

Evaluation of Ethanedinitrile as a Potential Fumigant against Plasmodiophora brassicae, the Clubroot Pathogen

1
Department of Plant Protection, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Kamýcká 129, 165 00 Praha-Suchdol, Czech Republic
2
Lučební Závody Draslovka a.s., Havlíčkova 605, Kolín IV, 280 99 Kolín, Czech Republic
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(4), 1177; https://doi.org/10.3390/agronomy13041177
Submission received: 14 March 2023
/
Revised: 14 April 2023
/
Accepted: 19 April 2023
/
Published: 21 April 2023
(This article belongs to the Topic Integrated Pest Management of Crops)
Abstract
:Clubroot, caused by Plasmodiophora brassicae Woronin, is one of the most challenging diseases to control in oilseed rape (Brassica napus L.) and other crucifers globally. Ethanedinitrile (EDN) is a promising fumigant shown to be effective against various pests and soil-borne pathogens. However, its efficacy against P. brassicae is yet to be tested. Therefore, the current study is focused on evaluating the effectiveness of EDN against P. brassicae at different rates, applied for different durations of exposure. Clubroot-susceptible oilseed rape plants, grown in fumigated soil, were evaluated for disease severity and growth parameters in greenhouse conditions. EDN exhibited good efficacy against the pathogen, with 81.39% disease suppression compared to the control. Complete control of the disease was achieved at the rates of 42 g/m3 and 50 g/m3 as well as in the treatment variant of 35 g/m3-48 h. In addition, EDN improved plant health by increasing shoot weight by 58.24%. However, the combination of a prolonged fumigation period and higher rates resulted in a decline in the seedling emergence rate and plant shoot weight, indicating a potential adverse residual effect of higher rates and longer durations of exposure in plants.
1. Introduction
Clubroot is a major soil-borne disease of the crops belonging to the Brassicaceae family. Plasmodiophora brassicae Woronin, an obligate biotrophic protist, causes the disease [1]. The classic symptoms of this disease include swelling or gall formation in the infected roots, resulting from hypertrophy and hyperplasia in the root tissues. These galls interfere with water and nutrient uptake in the infected plants, eventually leading to reduced yield or premature death of the plants in cases of severe infection [2]. P. brassicae has been reported in over 60 countries worldwide, causing an estimated global production loss of 10–15% in oilseed rape, one of its most economically significant hosts [3].
The control of this disease is considered to be a challenging task due to the long persistence of resting spores in the soil and the continued evolution of new pathotypes, leading to the altered distribution, frequency, and virulence of existing pathotypes in infested fields [1,4]. Currently, there are no economically viable and completely effective control measures against the pathogen. However, an integrated management approach, including the use of resistant cultivars and improved cultural practices, as well as the use of chemicals and soil amendments, is usually adopted to curtail the spread of the disease and minimize the losses. Chemical control is integral to this integrated management process in heavily infested fields. A wide range of chemicals, including various soil fungicides as well as soil fumigants, have been assessed for efficacy against this pathogen so far. Fungicides such as cyazofamid, fluazinam, AG3 phosphonate, and pentachloronitrobenzene (PCNB) are effective against the pathogen [5,6,7,8]. However, the efficacy of these fungicides have been inconsistent and non-cost-effective, since a large volume of water is needed for the application of these fungicides by the soil drenching method [9]. Moreover, due to various health and environmental risks, cyazofamid and fluazinam are not approved in Europe for use against clubroot disease, while PCNB has been restricted in Canada [9,10].
On the other hand, soil fumigants such as metam sodium and dazomet have shown more consistent results than soil fungicides in controlling the disease [11,12]. However, there have been certain limitations regarding the consistency of efficacy under varied environmental factors, environmental and health concerns, and cost efficiency [13,14]. Therefore, screening new potential products for chemical control against the pathogen has become crucial in such a situation, as it is complicated to control the disease solely through the use of resistant cultivars or cultural practices in heavily infested fields.
Ethanedinitrile (EDN; C2N2) is a relatively new cyanogen-based fumigant initially used in stored grain products and timber as an ozone-safe alternative to methyl bromide [15,16]. It has since shown promising potential in control of a wide range of pests, weeds and nematodes, as well as soil-borne pathogens such as Fusarium oxysporum, Phytophthora cactorum, and Rhizoctonia spp. [17,18,19,20,21,22]. EDN is a colorless, pungent gas which penetrates target material very quickly due to its low density and high vapor pressure [23,24,25]. Moreover, Rodrigues et al. reported only 1% residue in pine logs after fumigation, thereby suggesting that EDN is relatively safe for the environment [26]. Furthermore, the degradation products of EDN include ammonium, oxalic acid, and hydrogen cyanide (HCN), which occur naturally in the environment and, in some cases, exhibit fertilizing effects as well [27,28]. These favorable physical and environmental attributes of EDN make it a promising potential candidate for the control of P. brassicae. Therefore, the objectives of the current research were to evaluate the efficacy of different rates of EDN application, for different durations of exposure, to P. brassicae infestation in a controlled environment, as well as their impact on growth parameters in oilseed rape.
2. Materials and Methods
2.1. Inoculum Preparation
The soil was collected from different clubroot-infested fields across the Moravian region of the Czech Republic where oilseed rape was grown previously. Fresh clubroot galls were prepared by planting Chinese cabbage (B. rapa L. ssp. pekinensis) ‘Granaat’ seedlings in the collected soil. Root galls were harvested after six weeks and 50g of root galls was collected randomly to prepare inoculum consisting of mixed pathotypes. The protocol proposed by Tewari et al. was used to prepare inoculum suspension with some modifications [29]. First, 50 g of root galls was mixed with 250 mL of sterile distilled water and homogenized with the help of an electric mixer and grinder. The homogenate was then filtered through six layers of cheesecloth. The resting spores were quantified with the help of a haemocytometer (VWR, Mississauga, ON, Canada). Finally, the inoculum concentration was adjusted to 107 spores/mL using sterile distilled water.
2.2. Soil Preparation and Treatment
Sandy loam-textured soil was collected from the experimental field of the Czech University of Life Sciences Prague (CZU) and used for the experiment. The soil was roughly sieved, sterilized in an autoclave, and later inoculated with P. brassicae by mixing the soil thoroughly with the previously prepared inoculum with spore concentration of 107 spores/mL. The soil was then transferred to small net pots of volume 150 mL lined with mesh cloth. A makeshift pouch was prepared from the mesh cloth in the pots and tied on the top to hold the soil inside it. A total of three hundred pots were prepared following this method. The soil in the pots was then subjected to treatment by fumigation with EDN in twenty-four variants at the concentrations of 12 g/m3, 20 g/m3, 27 g/m3, 35 g/m3, 42 g/m3, and 50 g/m3 for the periods of 12 h, 24 h, 36 h, and 48 h (Table 1). The fumigation of the soil was conducted at the fumigation chamber facility of Draslovka Holding a.s., located in the Czech Republic. Each variant of treatment consisted of ten pots. A separate batch of pots was kept aside, with no fumigation treatment to be used as the control. After the fumigation, the pouches in the pots were opened, and the soil was left for air ventilation for fourteen days. The pots were then transferred to bigger self-watering pots of size 40 cm × 19 cm × 18 cm. Each self-watering pot consisted of five net pots marked with their respective treatment variant name.
