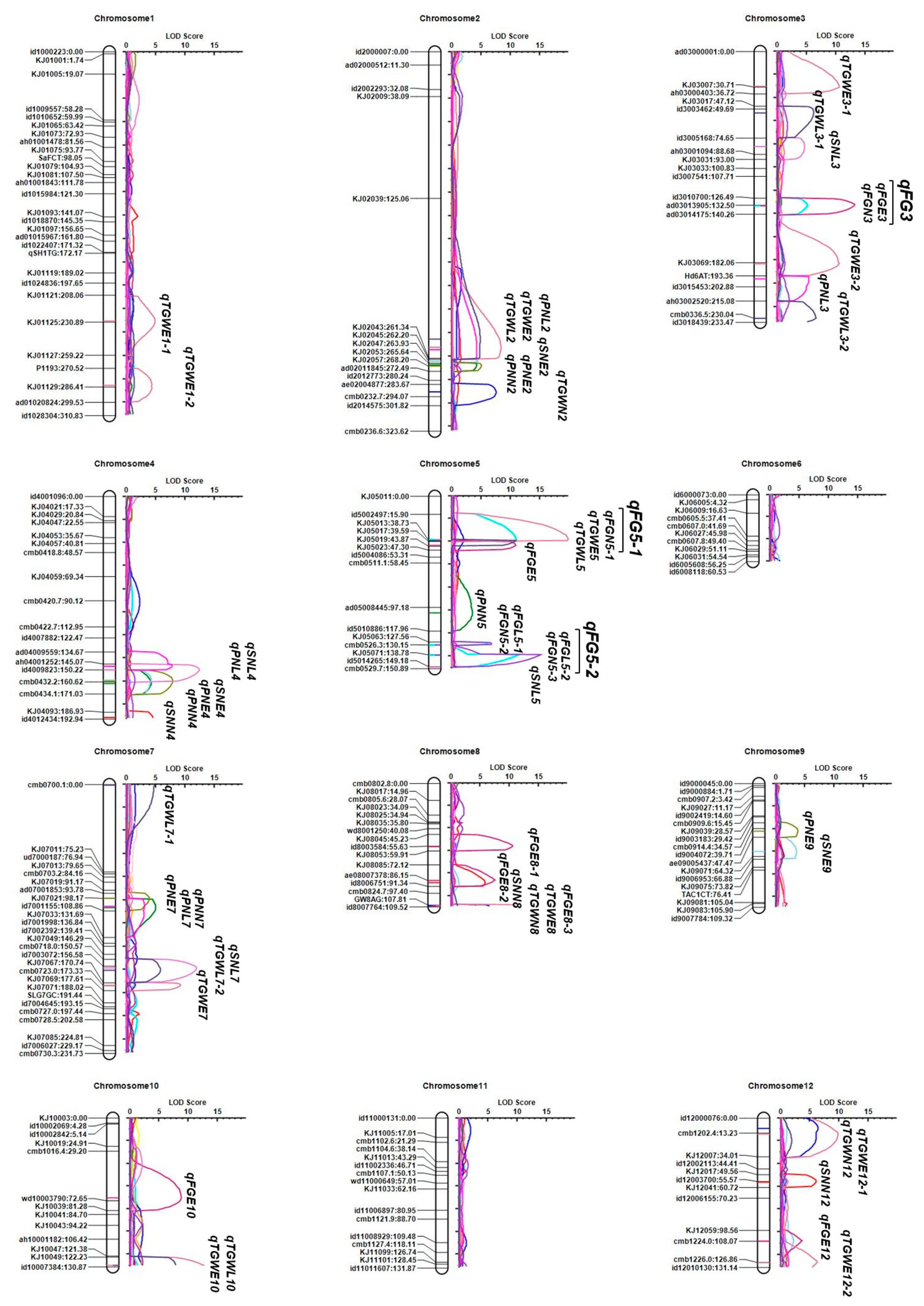
| Cultivation Season | Trait | QTL Name | Position (cM) | Left Marker | Right Marker | LOD | PVE (%) | Add | Left CI | Right CI |
|---|---|---|---|---|---|---|---|---|---|---|
| Early Season | Spikelet Number Per Panicle | qSNE2 | 264 | KJ02_047 | KJ02_053 | 4.50 | 14.08 | −13.75 | 262.5 | 265.5 |
| qSNE4 | 159 | id4009823 | cmb0432.2 | 4.23 | 14.43 | −14.86 | 152.5 | 170.5 | ||
| qSNE9 | 60 | ae09005437 | KJ09_071 | 3.52 | 12.73 | −13.56 | 51.5 | 66.5 | ||
| Panicle Number Per Plant | qPNE2 | 268 | KJ02_053 | KJ02_057 | 5.05 | 12.25 | 0.76 | 265.5 | 272.5 | |
| qPNE4 | 160 | id4009823 | cmb0432.2 | 7.87 | 20.67 | 1.06 | 154.5 | 167.5 | ||
| qPNE7 | 98 | ad07001853 | KJ07_021 | 3.95 | 9.39 | −0.66 | 93.5 | 105.5 | ||
| qPNE9 | 42 | id9004072 | ae09005437 | 3.72 | 9.12 | 0.72 | 36.5 | 47.5 | ||
| Percentage of Filled Grain | qFGE3 | 133 | ad03013905 | ad03014175 | 13.24 | 15.95 | 8.14 | 129.5 | 135.5 | |
| qFGE5 | 43 | KJ05_017 | KJ05_019 | 10.88 | 12.53 | 7.23 | 39.5 | 45.5 | ||
| qFGE8-1 | 56 | id8003584 | KJ08_053 | 10.45 | 11.82 | 7.02 | 52.5 | 58.5 | ||
| qFGE8-2 | 86 | KJ08_085 | ae08007378 | 7.44 | 7.86 | −5.79 | 82.5 | 88.5 | ||
| qFGE8-3 | 109 | GW8-AG | id8007764 | 4.17 | 4.18 | −4.25 | 107.5 | 109 | ||
| qFGE10 | 70 | cmb1016.4 | wd10003790 | 8.72 | 10.42 | 7.00 | 58.5 | 75.5 | ||
| qFGE12 | 108 | KJ12_059 | cmb1224.0 | 3.65 | 3.57 | −3.81 | 103.5 | 112.5 | ||
| Thousand Grain Weight | qTGWE1-1 | 230 | KJ01_121 | KJ01_125 | 4.90 | 3.17 | −0.63 | 221.5 | 238.5 | |
| qTGWE1-2 | 285 | P1193 | KJ01_129 | 4.32 | 2.77 | −0.44 | 275.5 | 285.5 | ||
| qTGWE2 | 252 | KJ02_039 | KJ02_043 | 8.37 | 8.12 | −0.75 | 238.5 | 262.5 | ||
| qTGWE3-1 | 30 | ad03000001 | KJ03_007 | 10.60 | 7.41 | 0.71 | 22.5 | 32.5 | ||
| qTGWE3-2 | 183 | KJ03_069 | Hd6-AT | 10.58 | 7.64 | −0.73 | 172.5 | 187.5 | ||
| qTGWE5 | 38 | id5002497 | KJ05_013 | 19.80 | 16.94 | −1.08 | 32.5 | 39.5 | ||
| qTGWE7 | 173 | KJ07_067 | cmb0723.0 | 9.24 | 6.25 | 0.68 | 171.5 | 175.5 | ||
