
Productivity and Nutritional Quality of Nitrogen-Fixing and Non-Fixing Shrub Species for Ruminant Production
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Nursery Management and Establishment
2.3. Experimental Design
- 50:50 mixture of Leucaena and Guazuma;
- 50:50 mixture of Leucaena and Moringa;
- Leucaena monoculture;
- Guazuma monoculture;
- Moringa monoculture.
2.4. Chemical Analysis
2.5. Tree Measurements
2.6. Statistical Analysis
3. Results
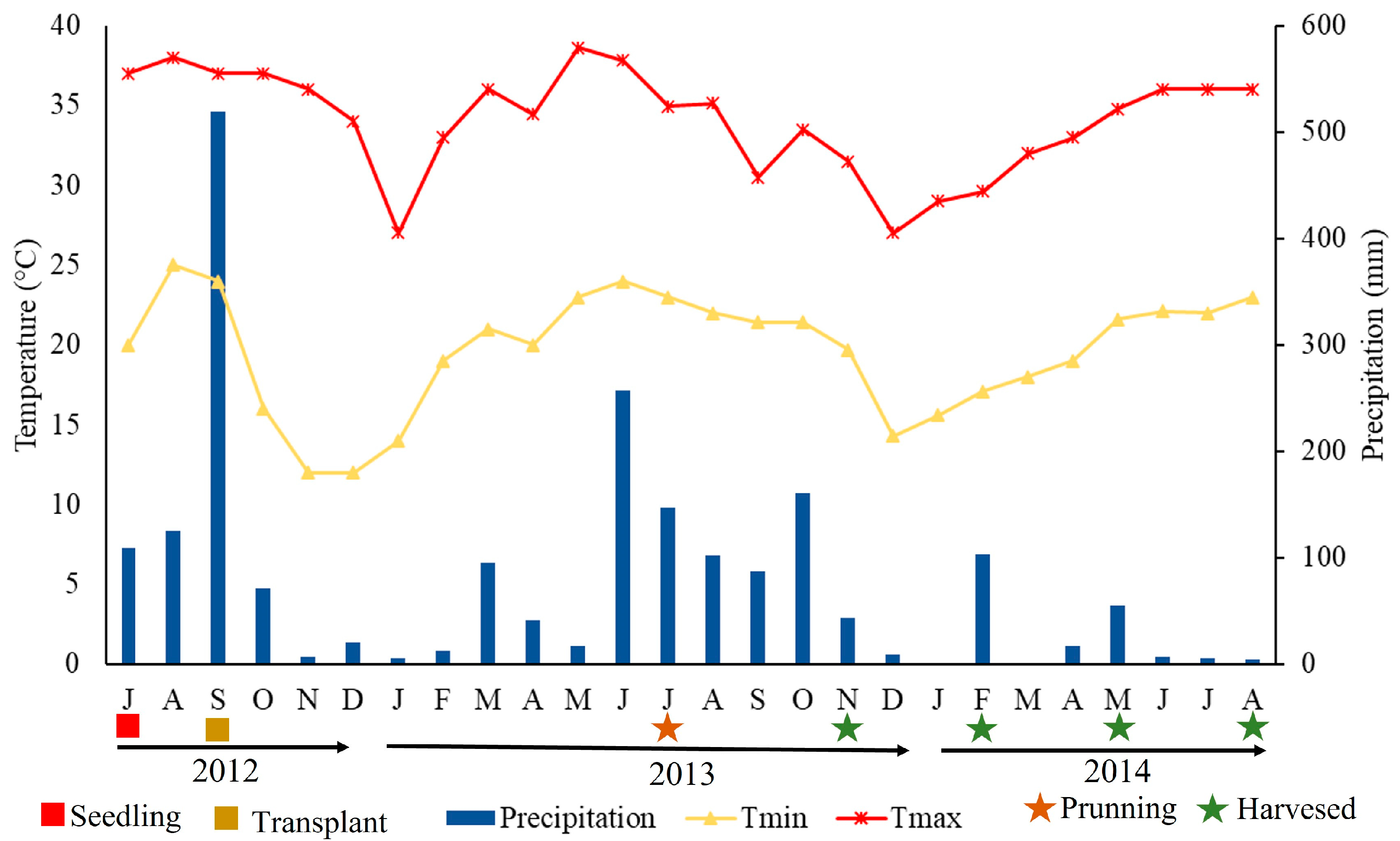
3.1. Environmental Variables
3.2. Soil Chemical and Physical Characteristics
3.3. Tree Survival in Mixtures and Pure Stands
3.4. Shrub Growth in Pure and Mixed Stands
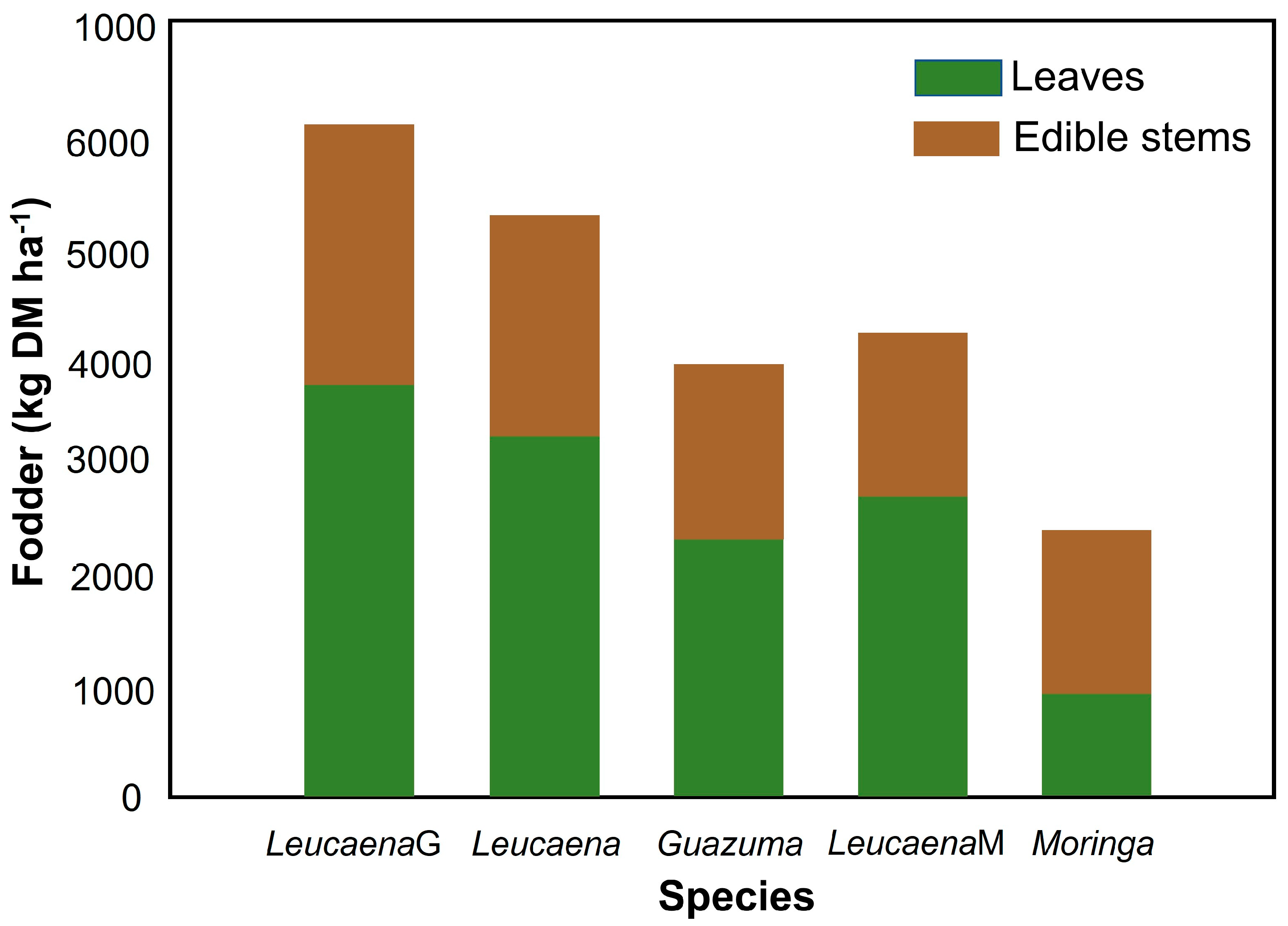
3.5. Biomass Yield
3.6. Chemical Composition of Pruned Foliage at Three-Month Regrowth Intervals
4. Discussion
4.1. Tree Survival and Growth
4.2. Biomass Yield
4.3. Nutritional Quality
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gómez-Pompa, A.; Bainbridge, D.A. Tropical forestry as if people mattered. In Tropical Forest: Management and Ecology; Lugo, A., Lowe, C., Eds.; Springer: New York, NY, USA, 1995; pp. 408–422. [Google Scholar]
- Rodríguez-Robayo, K.J.; Méndez-López, M.E.; Molina-Villegas, A.; Juárez, L. What do we talk about when we talk about milpa? A conceptual approach to the significance, topics of research and impact of the mayan milpa system. J. Rural. Stud. 2020, 77, 47–54. [Google Scholar] [CrossRef]
- Rico Gray, V.J.G.; Franco, A.P.G.; Sima, P. Composition and structure of a tropical dry forest in Yucatan, Mexico. Int. J. Ecol. Environ. Sci. 1998, 14, 21–29. [Google Scholar]
- Fernández, G.C.; Tapia, J.L.; de Stefano, R.D.; Ramírez, I.M.; Can, L.; Hernández, S.; Castillo, A. La Flora de la Península de Yucatán Mexicana: 250 años de conocimiento florístico. CONABIO. Biodiversitas 2012, 101, 6–10. [Google Scholar]
- Valencia-Salazar, S.S.V.; Vázquez, A.T.P.; Botero, I.C.M.; Balbuena, F.J.L.; Narváez, J.J.U.; Campos, M.R.S.; Ramírez y Avilés, L.; Solorio-Sánchez, F.J.; Ku-Vera, J.C. Potential of Samanea saman pod meal for enteric methane mitigation in crossbred heifers fed low-quality tropical grass. Agric. For. Meteorol. 2018, 258, 108–116. [Google Scholar] [CrossRef]
