



Outcome of Microalgae Biomass Application on Seed Germination and Hormonal Activity in Winter Wheat Leaves
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
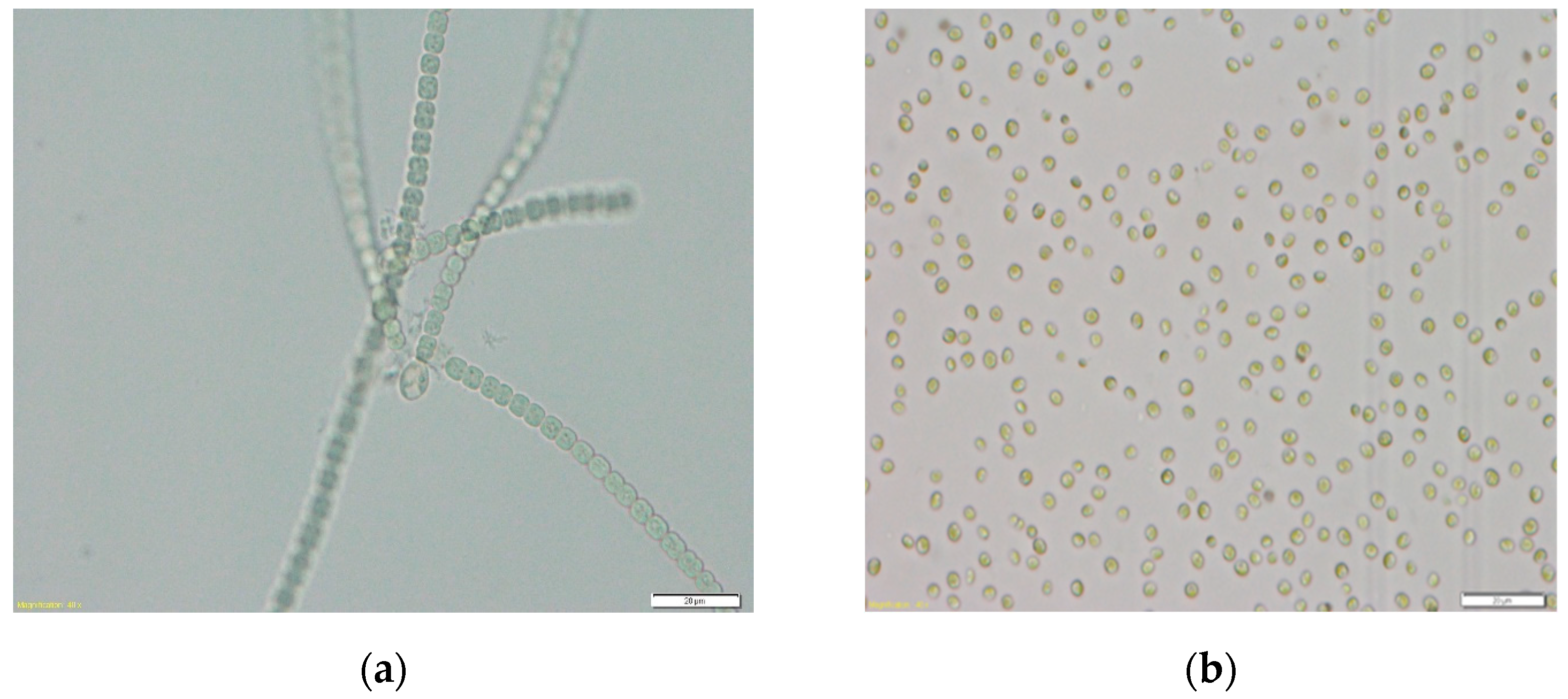
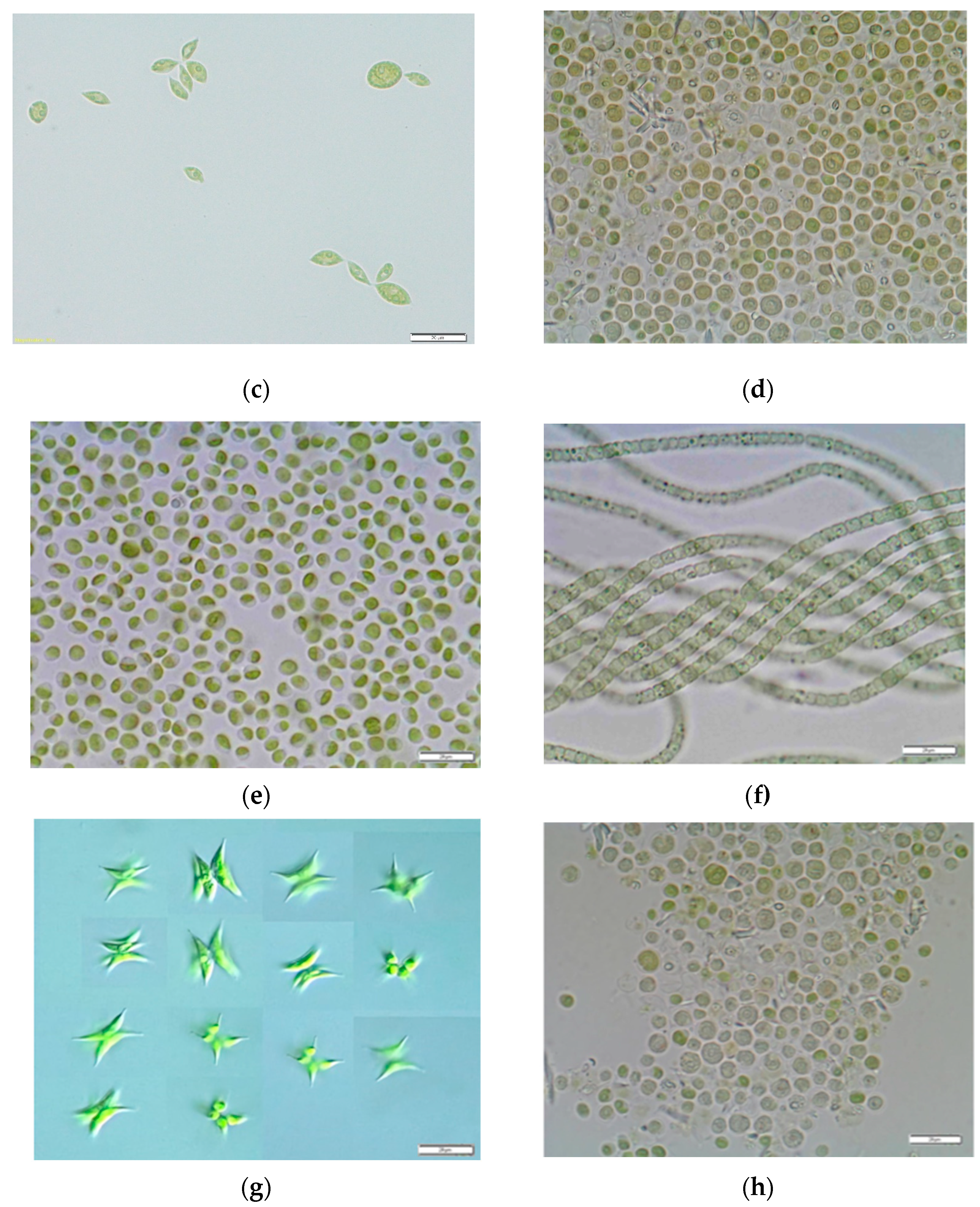
2.1. Growth Conditions of Algae Cultures and Preparation of Microalgae Biomass
2.2. Germination Test
2.3. Mungbean Rooting Bioassay
2.4. Growth Conditions of Wheat Plants and Microalgal Treatments
2.5. Photosynthesis Measurements
2.6. Metabolomics Analyses
2.7. Statistical Analyses
3. Results
3.1. Germination Bioassay
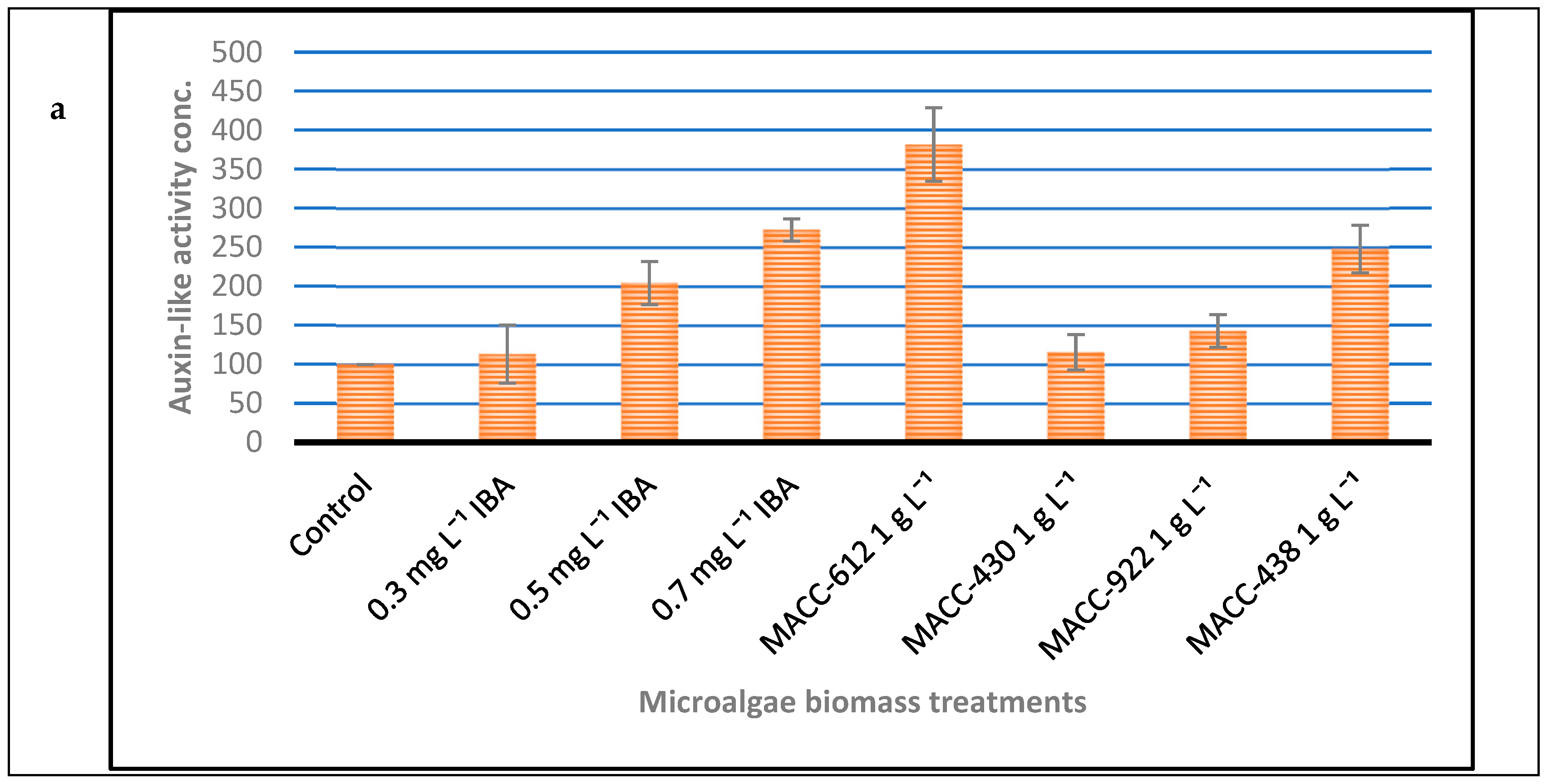
3.2. Mungbean Rooting Bioassay
3.3. Metabolite Characterization of the Microalgae Strains
3.4. Effects of Algae Strains on Photosynthesis and Metabolite Contents of Wheat
3.5. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, L.V.; Trewavas, A.J. Is algal development controlled by plant growth substances? J. Phycol. 1991, 27, 322–326. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ghareib, H.R. Bioactivity of Ulva lactuca L. acetone extract on germination and growth of lettuce and tomato plants. Afr. J. Biotechnol. 2009, 8, 3832–3838. [Google Scholar]
- Li, Y.-X.; Wijesekaraa, I.; Li, Y.; Kim, S.-K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Tarkowská, D.; Novák, O.; Strnad, M.; Ördög, V.; Van Staden, J. Hormone profiles in microalgae: Gibberellins and brassinosteroids. Plant Physiol. Biochem. 2013, 70, 348–353. [Google Scholar] [CrossRef]
- Potin, P. Oxidative burst and related responses in biotic interactions of algae. In Algal Chemical Ecology; Amsler, C.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; p. 245. [Google Scholar]