Seeds of the clubroot-susceptible winter rape cultivar ‘Inspiration’ were planted in pots and grown for six weeks in the greenhouse facility of CZU. Each treatment consisted of ten replicates in two self-watering pots. The plants were maintained at 20–22 °C and 16-h photoperiod. Seedling emergence was evaluated after three weeks of sowing. The plants were harvested after six weeks and analyzed for the presence of galls, degree of infection, and fresh weight of leaf and root.
2.3. Disease Assessment
Disease severity was graded on a scale of 0–3, where 0 represents no galls, 1 represents a few small galls, 2 represents moderate galls, and 3 represents severe galls [30]. The severity ratings for each experimental unit were converted to an index of disease (Disease Severity Index or DSI) using the following formula proposed by Strelkov et al. [31]:
where ∑ is the total sum, n represents the number of plants in each class, N represents the total number of plants, and 0, 1, 2, and 3 are the symptom severity classes (scores).
Subsequently, Percent Disease Control was calculated using DSI in the following formula:
2.4. Statistical Analysis
Statistical analysis was performed with SPSS Statistics for Mac, Version 26.0 (IBM SPSS Statistics for Mac, Version 26.0. Armonk, NY, USA: IBM Corp). Square root transformation was applied to the root weight and shoot weight data, and a mixed model analysis of variance (ANOVA) was performed to analyze the treatment effects and their interaction effects on the root weight and shoot weight of the plants. The impact of treatment within an individual EDN concentration group was analyzed using one-way ANOVA for each group. In addition, Tukey’s HSD post hoc test was performed for multiple comparisons.
A logarithmic transformation was applied to the disease severity data, and a Generalized Estimated Equation (GEE) was used to analyze the overall effect of treatment on disease severity. Kruskal–Wallis tests were performed on each concentration group to analyze the significant effects of treatment within individual groups across different periods of exposure. Post hoc analysis was conducted using the Dunn–Bonferroni test.
Differences were deemed significant at p < 0.05 for all the analyses. Normality was tested on the residuals, and the homogeneity of data variance was checked by Levene’s test. Additionally, Spearman’s rank correlation coefficients were calculated to analyze the relationship of treatment with the percent disease control (%) and seedling emergence rate (%).
3. Results
3.1. Effect on Disease Severity
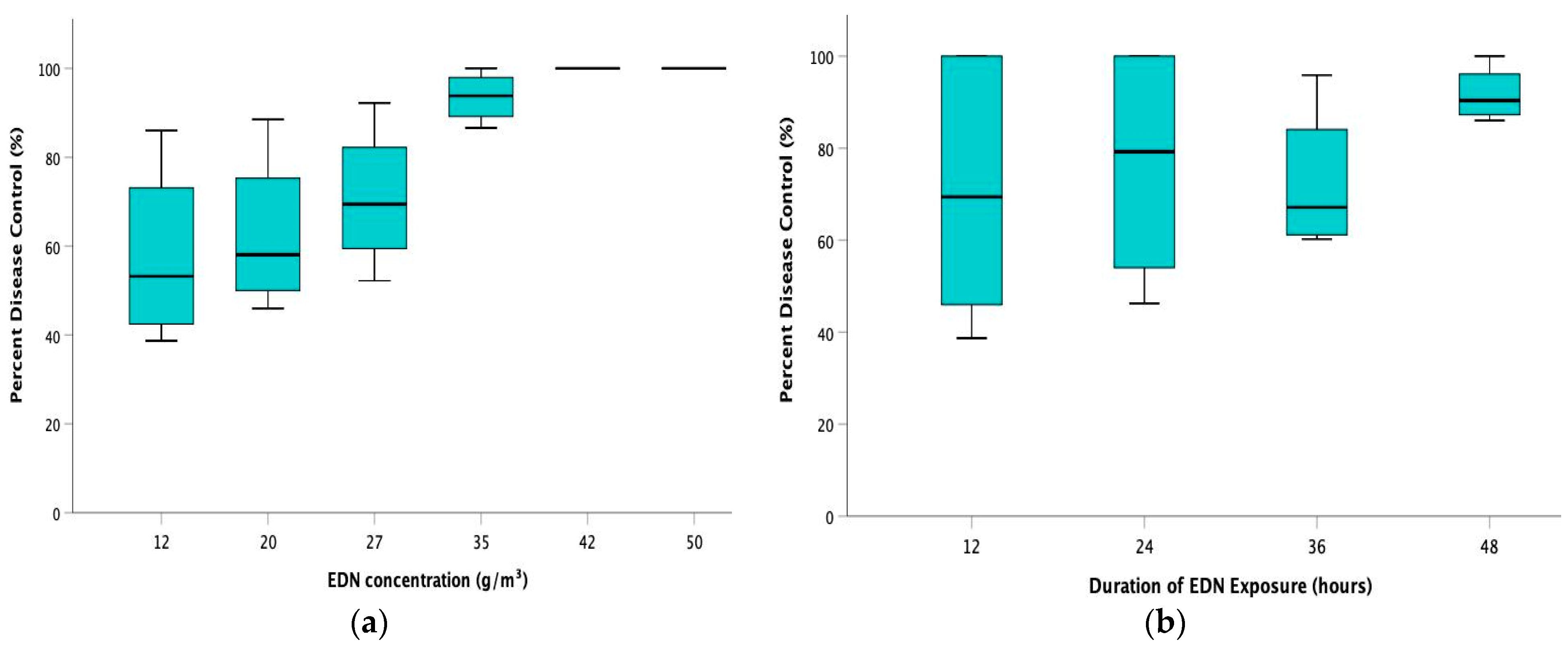
There was a significant reduction in disease severity in the treatment groups with all EDN concentration rates as compared to the non-fumigated control. The DSI in the treatment groups ranged from 4–57%, whereas the average DSI in the control group was 93.33% (Table 2). Among the treatment groups, the plants from the 12 g/m3-12 h variant reported the highest DSI of 57%. In contrast, plants from the treatment variant of 35 g/m3-36 h had the lowest DSI of 4%. It must be noted that there were no disease incidences (DSI = 0%) in the treatment groups of 42 g/m3 and 50 g/m3, as well as in the treatment variant of 35 g/m3-48 h. In all the treatment groups, the disease severity indices decreased gradually with an increased duration of exposure (Table 2). In the 12 g/m3 and 20 g/m3 treatment groups, DSI at 48 h of exposure were 13% and 10%, respectively, and significantly lower than that of the rest of the exposure durations (Table 2). In addition, in the treatment groups of 27 g/m3 and 35 g/m3, the average DSI ranged from 4–43% for different periods of exposure. However, there was no significant difference between treatments with varying periods of exposure despite the numerical decrease in the disease severity indices. The average DSI in the lower-to-intermediary EDN concentration groups of 12 g/m3 (39.25%), 20 g/m3 (32.5%), and 27 g/m3 (26.25%) were significantly higher as compared to the average DSI of the higher EDN concentrations groups of 35 g/m3 (6.25%), 42 g/m3 (0%), and 50 g/m3 (0%), respectively, at p < 0.001. However, there was no statistically significant difference between the concentration groups of 12 g/m3 and 20 g/m3, respectively. In the case of the duration of EDN exposure, the average DSI of the treatment group with 48 h of EDN exposure (7.5%) was statistically significantly lower in comparison to the other treatment groups, with 12 h (26.67%), 24 h (21.33%), and 36 h (24.75%), respectively, at p < 0.001. However, there was no statistically significant difference among the treatment groups with 12, 24 and 36 h duration of EDN exposure.