| qTGWE8 | 109 | GW8-AG | id8007764 | 10.88 | 7.58 | 0.74 | 107.5 | 109 | ||
| qTGWE10 | 130 | KJ10_049 | id10007384 | 12.66 | 9.33 | −0.79 | 127.5 | 130 | ||
| qTGWE12-1 | 14 | cmb1202.4 | KJ12_007 | 9.75 | 6.69 | 0.67 | 9.5 | 23.5 | ||
| qTGWE12-2 | 127 | cmb1226.0 | id12010130 | 6.26 | 3.95 | −0.52 | 122.5 | 131 | ||
| Normal Season (2019) | Spikelet Number Per Panicle | qSNN4 | 192 | KJ04_093 | id4012434 | 4.51 | 10.72 | −12.05 | 186.5 | 192 |
| qSNN8 | 88 | ae08007378 | id8006751 | 5.77 | 14.29 | −13.20 | 82.5 | 91.5 | ||
| qSNN12 | 56 | id12003700 | KJ12_041 | 6.09 | 14.87 | −13.73 | 51.5 | 60.5 | ||
| Panicle Number Per Plant | qPNN2 | 267 | KJ02_053 | KJ02_057 | 4.22 | 11.83 | 0.73 | 265.5 | 272.5 | |
| qPNN4 | 162 | cmb0432.2 | cmb0434.1 | 4.25 | 12.26 | 0.79 | 154.5 | 169.5 | ||
| qPNN5 | 102 | ad05008445 | id5010886 | 3.52 | 11.47 | −0.77 | 82.5 | 112.5 | ||
| qPNN7 | 106 | KJ07_021 | id7001155 | 4.99 | 14.68 | −0.81 | 98.5 | 114.5 | ||
| Percentage of Filled Grain | qFGN3 | 133 | ad03013905 | ad03014175 | 5.17 | 5.97 | 5.11 | 128.5 | 138.5 | |
| qFGN5-1 | 38 | id5002497 | KJ05_013 | 11.05 | 14.40 | 7.86 | 31.5 | 39.5 | ||
| qFGN5-2 | 130 | KJ05_063 | cmb0526.3 | 6.33 | 7.41 | −6.95 | 127.5 | 130.5 | ||
| qFGN5-3 | 139 | KJ05_071 | id5014265 | 11.48 | 15.05 | 9.90 | 138.5 | 140.5 | ||
| Thousand Grain Weight | qTGWN2 | 290 | ae02004877 | cmb0232.7 | 7.53 | 15.18 | −1.22 | 284.5 | 296.5 | |
| qTGWN8 | 109 | GW8-AG | id8007764 | 6.55 | 12.48 | 1.10 | 107.5 | 109 | ||
| qTGWN12 | 9 | id12000076 | cmb1202.4 | 6.33 | 12.92 | 1.08 | 2.5 | 13.5 | ||
| Late Season | Spikelet Number Per Panicle | qSNL3 | 82 | id3005168 | ah03001094 | 4.67 | 8.07 | −8.16 | 74.5 | 91.5 |
| qSNL4 | 150 | ah04001252 | id4009823 | 12.51 | 23.17 | −14.62 | 146.5 | 154.5 | ||
| qSNL5 | 150 | id5014265 | cmb0529.7 | 3.29 | 5.07 | −7.17 | 149.5 | 150 | ||
| qSNL7 | 158 | id7003072 | KJ07_067 | 11.93 | 22.70 | −14.07 | 154.5 | 163.5 | ||