- Plaza, B.M.; Gómez-Serrano, C.; Acién-Fernández, F.G.; Jimenez-Becker, S. Effect of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp.) on Florida × hybrida growth. J. Appl. Phycol. 2018, 30, 2359–2365. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella Vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Escapa, C.; Coimbra, R.N.; Paniagua, S.; García, A.I.; Otero, M. Nutrients and pharmaceuticals removal from wastewater by culture and harvesting of Chlorella sorokiniana. Biores. Technol. 2015, 185, 276–284. [Google Scholar] [CrossRef]
- Escapa, C.; Coimbra, R.N.; Paniagua, S.; García, A.I.; Otero, M. Paracetamol and salicylic acid removal from contaminated water by microalgae. J. Environ. Manag. 2017, 20, 799–806. [Google Scholar] [CrossRef]
- Shaaban, M.M. Green microalgae water extract as foliar feeding to wheat plants. Pak. J. Biol. Sci. 2001, 4, 628–632. [Google Scholar] [CrossRef] [Green Version]
- Takács, G.; Stirk, W.A.; Gergely, I.; Molnár, Z.; Van Staden, J.; Ördög, V. Biostimulating effects of the cyanobacterium Nostoc piscinale on winter wheat in field experiments. S. Afr. J. Bot. 2019, 126, 99–106. [Google Scholar] [CrossRef]
- Waterman, P.G.; Mole, S. Analysis of Phenolic Plant Metabolites In-Methods in Ecology; Blackwell Scientific Publications: Oxford, UK, 1994. [Google Scholar]
- Tamiya, H. Mass culture of algae. Annu. Rev. Plant Physiol. 1957, 8, 309–334. [Google Scholar] [CrossRef]
- Ördög, V. Apparatus for laboratory algal bioassay. Int. Rev. Der Gesamten Hydrobiol. 1982, 67, 127–136. [Google Scholar]
- Hess, C.E. The mung bean bioassay for the detection of root promotion substances. Plant Physiol. 1961, 36, XXI. [Google Scholar]