- Young, A. Agroforestry for Soil Management; CAB International: Wallingford, UK, 1997; p. 320. [Google Scholar]
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Oliveira, N. Nutrient contribution of litterfall in a short rotation plantation of pure or mixed plots of Populus alba L. and Robinia pseudoacacia L. Forests 2020, 11, 1133. [Google Scholar] [CrossRef]
- Hernández-Daumas, S. Carbon and Nitrogen Cycling in a Tree—Grass Intercropping System in the Humid Tropics of Mexico. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 2000. [Google Scholar]
- Dakora, F.D.; Keya, S.O. Contribution of legume nitrogen fixation to sustainable agriculture in sub-Saharan Africa. Soil Biol. Biochem. 1997, 26, 809–817. [Google Scholar] [CrossRef]
- Parrota, J. Productivity, nutrient cycling, and succession in single- and mixed-species plantations of Casuarina equisetifolia, Eucalyptus robusta, and Leucaena leucocephala in Puerto Rico. For. Ecol. Manag. 1999, 124, 45–77. [Google Scholar] [CrossRef]
- Derero, A.; Kitaw, G. Nutritive values of seven high priority indigenous fodder tree species in pastoral and agro-pastoral areas in Eastern Ethiopia. Agric. Food Secur. 2018, 7, 68. [Google Scholar] [CrossRef]
- Flores, J.S.; Bautista, F. Knowledge of the Yucatec Maya in seasonal tropical forest management: The forage plants. Rev. Mex. Biodivers. 2012, 83, 503–518. [Google Scholar]
- Mesa-Sierra, N.; de la Peña-Domene, M.; Campo, J.; Giardina, C.P. Restoring Mexican Tropical Dry Forests: A National Review. Sustainability 2022, 14, 3937. [Google Scholar] [CrossRef]
- Leng, R.A. Tree Foliage in Ruminant Nutrition; FAO Animal Production and Health Paper 139; FAO: Rome, Italy, 1997; 100p. [Google Scholar]
- Bryant, J.A. Nitrogen-fixing leguminous trees and shrubs: A basic resource of agroforestry. In The Silvicultural Basis for Agroforestry Systems; Ashton, M.S., Montagnini, F., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 41–60. [Google Scholar]
- Rathor, B.M. Intercropping leguminous crop plant with non-legumes. Life Sci. Inform. Publ. 2016, 2, 183–201. [Google Scholar]