Analysis of PDC revealed that EDN fumigation resulted in an 81.39% average reduction in disease severity compared to the control group. Among the different EDN concentration groups, the 42 g/m3 and 50 g/m3 rates showed 100% control in the disease, followed by 35 g/m3 (93.56%) (Figure 1a). On the contrary, the concentration rate of 12 g/m3 exhibited the least control over the disease among the treatments, with a PDC of 57.8%. Concerning the duration of EDN exposure, a comparison of PDC showed that the highest control of the disease (91.69%) was achieved over 48 h, while the least control over the disease (70.58%) was obtained within 12-h of EDN exposure (Figure 1b). Spearman’s rank correlation analysis reported a statistically significant and strong positive correlation between EDN concentration rates and PDC (rs = 0.831) at p < 0.001. This indicates that EDN concentration rates have a considerable direct effect on the rate of control of the disease. On the other hand, there was a small positive correlation between the durations of EDN exposure and PDC (rs = 0.215), but it was statistically non-significant.
3.2. Effect on Seedling Emergence
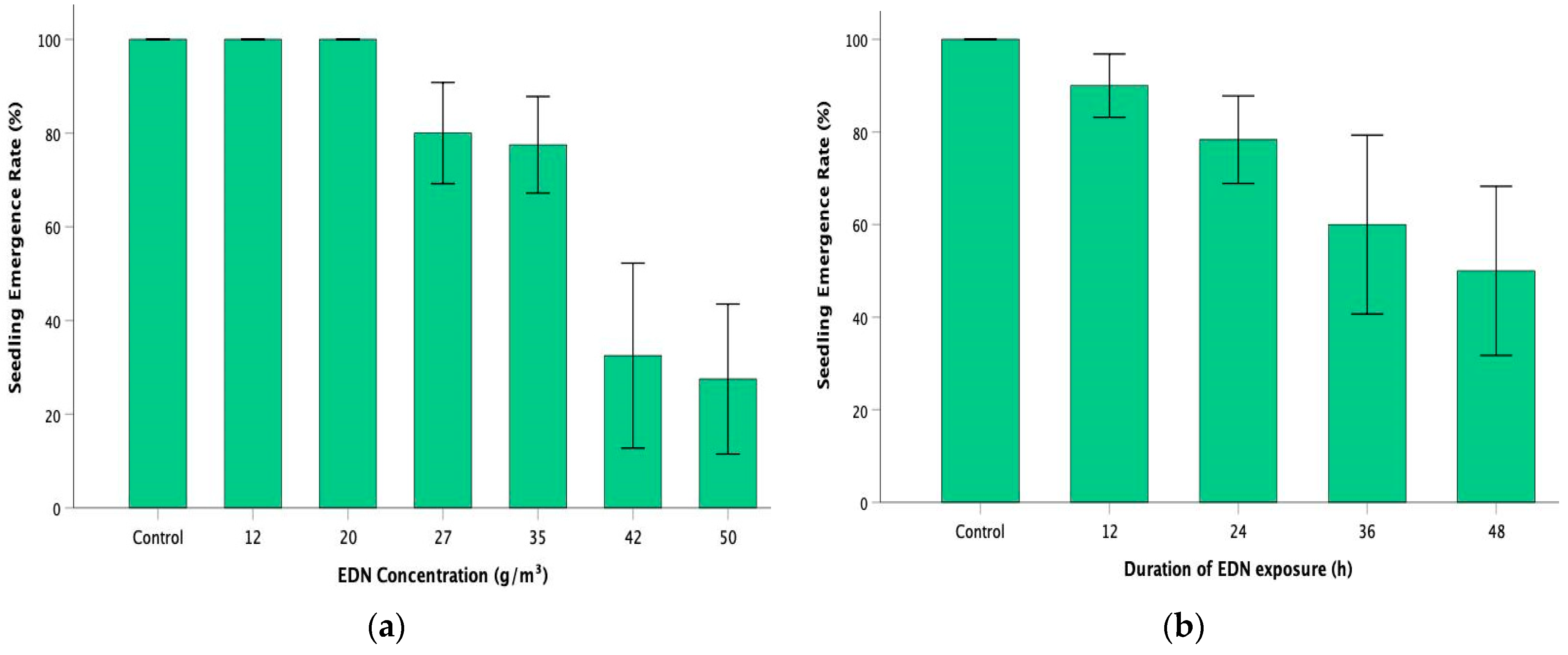
Seedling emergence was not affected at the lower concentration rates of EDN (12 g/m3 and 20 g/m3), with an emergence rate of 100% compared to the control (100%) (Figure 2a). It declined slightly to 80% and 77.5% in the intermediary EDN concentration groups of 27 g/m3 and 35 g/m3. However, there was a drastic decline in the seedling emergence at higher concentration rates of EDN (42 g/m3 and 50 g/m3), with 32.5% and 27.5% rates of seedling emergence, respectively. In the context of the duration of exposure, there were 90%, 78.3%, and 60% rates of seedling emergence in 12 h, 24 h, and 36 h of exposure, respectively (Figure 2b). The seedling emergence rate was the most reduced—50% in 48 h, compared to the other periods of exposure. It must be noted that there was no seedling emergence in the higher treatment variant groups of 42 g/m3-36 h, 42 g/m3-48 h, 50 g/m3-36 h, and 50 g/m3-48 h. Spearman’s rank correlation analysis reported a statistically significant and strong but negative correlation between EDN concentration rates and the seedling emergence rate (rs = −0.849) at p < 0.001. Similarly, there was a moderately strong but negative correlation between the durations of EDN exposure and the seedling emergence rate (rs = −0.539) at p = 0.002. This indicates that higher rates of EDN concentration and more extended periods of exposure to the fumigant have a negative impact on the seedling emergence rate of oilseed rape plants.
3.3. Effect on Plant Root Weight
Plants grown in the treated soil showed a considerable decline in root weight compared to the control group. The average root weight was the highest in the control plants (4.54 g), while the average root weights in treated plants ranged from 0.16 g to 2.89 g, with the lowest being in the 35 g/m3-48 h treatment variant (0.16 g) (Table 3). The cumulative average plant root weight in the treatment group was 1.21 g, compared with the control plants, with an average root weight of 4.54 g. In the treatment groups with lower-to-intermediary EDN concentration rates of 12–27 g/m3, there was a decrease in root weight, but it was not statistically significant (Figure 3a). On the other hand, the effect of EDN fumigation on the root weight of plants was more evident with concentration rates of 35 g/m3 to 50 g/m3. The average root weight of plants in these higher concentration groups ranged from 0.48 g to 0.56 g, which was 50–64% less than the plants in the treatment group of 12 g/m3. The root weight of treated plants decreased with the gradual increase in the periods of exposure across all treatments with different EDN concentrations. However, within individual concentration groups, the differences in root weight with varying exposure durations were not statistically significant (Table 3). It must be noted that the only exception was seen in the treatment group of 50 g/m3, where the root weight of plants with 12 h exposure (0.73 g) was significantly higher compared to that of 24 h duration of exposure (0.23 g) at p < 0.001. Regarding the period of EDN exposure, there was a significant reduction of 70.9% in average root weight in the treated plants (1.32 g) compared to the plants in the control group (4.54 g) at p < 0.001 (Figure 3b). However, the differences in the root weight among the treatment groups were not significant in the context of periods of EDN exposure.