- Darko, E.; Khalil, R.; Dobi, Z.; Kovács, V.; Szalai, G.; Janda, T.; Molnár, I. The addition of Aegilops biuncialis chromosomes 2M or 3M improves the salt tolerance of wheat in different ways. Sci. Rep. 2020, 10, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Majláth, I.; Éva, C.; Tajti, J.; Khalil, R.; Elsayed, N.; Darko, E.; Szalai, G.; Janda, T. Exogenous methylglyoxal enhances the reactive aldehyde detoxification capability and frost-hardiness of wheat. Plant Physiol. Biochem. 2020, 149, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields were calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Hrdlička, J.; Gucký, T.; Novák, O.; Kulkarni, M.; Gupta, S.; Van Staden, J.; Doležal, K. Quantification of karrikins in smoke water using ultra-high-performance liquid chromatography-tandem mass spectrometry. Plant Methods 2019, 15, 81. [Google Scholar] [CrossRef]
- Novák, O.; Napier, R.; Ljung, K. Zooming in on plant hormone analysis: Tissue- and cell-specific approaches. Annu. Rev. Plant Biol. 2017, 68, 323–348. [Google Scholar] [CrossRef]
- Janda, T.; Gondor, O.; Yordanova, R.; Szalai, G.; Pál, M. Salicylic acid, and photosynthesis: Signaling and effects. Acta Physiol. Plant 2014, 36, 2537–2546. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Czékus, Z.; Tari, I.; Ördög, A. The Multifaceted Roles of Plant Hormone Salicylic Acid in Endoplasmic Reticulum Stress and Unfolded Protein Response. Int. J. Mol. Sci. 2019, 20, 5842. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Borbély, P.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of salicylic acid on photosynthetic activity and chloroplast morphology under light and prolonged darkness. Photosynthetica 2019, 57, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Janda, T.; Majláth, I.; Szalai, G. Involvement of Salicylic Acid and Other Phenolic Compounds in Light-Dependent Cold Acclimation in Maize. Int. J. Mol. Sci. 2020, 21, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pál, M.; Szalai, G.; Lantos, E.; Nagyéri, G.; Janda, T. Comparative study of salicylic acid contents in young wheat and rice plants and their anticancer activities in HepG2 and Caco-2 cells. Biologia Fut. 2020, 71, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Filgueiras, C.C.; Martins, A.D.; Pereira, R.V.; Willett, D.S. The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture. Int. J. Mol. Sci. 2019, 20, 5851. [Google Scholar] [CrossRef] [Green Version]
- Pokotylo, I.; Kravets, V.; Ruelland, E. Salicylic Acid Binding Proteins (SABPs): The Hidden Forefront of Salicylic Acid Signalling. Int. J. Mol. Sci. 2019, 20, 4377. [Google Scholar] [CrossRef] [Green Version]
- Tajti, J.; Hamow, K.A.; Majláth, I.; Gierczik, K.; Németh, E.; Janda, T.; Pál, M. Polyamine-Induced Hormonal Changes in eds5 and sid2 Mutant Arabidopsis Plants. Int. J. Mol. Sci. 2019, 20, 5746. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Fariduddin, Q.; Janda, T. Multifaceted Role of Salicylic Acid in Combating Cold Stress in Plants: A Review. J. Plant Growth Regul. 2020, 40, 464–485. [Google Scholar] [CrossRef]
- Nadarajah, K.; Abdul Hamid, N.W.; Abdul Rahman, N. SA-Mediated Regulation and Control of Abiotic Stress Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 5591. [Google Scholar] [CrossRef]
- Ranglova, K.; Lakatos, E.G.; Manoel, J.A.C.; Grivalský, T.; Estrella, F.S.; Fernandez, F.G.A.; Molnar, Z.; Ördög, V.; Masojídek, J. Growth, biostimulant, and biopesticide activity of the MACC-1 Chlorella strain cultivated outdoors in inorganic medium and wastewater. Algal Res. 2021, 53, 102136. [Google Scholar] [CrossRef]