- Schucknecht, A.; Krämer, A.; Asam, S.; Mejia-Aguilar, A.; Garcia-Franco, N.; Schuchardt, M.A.; Jentsch, A.; Kiese, R. Vegetation traits of pre-Alpine grasslands in southern Germany. Sci. Data 2020, 7, 316. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Medina, H.; Cobos-Gasca, V.; Acosta-Rodríguez, J.L.; Peña Fierro, S.; Castilla-Martínez, M.; Castillo-Carrillo, C.; Franco-Brito, S.; López-Castillo, D.; López-Díaz, M.; Luna-Flores, W.; et al. La sequía de la península de Yucatán. Tecnol. Cienc. Agua 2016, 7, 151–165. [Google Scholar]
- Bautista, F.; Díaz-Garrido, S.; Castillo-González, M.; Zinck, J.A. Spatial heterogeneity of the soil cover in the Yucatan Karst: Comparison of Mayan, WRB, and numerical classifications. Eurasian Soil Sci. 2005, 38, 80–87. [Google Scholar]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis, Part 3: Chemical Analysis; Sparks, D.L., Ed.; Soil Science Society of America, Inc.; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis, Part 3: Chemical Analysis; Sparks, D.L., Ed.; Soil Science Society of America, Inc.; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon and Organic Matter. In Methods of Soil Analysis, Part 3: Chemical Analysis; Sparks, D.L., Ed.; Soil Science Society of America, Inc.; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Kuo, S.P. Phosphorus. In Methods of Soil Analysis, Part 3: Chemical Analysis; Sparks, D.L., Ed.; Soil Science Society of America, Inc.; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 869–920. [Google Scholar]
- Allen, E.S. Chemical Analysis of Ecological Material; Blackwell: Oxford, UK, 1989; 565p. [Google Scholar]
- Rowell, L.D. Soil Science: Methods and Applications; Department of Soil Science, University of Reading co-published in the US with John Willey and Sons Inc.: New York, NY, USA, 1994; 350p. [Google Scholar]
- Van Soest, J.P.; Robertson, J.B.; Lewis, B.A. Methods for dietary neutral detergent fiber and nonstarch polysacharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Quantification of Tannins in Tree Foliage: A Laboratory Manual for FAO/IAEA Co-Ordinated Research Project on “Use of Nuclear and Related Techniques to Develop Simple Tannin Assays for Predicting and Improving the Safety and Efficiency of Feeding Ruminants on Tanniniferous Tree Foliage”; FAO/IAEA Working Document; IAEA: Vienna, Italy, 2000. [Google Scholar]