3.4. Effect on Plant Shoot Weight
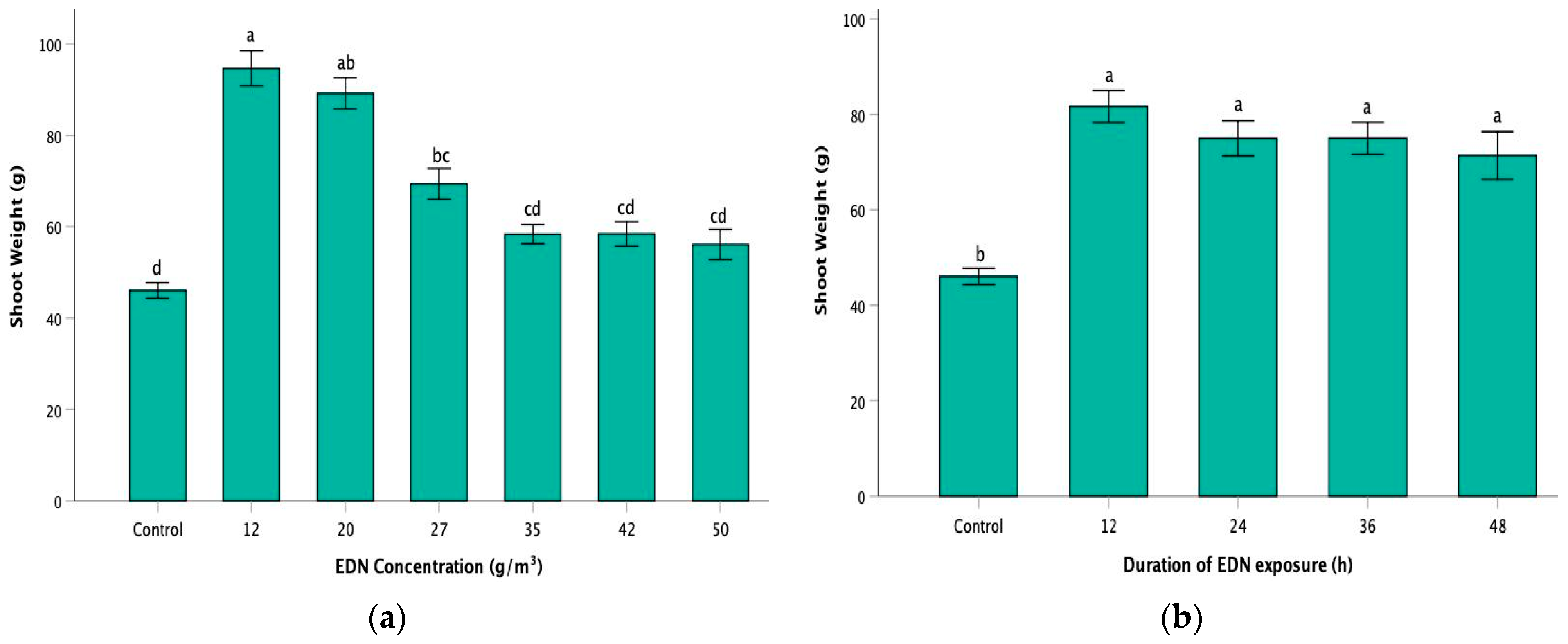
EDN fumigation treatment showed a considerable effect on the shoot weight of the plants. An increase of 58.24% was reported in the cumulative average plant shoot weight of the treatment group (72.87 g) when compared to that of the control group (46.05 g) (Table 4). The average shoot weight in treated plants ranged from 49.58 g to 106.76 g, with the highest being in the 12 g/m3-12 h treatment variant (106.76 g) (Table 4). The plants showed a gradual decrease in shoot weight with increasing concentration rates (Figure 4a). Comparison among the treatment groups in the context of the concentration rates indicated a significant difference in the average plant shoot weight in the lower concentration group (12–20 g/m3) compared to that in the higher concentration group (35–50 g/m3) at p < 0.05 (Figure 4a). However, there was no substantial difference in the average plant shoot weight between the lower rates of 12 g/m3 (94.65 g) and 20 g/m3 (89.18 g). Similarly, the differences in average plant shoot weight among the treatment groups with the higher rates of EDN fumigation (35–50 g/m3) were insignificant. In addition, the average plant shoot weights in the higher concentration rates of 42 g/m3 (57.09 g) and 50 g/m3 (55 g) contributed to a 24% and 19.4% increase, respectively, compared to the control group, but this increment was not statistically significant (Figure 4a). Regarding the effect of the period of EDN exposure on plant shoot weight, there was a remarkable increase of 57.22% in cumulative average shoot weight in the treated plants (72.4 g) compared to the plants in the control group (46.05 g) at p < 0.001 (Figure 4b). However, the differences in the average shoot weight among the treatment groups were insignificant in the context of periods of EDN exposure. It was also observed that the plant shoot weight declined with an increase in the duration of EDN exposure. While comparing the effect of exposure duration in the individual rate of concentration groups, the results indicated no significant differences in shoot weight across all treatment durations in the groups with 12 g/m3 and 20 g/m3 rates (Table 4). Regarding the treatment groups from 27 g/m3 to 50 g/m3, a similar pattern was observed, where the plant shoot weight increased substantially in 12 h of fumigation compared to the control. However, despite the numerical increments, the average shoot weights in the other exposure periods (24–48 h) were not significantly different from those of the control.
4. Discussion
EDN has been found effective in controlling various soil-borne pathogens such as Pythium sulcatum, Phytophthora cactorum, Sclerotium rolfsii, Rhizoctonia fragariae, and R. solani in previous studies [17,21,32]. However, the effectiveness of EDN against P. brassicae has not been studied before. Considering the extreme persistence of P. brassicae resting spores in the soil for as long as 20 years, it is necessary to find an efficient management tool against the pathogen. The current study suggests that EDN has a promising potential for controlling clubroot disease, as the fumigation treatment reduced DSI by 81.39% compared to the control. The disease was completely controlled at higher concentration rates of 42 g/m3 and 50 g/m3, regardless of fumigation duration, as well as 35 g/m3 for 48 h of fumigation. Furthermore, even at the lowest concentration of EDN (12 g/m3), the disease severity was reduced by 57.8%. Similar results were found in a prior study by Ren et al. [21], where Pythium sulcatum infection in carrot seedlings was reduced by 79% at EDN concentration rates of 4–20 g/m2. In the same investigation, full control was obtained in Phytophthora cactorum, Sclerotium rolfsii, and Rhizoctonia fragariae infections at higher EDN concentration rates of 25–50 g/m2. Similarly, Mattner et al. attained 100% control in Phytophthora cactorum, Rhizoctonia fragariae, and Sclerotium rolfsii inoculum up to a soil depth of 30 cm using an effective rate of 42.5 g/m2 EDN [32]. In another study by Yu et al. [25], EDN was found to be more effective than a fungicide concoction of 1,3-dichloropropene and 1 chloropicrin when used against Macrophomina phaseolina. Our findings are consistent with previous research and suggest that EDN is effective in controlling clubroot disease.