| Panicle Number Per Plant | qPNL2 | 254 | KJ02_039 | KJ02_043 | 4.26 | 13.21 | 0.83 | 227.5 | 262.5 | |
| qPNL3 | 196 | Hd6-AT | id3015453 | 5.42 | 12.65 | 0.83 | 193.5 | 209.5 | ||
| qPNL4 | 147 | ah04001252 | id4009823 | 7.81 | 19.81 | 1.09 | 140.5 | 149.5 | ||
| qPNL7 | 105 | KJ07_021 | id7001155 | 3.07 | 7.19 | −0.61 | 98.5 | 108.5 | ||
| Percentage of Filled Grain | qFGL5-1 | 130 | KJ05_063 | cmb0526.3 | 6.79 | 8.08 | −9.99 | 127.5 | 130.5 | |
| qFGL5-2 | 139 | KJ05_071 | id5014265 | 15.16 | 21.66 | 16.38 | 138.5 | 141.5 | ||
| Thousand Grain Weight | qTGWL2 | 245 | KJ02_039 | KJ02_043 | 4.83 | 12.30 | −0.98 | 221.5 | 262.5 | |
| qTGWL3-1 | 53 | id3003462 | id3005168 | 6.19 | 8.76 | 0.83 | 47.5 | 63.5 | ||
| qTGWL3-2 | 230 | ah03002520 | cmb0336.5 | 6.59 | 8.57 | −0.82 | 221.5 | 233 | ||
| qTGWL5 | 39 | KJ05_013 | KJ05_017 | 10.89 | 15.44 | −1.10 | 38.5 | 41.5 | ||
| qTGWL7-1 | 0 | cmb0700.1 | KJ07_011 | 4.72 | 5.91 | −0.68 | 0.0 | 21.5 | ||
| qTGWL7-2 | 160 | id7003072 | KJ07_067 | 5.80 | 8.36 | 0.84 | 152.5 | 166.5 | ||
| qTGWL10 | 130 | KJ10_049 | id10007384 | 7.84 | 10.66 | −0.91 | 123.5 | 130 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-M.; Kabange, N.R.; Kang, J.-W.; Kwon, Y.; Cha, J.-K.; Park, H.; Oh, K.-W.; Seo, J.; Koh, H.-J.; Lee, J.-H. Identifying QTLs Related to Grain Filling Using a Doubled Haploid Rice (Oryza sativa L.) Population. Agronomy 2023, 13, 912. https://doi.org/10.3390/agronomy13030912
Lee S-M, Kabange NR, Kang J-W, Kwon Y, Cha J-K, Park H, Oh K-W, Seo J, Koh H-J, Lee J-H. Identifying QTLs Related to Grain Filling Using a Doubled Haploid Rice (Oryza sativa L.) Population. Agronomy. 2023; 13(3):912. https://doi.org/10.3390/agronomy13030912
Chicago/Turabian StyleLee, So-Myeong, Nkulu Rolly Kabange, Ju-Won Kang, Youngho Kwon, Jin-Kyung Cha, Hyeonjin Park, Ki-Won Oh, Jeonghwan Seo, Hee-Jong Koh, and Jong-Hee Lee. 2023. "Identifying QTLs Related to Grain Filling Using a Doubled Haploid Rice (Oryza sativa L.) Population" Agronomy 13, no. 3: 912. https://doi.org/10.3390/agronomy13030912