- Harinder, P.S.M. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Food Agriculture Organization of the United Nations/International Atomic Energy (FAO/IAEA): Rome, Italy, 2003; pp. 49–53. [Google Scholar]
- Khanna, P.K. Comparison of growth and nutrition of young monocultures and mixed stands of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 1997, 94, 105–113. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Khanna, P.K. Growth dynamics in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2004, 193, 81–95. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G. Thinking about efficiency of resource use in forests. For. Ecol. Manag. 2004, 193, 5–16. [Google Scholar] [CrossRef]
- Chirwa, T.S.; Mafongoya, P.L.; Chintu, R. Mixed planted-fallow using coppicing and non-coppicing tree species for degraded Acrisols in eastern Zambia. Agrofor. Syst. 2003, 59, 243–251. [Google Scholar] [CrossRef]
- Salako, F.K.; Tiang, G. Litter and biomass production from planted and natural fallows on a degraded soil in southwestern Nigeria. Agrofor. Syst. 2001, 51, 239–251. [Google Scholar] [CrossRef]
- Manh, L.H.; Dung, N.N.X.; Vo Tong, X. Biomass production of some legumes in the hilly area of Thinh Bien District, An Giang Province. In Proceedings on Sustainable Livestock Production on Local Feed Resources; Preston, R., Ogle, B., Eds.; HUAF-SAREC: Hue, Vietnam, 2003. [Google Scholar]
- Duguma, B.; Kang, B.T.; Okali, D.U.U. Effect of pruning intensities of three woody leguminous species grown in alley cropping with maize and cowpea on an alfisol. Agrofor. Syst. 1988, 6, 19–35. [Google Scholar] [CrossRef]
- Makhubedu, T.I.; Letty, B.A.; Mafongoya, P.L.; Scogings, P.F. Unraveling the Effects of Pruning Frequency on Biomass Productivity, Nonstructural Carbohydrates and Nitrogen Fixation Rates of Sesbania sesban. Forests 2022, 13, 2035. [Google Scholar] [CrossRef]