The seedling emergence rate was affected by the fumigation treatment mostly at higher EDN concentrations of 42 g/m3 and 50 g/m3, with no seedling emergence at combination treatment with higher concentrations and longer exposure durations. Several investigations in the past have reported that EDN can inhibit the germination of weed seeds such as Amaranthus hybridus, Portulaca oleracea, Senna obtusifolia, Cyperus esculentus, and C. rotundus [22,25,33]. HCN, released upon EDN disintegration, is a water-soluble compound, and its absorption in the weed seeds leads to the inhibition of germinating seeds [34]. The same mechanism may be considered the reason behind the inhibition of seedling emergence in oilseed rape plants in the current study. Higher doses of EDN are more likely to show this inhibiting effect as there may be higher retention of residue in the soil. Our study agrees with the findings of Mattner et al. [32], who reported suppression of seed germination in oilseed rape plants at an EDN concentration rate of 42.5 g/m3. Moreover, Stevens et al. found that tomato plant vigor declined at EDN rates of 56–67.2 g/m2 compared to lower doses of EDN and the non-fumigated control [22]. Similarly, Yu et al. reported the highest mortality rate of 29% in strawberry plants at an EDN concentration of 44.8 g/m2 [25]. However, both studies disproved any phytotoxic effect of EDN on the treated plants due to its highly volatile nature and shorter plant back interval. It should be noted that seedling transplants were employed in both studies and a post-fumigation interval period of one to two months was maintained. This longer interval period decreased the residue retention of EDN in the soil to a minimum, while giving the seedlings adequate time to establish their health. Furthermore, the suppression of seedling emergence is not limited to EDN. Dazomet, another fumigant used to control clubroot that releases methyl isothiocyanate (MITC) on degradation, has been shown to have similar effects on seedling emergence as EDN at higher doses [12]. The investigation has, however, indicated that the insufficient time gap between sowing and fumigation might have been the cause of this impact. Therefore, the post-fumigation interval period, as well as the duration of EDN exposure, may be considered important factors in the seedling emergence of plants.
The effect of EDN fumigation was evident in the root weight of the plants. The decrease in gall formation in the roots was responsible for the significant reduction in root weight in treated plants compared to control plants. Increased EDN concentration rates and longer durations of EDN exposure contributed further to lesser gall formation in roots, which subsequently resulted in lesser root weight in comparison to control plants. This suggests that the higher concentration rates of EDN (35–50 g/m3) were more efficient than the lower rates at preventing gall development. To the best of our knowledge, no previous research has investigated the impact of EDN fumigation on plant root weight. However, similar outcomes have been reported when metam sodium was used to treat clubroot disease; Hwang et al. found that the application of metam sodium at the rate of 800–1600 g/m3 caused a reduction in root weight of 81.7–83.7% [11].
EDN fumigation had a favorable effect on plant shoot weight at all concentration rates compared to the control, with the biggest increases occurring at the lower to intermediate concentration rates, 12 g/m3 to 27 g/m3. The reason for this could be ammonium, one of the breakdown products of EDN and a common source of nitrogen [35], and its deposition in the soil can lead to improved plant health [28]. In a previous study, Yu et al. discovered higher aboveground biomass in EDN-fumigated strawberry plants compared to controls in one of their trials, though the effect was statistically insignificant [25]. Additionally, Douda et al. also reported a significant increase in plant leaf weight when EDN was applied to control root-knot nematodes in carrots. These results are in line with the findings of our investigation [18]. On the other hand, the current findings also indicated a significant decline in the average plant shoot weight at higher concentration rates (35–50 g/m3), though it remained higher than in control plants. One possible explanation is that while the presence of ammonium in the soil is beneficial to plant health, an excess of it can cause plant toxicity in some cases [36]. An essential part of this process is soil nitrification, which involves the biological conversion of ammonium to nitrate in order to remove excess nitrogen from the soil. Since fumigants are generally non-selective to their target pathogens and pests, soil fumigation may lead to the killing of beneficial soil microbes, which, in turn, leads to the inhibition of the soil nitrification process [28,37,38,39]. This inhibition can remain for longer periods when soil fumigation is performed with higher EDN concentration rates or longer periods of fumigation. Waterford et al. found that EDN had a negative effect on strawberry runners when applied at a high rate of 100 g/m3 without a ventilation period [17]. Similarly, Stevens et al. reported that EDN at a higher concentration rate of 67.2 g/m2 reduced plant height and vigor in tomato plants when compared to a lower rate of 33.6 g/m2 as well as the non-fumigated control [22]. However, both studies rejected the possibility of phytotoxicity by EDN, as the effects were reversible after some time. Therefore, an interval period after fumigation seems crucial in this case.
Our findings indicate that EDN is effective against P. brassicae and thus may be considered as a potential management tool for clubroot disease. Indeed, EDN may be a better alternative to MITC-generating soil fumigants used against the pathogen, such as metam sodium and dazomet. According to Hwang et al. [11,12], dazomet and metam sodium could not eradicate the pathogen completely even at the highest rates of 24 g/m3 and 1.6 kg/m3. It was hypothesized that this was because MITC generators require warm temperatures for maximum effectiveness [11]. In contrast, EDN is effective in lower temperatures due to its lower boiling point of −21 °C. Moreover, since EDN is highly effective even at lower concentration rates, the lower doses can be deployed in integration with other clubroot management strategies. This will be advantageous because it will reduce the cost of the fumigation process as well as the exposure risk of toxic fumigants to the environment. Higher rates of EDN, on the other hand, may be considered in cases of severe infestation. However, a longer interval period after fumigation will be ideal for overall plant health in such cases, considering the reversible effect of higher EDN rates on plant shoot weight and seedling emergence. It should also be noted that since EDN is a highly toxic chemical, the treatments need to be administered by trained professionals with appropriate safety gear [23]. Additionally, soil fumigation in fields may involve labor-intensive operations which may eventually turn out to be expensive. In a previous study, Douda et al. reported a significant positive effect on carrot yield after soil fumigation with EDN in the previous season, leading to a conclusion that EDN treatment might be economically feasible under Czech Republic conditions [18]. However, considering the returns in yield, the cost efficiency of EDN treatment against clubroot disease in oilseed rape, as well as other cruciferous vegetables, needs to be evaluated in field conditions.
In summary, this is the first report to demonstrate the efficiency of EDN fumigation against clubroot disease in oilseed rape under controlled conditions. However, its effectiveness, like that of any other soil fumigant, is dependent on many environmental factors, including the soil texture, pH, and moisture level [34]. Therefore, further investigation of EDN effectiveness against P. brassicae under field conditions is required. Additional research may be useful to investigate the effect of EDN fumigation against P. brassicae in different soil textures, as well as its residue retention potential in the soil, leading to any possible phytotoxic effect in different cruciferous crops.
Author Contributions
Statistical analysis, writing, U.R.P.; fumigation of samples J.H.; samples evaluation, U.R.P., J.W. and M.M.; conceptualization, M.Z. and M.M.; supervision, manuscript review, and editing, M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Technology Agency of the Czech Republic, Grant No. TH02030328.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Dixon, G.R. The biology of Plasmodiophora brassicae Wor.—A review of recent advances. Acta Hortic. 2006, 706, 271–282. [Google Scholar] [CrossRef]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Zamani-Noor, N.; Wallenhammar, A.-C.; Kaczmarek, J.; Patar, U.R.; Zouhar, M.; Manasova, M.; Jędryczka, M. Pathotype characterization of Plasmodiophora brassicae, the cause of clubroot in central Europe and Sweden (2016–2020). Pathogens 2022, 11, 1440. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Lazarovits, G. Effect of soil application of AG3 phosphonate on the severity of clubroot of bok choy and cabbage caused by Plasmodiophora brassicae. Plant Dis. 2006, 90, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Gossen, B.D.; Adhikari, K.K.C.; Mcdonald, M.R. Effect of seeding date on development of clubroot in short-season brassica crops. Can. J. Plant Pathol. 2012, 34, 516–523. [Google Scholar] [CrossRef]
- Mitani, S.; Sugimoto, K.; Hayashi, H.; Takii, Y.; Ohshima, T.; Matsuo, N. Effects of cyazofamid against Plasmodiophora brassicae Woronin on chinese cabbage. Pest Manag. Sci. 2003, 59, 287–293. [Google Scholar] [CrossRef]
- Peng, G.; McGregor, L.; Lahlali, R.; Gossen, B.D.; Hwang, S.F.; Adhikari, K.K.; Strelkov, S.E.; McDonald, M.R. Potential biological control of clubroot on canola and crucifer vegetable crops. Plant Pathol. 2011, 60, 566–574. [Google Scholar] [CrossRef]
- Peng, G.; Lahlali, R.; Hwang, S.F.; Pageau, D.; Hynes, R.K.; McDonald, M.R.; Gossen, B.D.; Strelkov, S.E. Special Issue: Crop rotation, cultivar resistance, and fungicides/biofungicides for managing clubroot (Plasmodiophora brassicae) on canola. Can. J. Plant Pathol. 2014, 36, 99–112. [Google Scholar] [CrossRef]
- Struck, C.; Rüsch, S.; Strehlow, B. Control strategies of clubroot disease caused by Plasmodiophora brassicae. Microorganisms 2022, 10, 620. [Google Scholar] [CrossRef]
- Hwang, S.F.; Ahmed, H.U.; Strelkov, S.E.; Zhou, Q.; Gossen, B.D.; Peng, G.; Turnbull, G.D.; Fu, H. Effects of rate and application method on the efficacy of metam sodium to reduce clubroot (Plasmodiophora brassicae) of canola. Eur. J. Plant Pathol. 2018, 150, 341–349. [Google Scholar] [CrossRef]
- Hwang, S.-F.; Ahmed, H.U.; Strelkov, S.; Zhou, Q.; Gossen, B.; McDonald, M.-R.; Peng, G.; Turnbull, G.D. Suppression of clubroot by Dazomet fumigant. Can. J. Plant Sci. 2017, 98, 172–182. [Google Scholar] [CrossRef]
- Donald, C.; Porter, I. Integrated control of clubroot. J. Plant Growth Regul. 2009, 28, 289–303. [Google Scholar] [CrossRef]
- Hwang, S.F.; Ahmed, H.U.; Zhou, Q.; Strelkov, S.E.; Gossen, B.D.; Peng, G.; Turnbull, G.D. Efficacy of vapam fumigant against clubroot (Plasmodiophora brassicae) of canola. Plant Pathol. 2014, 63, 1374–1383. [Google Scholar] [CrossRef]
- Hooper, J.L.; Desmarchelier, J.M.; Ren, Y.; Allen, S.E. Toxicity of cyanogen to insects of stored grain. Pest Manag. Sci. 2003, 59, 353–357. [Google Scholar] [CrossRef]
- Wright, E.J.; Ren, Y.L.; Dowsett, H. Cyanogen: A new fumigant with potential for timber. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, Orlando, FL, USA, 6–8 November 2002. [Google Scholar]
- Waterford, C.J.; Ren, Y.L.; Mattner, S.; Sarwar, M. Ethanedinitrile (C2N2)—A novel soil fumigant for insect, nematode, pathogen and weed control. In Proceedings of the 2002 Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, Orlando, FL, USA, 31 October–3 November 2004. [Google Scholar]
- Douda, O.; Manasova, M.; Zouhar, M.; Hnatek, J.; Stejskal, V. Field validation of the effect of soil fumigation of ethanedinitrile (EDN) on the mortality of Meloidogyne hapla and carrot yield parameters. Agronomy 2021, 11, 208. [Google Scholar] [CrossRef]
- Pranamornkith, T.; Hall, M.K.D.; Najar-Rodriguez, A.; Adlam, A.R.; Somerfield, K.G.; Page, B.B.C.; Hedderley, D.I.; Brash, D.W. Ethanedinitrile: Potential methyl bromide alternative to control Arhopalus ferus (Mulsant) in New Zealand sawn timber exports. N. Z. Plant Prot. 2014, 67, 75–79. [Google Scholar] [CrossRef]
- Ramadan, G.R.M.; Zhu, K.Y.; Abdelgaleil, S.A.M.; Shawir, M.S.; El-Bakary, A.S.; Edde, P.A.; Phillips, T.W. Ethanedinitrile as a fumigant for Lasioderma serricorne (Coleoptera: Anobiidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae): Toxicity and mode of action. J. Econ. Entomol. 2020, 113, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.L.; Waterford, C.J.; Matthiessen, J.N.; Mattner, S.; Gregorio, R.; Sarwar, M. First results from ethanedinitrile (C2N2) field trials in Australia. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, San Diego, CA, USA, 3–6 November 2003. [Google Scholar]
- Stevens, M.C.; Freeman, J.H.; Boyd, N.S. Impact of ethanedinitrile rates and application method on nutsedge species and tomato root galling. Crop. Prot. 2019, 116, 1–6. [Google Scholar] [CrossRef]
- O’Brien, I.G.; Desmarchelier, F.J.M.; Ren, Y. Cyanogen Fumigants and Methods of Fumigation Using. Cyanogen. Patent No. 6001383, 14 December 1999. [Google Scholar]
- Matheson Tri-Gas Cyanogen Safety Data Sheet; Matheson Tri-Gas: Basking Ridge NJ, USA, 2008.