- Zapata-Campos, C.C.; García-Martínez, J.E.; Chavira, J.S.; Valdés, J.A.A.; Morales, M.A.M.; Mellado, M. Chemical composition and nutritional value of leaves and pods of Leucaena leucocephala, Prosopis laevigata and Acacia farnesiana in a xerophilous shrubland. Emir. J. Food Agric. 2020, 32, 723–730. [Google Scholar] [CrossRef]
- Sasu, P.; Attoh-Kotoku, V.; Akorli, D.E.; Adjei-Mensah, B.; Tankouano, R.A.; Kwaku, M. Nutritional evaluation of the leaves of Oxytenanthera abyssinica, Bambusa balcooa, Moringa oleifera, Terminalia catappa, Blighia sapida, and Mangifera indica as non-conventional green roughages for ruminants. J. Agric. Food Res. 2023, 11, 100466. [Google Scholar] [CrossRef]
- Sosa, R.E.E.; Perez, R.D.; Ortega, R.L.; Zapata, G.B. Tropical trees and shrubs forage potential for sheep feeding. Técnica Pecu. México 2004, 42, 129–144. [Google Scholar]
- Sarwatt, S.V.; Kapange, S.S.; Kakengi, A.M.V. Subsituting sunflower seed-cake with Moringa oleifera leaves as a supplemental goat feed in Tanzania. Agrofor. Syst. 2002, 56, 241–247. [Google Scholar] [CrossRef]
- Soliva, C.R.; Kreuzer, M.; Foidl, N.; Foidl, G.; Machmüller, A.; Hess, H.D. Feeding leaves for ruminants and their effects on ruminal fermentation in vitro. Anim. Feed. Sci. Technol. 2005, 118, 47–62. [Google Scholar] [CrossRef]
- Palm, C.A. Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agrofor. Syst. 1995, 30, 105–124. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Aihou, K.; Aman, S.; Tossah, B.K.; Diels, J.; Sanginga, N.; Merckx, R. Leaf quality of selected hedgerow species at two canopy ages in the derived savanna zone of West Africa. Agrofor. Syst. 2001, 53, 21–30. [Google Scholar] [CrossRef]
- Distel, R.A.; Arroquy, J.I.; Lagrange, S.; Villalba, J.J. Designing diverse agricultural pastures for improving ruminant production systems. Front. Sustain. Food Syst. 2020, 4, 596869. [Google Scholar] [CrossRef]
- Figueredo, E.S.; Rodrigues, R.C.; de Araújo, R.A.; dos Santos Costa, C.; de Sousa Santos, F.N.; da Silva, I.R.; Ribeiro de Jesus, A.P.; dos Santos Araújo, J.; da Silva Cabra, L.; Ribeiro Araújo, I.G. Maturity dependent variation in composition and characteristics of potentially digestible tissues of leucena. Sem. Ciênc. Agr. Londrina 2019, 40, 3133–3142. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Block | Stone | pH | N | C | P | Exch K | Exch Ca | Exch Mg |