- Yu, J.; Baggio, J.S.; Boyd, N.S.; Freeman, J.H.; Peres, N.A. Evaluation of ethanedinitrile (EDN) as a preplant soil fumigant in Florida strawberry production. Pest Manag. Sci. 2020, 76, 1134–1141. [Google Scholar] [CrossRef]
- Najar-Rodriguez, A.J.; Afsar, S.; Esfandi, K.; Hall, M.K.D.; Adlam, A.R.; Wilks, C.; Noakes, E.; Richards, K. Laboratory toxicity and large-scale commercial validation of the efficacy of ethanedinitrile, a potential alternative fumigant to methyl bromide, to disinfest New Zealand Pinus radiata export logs. J. Stored Prod. Res. 2020, 88, 101671. [Google Scholar] [CrossRef]
- Ren, Y.L.; Sarwar, M.; Wright, E.J. Development of cyanogen for soil fumigation. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, Orlando, FL, USA, 6–8 November 2002. [Google Scholar]
- Stevens, M.C.; Yang, R.; Freeman, J.H. Deposition and transformation of nitrogen after soil fumigation with ethanedinitrile. HortScience 2020, 55, 2023–2027. [Google Scholar] [CrossRef]
- Tewari, J.P.; Strelkov, S.E.; Orchard, D.; Hartman, M.; Lange, R.M.; Turkington, T.K. Identification of clubroot of crucifers on canola (Brassica napus) in Alberta. Can. J. Plant Pathol. 2005, 27, 143–144. [Google Scholar] [CrossRef]
- Kuginuki, Y.; Yoshikawa, H.; Hirai, M. Variation in virulence of Plasmodiophora brassicae in Japan tested with clubroot-resistant cultivars of chinese cabbage (Brassica rapa L. ssp. pekinensis). Eur. J. Plant Pathol. 1999, 105, 327–332. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Tewari, J.P.; Smith-Degenhardt, E. Characterization of Plasmodiophora brassicae populations from Alberta, Canada. Can. J. Plant Pathol. 2006, 28, 467–474. [Google Scholar] [CrossRef]
- Mattner, S.W.; Gounder, R.K.; Mann, R.C.; Porter, I.J.; Matthiessen, J.N.; Ren, Y.L.; Sarwar, M. Ethanedinitrile (C2N2)—A novel soil fumigant for strawberry production. Acta Hortic. 2006, 708, 197–203. [Google Scholar] [CrossRef]
- Rosskopf, E.N.; Kokalis-Burelle, N.; Peterson, G.; Waterford, C. Preliminary Investigation of ethanedinitrile for control of weeds and nematodes important to Florida production systems. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, San Diego, CA, USA, 3–6 November 2003. [Google Scholar]
- Thalavaiasundaram, S.; Ajwa, H.; Stevens, M.C. Dose-response of pests to ethanedinitrile dose-response of weed seeds, soil borne pathogens, and plant-parasitic nematodes to ethanedinitrile. Australas. Plant Pathol. 2023, 52, 133–143. [Google Scholar] [CrossRef]
- Ajwa, H.; Xuan, R.; Guerrero, J. Fate of EDN fumigant in soil. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, Orlando, FL, USA, 8–10 November 2016. [Google Scholar]
- Simonne, E.H.; Mills, H.A.; Smittle, D.A. Ammonium reduces growth, fruit yield and fruit quality of watermelon. J. Plant Nutr. 1992, 15, 2727–2741. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Powlson, D.S. Residual effects of soil fumigation on soil respiration and mineralization. Soil Biol. Biochem. 1970, 2, 99–108. [Google Scholar] [CrossRef]
- Rovira, A.D. Studies on Soil Fumigation—I: Effects on ammonium, nitrate and phosphate in soil and on the growth, nutrition and yield of wheat. Soil Biol. Biochem. 1976, 8, 241–247. [Google Scholar] [CrossRef]
- Shen, S.M.; Pruden, G.; Jenkinson, D.S. Mineralization and immobilization of nitrogen in fumigated soil and the measurement of microbial biomass nitrogen. Soil Biol. Biochem. 1984, 16, 437–444. [Google Scholar] [CrossRef]
Figure 1.
Effect of (a) rate of ethanedinitrile (EDN) concentration and (b) duration of EDN exposure on the disease severity index of clubroot (%). The medians (middle line) and 25% and 75% quartiles, as well as the minimum and maximum values, are presented by box–whisker plots.
Figure 1.
Effect of (a) rate of ethanedinitrile (EDN) concentration and (b) duration of EDN exposure on the disease severity index of clubroot (%). The medians (middle line) and 25% and 75% quartiles, as well as the minimum and maximum values, are presented by box–whisker plots.

Figure 2.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the seedling emergence rate (%) in oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean.
Figure 2.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the seedling emergence rate (%) in oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean.

Figure 3.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the plant root weight of oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean. A mixed model ANOVA reported a significant effect of EDN concentration rates, durations of EDN exposure, and their interactions on clubroot disease severity at p < 0.001. Results of the o-way ANOVA–Welch followed by the Games–Howell post hoc test, performed to analyze the main factor effects on plant root weight, are shown. Bars with different superscript letters indicate significant differences (p < 0.05).
Figure 3.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the plant root weight of oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean. A mixed model ANOVA reported a significant effect of EDN concentration rates, durations of EDN exposure, and their interactions on clubroot disease severity at p < 0.001. Results of the o-way ANOVA–Welch followed by the Games–Howell post hoc test, performed to analyze the main factor effects on plant root weight, are shown. Bars with different superscript letters indicate significant differences (p < 0.05).

Figure 4.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the plant shoot weight of oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean. A mixed model ANOVA reported a significant effect of EDN concentration rates and durations of EDN exposure on plant shoot weight at p < 0.001. However, there was no significant interaction effect of EDN concentration rates and exposure durations on plant shoot weight (p = 0.32). Results of the one-way ANOVA–Welch test followed by the Games–Howell post hoc test, performed to analyze the main factor effects on plant shoot weight, are shown. Bars with different superscript letters indicate significant differences (p < 0.05).
Figure 4.
Effect of (a) rate of EDN concentration and (b) duration of EDN exposure on the plant shoot weight of oilseed rape. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. Error bars represent one standard error of the mean. A mixed model ANOVA reported a significant effect of EDN concentration rates and durations of EDN exposure on plant shoot weight at p < 0.001. However, there was no significant interaction effect of EDN concentration rates and exposure durations on plant shoot weight (p = 0.32). Results of the one-way ANOVA–Welch test followed by the Games–Howell post hoc test, performed to analyze the main factor effects on plant shoot weight, are shown. Bars with different superscript letters indicate significant differences (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Schematic table showing the distribution of different treatment variants and control.
| Control 1 (n = 10) | V1 = C1 × T1 (n = 10) | V2 = C1 × T2 (n = 10) | V3 = C1 × T3 (n = 10) | V4 = C1 × T4 (n = 10) |
| Control 2 (n = 10) | V5 = C2 × T1 (n = 10) | V6 = C2 × T2 (n = 10) | V7 = C2 × T3 (n = 10) | V8 = C2 × T4 (n = 10) |
| Control 3 (n = 10) | V9 = C2 × T1 (n = 10) | V10 = C2 × T2 (n = 10) | V11 = C2 × T3 (n = 10) | V12 = C2 × T4 (n = 10) |
| Control 4 (n = 10) | V13 = C3 × T1 (n = 10) | V14 = C3 × T2 (n = 10) | V15 = C3 × T3 (n = 10) | V16 = C3 × T4 (n = 10) |
| Control 5 (n = 10) | V17 = C4 × T1 (n = 10) | V18 = C4 × T2 (n = 10) | V19 = C4 × T3 (n = 10) | V20 = C4 × T4 (n = 10) |
| Control 6 (n = 10) | V21 = C5 × T1 (n = 10) | V22 = C5 × T2 (n = 10) | V23 = C5 × T3 (n = 10) | V24 = C5 × T4 (n = 10) |
Control 1–6 represents pathogen-inoculated, non-fumigated soil. V1–V24 represent different variants of treatment. C1, C2, C3, C4, and C5 indicate concentration rates of 12 g/m3, 20 g/m3, 27 g/m3, 35 g/m3, 42 g/m3, and 50 g/m3 respectively. T1, T2, T3, and T4 represent 12 h, 24 h, 36 h, and 48 h of duration of treatment. The number of pots for each treatment variant and control is indicated by ‘n’.
Table 2.
Effect of the rate of ethanedinitrile (EDN) concentration and duration of EDN exposure on clubroot severity (DSI%) of oilseed rape.
Table 2.
Effect of the rate of ethanedinitrile (EDN) concentration and duration of EDN exposure on clubroot severity (DSI%) of oilseed rape.
| Concentration/ Duration | Control | 12 h | 24 h | 36 h | 48 h |
|---|---|---|---|---|---|
| 12 g/m3 | 93 Aa | 57 Ab | 50 Ab | 37 Abc | 13 Ac |
| 20 g/m3 | 87 Aa | 47 Ab | 40 Ac | 33 Abc | 10 Ac |
| 27 g/m3 | 90 Ba | 43 Bb | 30 Bb | 25 Bb | 7 Bb |
| 35 g/m3 | 97 BCa | 13 BCb | 8 BCb | 4 BCb | 0 BCb |
| 42 g/m3 | 100 Ca | 0 Cb | 0 Cb | NA | NA |
| 50 g/m3 | 93 Ca | 0 Cb | 0 Cb | NA | NA |
GEE analysis reporting a significant effect of EDN concentration rates, durations of EDN exposure, and their interactions on clubroot disease severity at p < 0.001 are shown. Kruskal–Wallis test results in individual concentration groups reporting a significant effect of duration of EDN exposure on clubroot disease severity at p < 0.05 are shown. Different superscript capital letters indicate significant differences between rows (p < 0.05). Variants in a row followed by different superscript lower-case letters are significantly different based on the Dunn–Bonferroni test at p < 0.05. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. NA stands for no plants in the particular variant.
Table 3.
Effect of rate of EDN concentration and duration of EDN exposure on root weight of oilseed rape.
Table 3.
Effect of rate of EDN concentration and duration of EDN exposure on root weight of oilseed rape.
| Concentration/ Duration | Control | 12 h | 24 h | 36 h | 48 h |
|---|---|---|---|---|---|
| 12 g/m3 | 4.61 ± 1.48 a | 2.89 ± 0.91 b | 2.51 ± 0.68 b | 2.26 ± 1.27 b | 2.18 ± 0.74 b |
| 20 g/m3 | 4.18 ± 0.54 a | 2.68 ± 0.92 b | 2.29 ± 0.49 b | 1.82 ± 1.50 b | 1.55 ± 0.61 b |
| 27 g/m3 | 4.76 ± 1.80 a | 1.61 ± 1.07 b | 1.47 ± 1.07 b | 0.88 ± 0.70 b | 0.57 ± 0.09 b |
| 35 g/m3 | 4.96 ± 2.50 a | 1.05 ± 0.64 b | 0.69 ± 0.53 b | 0.29 ± 0.25 b | 0.16 ± 0.13 b |
| 42 g/m3 | 4.61 ± 1.93 a | 0.80 ± 0.43 b | 0.32 ± 0.20 b | NA | NA |
| 50 g/m3 | 4.13 ± 1.22 a | 0.73 ± 0.15 b | 0.23 ± 0.19 c | NA | NA |
One-way ANOVA test results in individual concentration groups reporting a significant effect of the duration of EDN exposure on plant root weight are shown. Values are presented as mean ± standard deviation (SD). Variants in a row followed by different superscript letters are significantly different based on Tukey’s HSD test at p ≤ 0.001. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. NA stands for no plants in the particular variant.
Table 4.
Effect of rate of EDN concentration and duration of EDN exposure on shoot weight of oilseed rape.
Table 4.
Effect of rate of EDN concentration and duration of EDN exposure on shoot weight of oilseed rape.
| Concentration/ Duration | Control | 12 h | 24 h | 36 h | 48 h |
|---|---|---|---|---|---|
| 12 g/m3 | 49.07 ± 8.76 b | 106.76 ± 20.21 a | 98.18 ± 24.58 a | 89.23 ± 15.60 a | 84.43 ± 31.37 a |
| 20 g/m3 | 41.11 ± 14.55 b | 101.89 ± 18.77 a | 93.32 ± 17.36 a | 83.76 ± 17.41 a | 77.74 ± 27.10 a |
| 27 g/m3 | 53.69 ± 19.58 b | 80.72 ± 21.20 a | 68.52 ± 18.32 ab | 65.51 ± 15.73 ab | 54.34 ± 7.64 ab |
| 35 g/m3 | 47.08 ± 14.16 b | 65.73 ± 11.40 a | 57.21 ± 7.23 ab | 55.76 ± 14.04 ab | 49.65 ± 6.82 ab |
| 42 g/m3 | 42.31 ± 10.69 b | 62.28 ± 15.19 a | 52.28 ± 8.37 ab | NA | NA |
| 50 g/m3 | 43.02 ± 6.4 b | 60.42 ± 11.74 a | 49.58 ± 3.02 ab | NA | NA |
One-way ANOVA test results in individual concentration groups reporting a significant effect of the duration of EDN exposure on plant root weight are shown. Values are presented as mean ± SD. Variants in a row followed by different superscript letters are significantly different based on Tukey’s HSD test at p ≤ 0.001. Control represents oilseed rape plants grown in pathogen-inoculated, non-fumigated soil. NA stands for no plants in the particular variant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Patar, U.R.; Manasova, M.; Hnatek, J.; Wenzlova, J.; Zouhar, M. Evaluation of Ethanedinitrile as a Potential Fumigant against Plasmodiophora brassicae, the Clubroot Pathogen. Agronomy 2023, 13, 1177. https://doi.org/10.3390/agronomy13041177
AMA Style
Patar UR, Manasova M, Hnatek J, Wenzlova J, Zouhar M. Evaluation of Ethanedinitrile as a Potential Fumigant against Plasmodiophora brassicae, the Clubroot Pathogen. Agronomy. 2023; 13(4):1177. https://doi.org/10.3390/agronomy13041177
Chicago/Turabian StylePatar, Usha Rani, Marie Manasova, Jonas Hnatek, Jana Wenzlova, and Miloslav Zouhar. 2023. "Evaluation of Ethanedinitrile as a Potential Fumigant against Plasmodiophora brassicae, the Clubroot Pathogen" Agronomy 13, no. 4: 1177. https://doi.org/10.3390/agronomy13041177
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.