|---|---|---|---|---|---|---|---|---|
| (%) | % | Mg kg−1 | ||||||
| I | 78 | 7.8 | 0.89 | 6.4 | 28 | 530 | 872 | 352 |
| II | 60 | 7.8 | 0.98 | 5.0 | 45 | 565 | 824 | 328 |
| III | 79 | 7.9 | 0.99 | 7.2 | 81 | 457 | 1077 | 310 |
| IV | 79 | 7.9 | 0.96 | 6.1 | 111 | 517 | 1573 | 388 |
| Mean | 74 | 7.8 | 0.95 | 6.2 | 66 | 517 | 1086 | 345 |
| Treatment | Survival (%) | ||
|---|---|---|---|
| 1 Month | 11 Months | 24 Months | |
| Mean | Mean | Mean | |
| Moringa | 91 a | 85 a | 69 a |
| Moringa (L) | 93 ab | 90 ab | 76 a |
| Guazuma | 97 ab | 95 ab | 88 a |
| Guazuma (L) | 97 ab | 96 ab | 80 a |
| Leucaena | 97 ab | 89 ab | 80 a |
| Leucaena (M) | 98 ab | 97 b | 81 a |
| Leucaena (G) | 100 b | 98 b | 86 a |
| SED | 2.5 | 3.4 | 6.8 |
| Species | Months after Planting | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 6 | 11 | |||||
| Height | Diameter | Height | Diameter | Height | Diameter | Height | Diameter | |
| Moringa | 36 b | 6 b | 38 ab | 9 bc | 84 | 18 | 180 | 29 |
| Moringa (L) | 31 b | 6 b | 38 ab | 9 bc | 83 | 17 | 193 | 28 |
| Guazuma | 17 a | 4 a | 22 a | 6 ab | 54 | 14 | 119 | 24 |
| Guazuma (L) | 18 a | 4 a | 26 ab | 6 ab | 62 | 13 | 131 | 21 |
| Leucaena | 43 b | 4 a | 47 b | 6 ab | 84 | 16 | 222 | 29 |
| Leucaena (M) | 32 b | 4 a | 33 ab | 5 a | 60 | 12 | 178 | 23 |
| Leucaena (G) | 35 b | 4 a | 41 ab | 6 ab | 79 | 14 | 201 | 24 |
| SED | 3.71 | 0.53 | 6.56 | 1.11 | 17.6 ns | 2.59 ns | 45.7 ns | 4.20 ns |
| Species | July | November | February | May | August | Total Գ |
|---|---|---|---|---|---|---|
| 1 | 2 * | 3 * | 4 * | 5 * | (kg ha−1) | |
| Moringa | 510 a | 440 a | 258 a | 457 a | 265 a | 1930 |
| Guazuma | 1167 ab | 1384 b | 259 a | 1332 a | 952 b | 5094 |
| Leucaena | 2456 bc | 1698 b | 1171 b | 1413 a | 1012 b | 7750 |
| Leucaena (M) | 1613 ab | 1329 b | 925 b | 1086 a | 918 b | 5871 |
| Leucaena (G) | 2911 bc | 1714 b | 1027 b | 1894 a | 1499 b | 9045 |
| Component | Content in Prunings (%) | ||||||
|---|---|---|---|---|---|---|---|
| N | CP | NDF | ADF | Pp | Tt | Ct | |
| Leaves | |||||||
| Leucaena | 3.6 | 23 | 36 | 21 | 5.3 | 4.2 | 2.7 |
| Leucaena (M) | 3.7 | 23 | 36 | 22 | 5.1 | 4.1 | 2.7 |
| Leucaena (G) | 3.5 | 22 | 35 | 21 | 5.2 | 4.3 | 2.3 |
| Moringa | 3.0 | 18 | 31 | 21 | 3.2 | 2.6 | 0.5 |
| Moringa (L) | 2.6 | 16 | 33 | 23 | 2.9 | 2.2 | 0.5 |
| Guazuma | 2.0 | 12 | 37 | 21 | 4.3 | 3.6 | 3.5 |
| Guazuma (L) | 2.4 | 15 | 36 | 21 | 4.0 | 3.4 | 3.1 |
| Twigs | |||||||
| Leucaena | 0.5 | 3.5 | 40 | 24 | 1.2 | 0.95 | 0.33 |
| Leucaena (M) | 0.6 | 4.0 | 40 | 25 | 0.2 | 0.18 | 0.05 |
| Leucaena (G) | 0.4 | 2.5 | 39 | 24 | 0.9 | 0.95 | 0.38 |
| Moringa | 0.6 | 3.8 | 37 | 26 | 0.8 | 0.70 | 0.00 |
| Moringa (L) | 0.2 | 0.8 | 34 | 25 | 0.7 | 0.60 | 0.05 |
| Guazuma | 0.2 | 1.8 | 39 | 24 | 0.2 | 0.07 | 0.15 |
| Guazuma (L) | 0.5 | 2.8 | 39 | 24 | 0.3 | 0.15 | 0.13 |
| Fodder * | |||||||
| Leucaena | 3.1 | 20 | 46 | 30 | 4.2 | 3.2 | 2.3 |
| Leucaena (M) | 3.1 | 20 | 45 | 30 | 4.9 | 3.9 | 2.7 |
| Leucaena (G) | 3.1 | 20 | 45 | 30 | 4.3 | 3.3 | 1.9 |
| Moringa | 2.4 | 15 | 41 | 29 | 2.5 | 1.9 | 0.5 |
| Moringa (L) | 2.5 | 16 | 40 | 30 | 2.2 | 1.6 | 0.5 |
| Guazuma | 1.8 | 11 | 47 | 31 | 4.1 | 3.5 | 3.7 |
| Guazuma (L) | 1.9 | 12 | 46 | 31 | 3.8 | 3.2 | 3.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzec-Gamboa, M.; Álvarez-Rivera, O.O.; Ramírez y Avilés, L.; Ku-Vera, J.; Solorio-Sánchez, F.J. Productivity and Nutritional Quality of Nitrogen-Fixing and Non-Fixing Shrub Species for Ruminant Production. Agronomy 2023, 13, 1089. https://doi.org/10.3390/agronomy13041089
Tzec-Gamboa M, Álvarez-Rivera OO, Ramírez y Avilés L, Ku-Vera J, Solorio-Sánchez FJ. Productivity and Nutritional Quality of Nitrogen-Fixing and Non-Fixing Shrub Species for Ruminant Production. Agronomy. 2023; 13(4):1089. https://doi.org/10.3390/agronomy13041089
Chicago/Turabian StyleTzec-Gamboa, Magnolia, Oscar Omar Álvarez-Rivera, Luis Ramírez y Avilés, Juan Ku-Vera, and Francisco Javier Solorio-Sánchez. 2023. "Productivity and Nutritional Quality of Nitrogen-Fixing and Non-Fixing Shrub Species for Ruminant Production" Agronomy 13, no. 4: 1089. https://doi.org/10.3390/agronomy13041089