- Navarro-López, E.; Ruíz-Nieto, A.; Ferreira, A.; Acién, F.G.; Gouveia, L. Biostimulant Potential of Scenedesmus obliquus Grown in Brewery Wastewater. Molecules 2020, 25, 664. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-J.; Yang, M.F.; Chen, H.; Qu, L.-Q.; Chen, F.; Shen, S.-H. Abscisic acid pretreatment enhances salt tolerance of rice seedlings: Proteomic evidence. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, L.; Forde, B.G.; Davies, W.J. The Biphasic Root Growth Response to Abscisic Acid in Arabidopsis Involves Interaction with Ethylene and Auxin Signalling Pathways. Front. Plant Sci. 2017, 8, 1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, S.; Rodriguez, J.; Evans, S.E.; Davies, W. Root and shoot growth of plants treated with abscisic acid. Ann. Bot. 1981, 47, 595–602. [Google Scholar] [CrossRef]
- De Smet, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, K.; Tao, H.; Wen, X.; Geng, Y.; Li, Y. Enhanced growth and lipid production of Chlorella pyrenoidosa by plant growth regulator GA3. Fresenius Environ. Bull. 2015, 24, 3414–3419. [Google Scholar]
- Reigosa, M.J.; Pazos-Malvido, E. Phytotoxic effects of 21 Plant secondary metabolites on Arabidopsis thaliana germination and root growth. J. Chem. Ecol. 2007, 33, 1456–1466. [Google Scholar] [CrossRef]
- Wu, L.; Guo, X.; Harivandi, M.A. Allelopathic effects of phenolic acids detected in buffalograss (Buchloe dactyloides) clippings on the growth of annual bluegrass (Poa annua) and buffalograss seedlings. Environ. Exp. Bot. 1998, 39, 159–167. [Google Scholar] [CrossRef]
- Toribio, A.J.; Jurado, M.M.; Suárez-Estrella, F.; López-González, J.A.; Martínez-Gallardo, M.R.; López, M.J. Application of sonicated extracts of cyanobacteria and microalgae for the mitigation of bacterial canker in tomato seedlings. J. Appl. Phycol. 2021, 33, 3817–3829. [Google Scholar] [CrossRef]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon 2021, 7, 5908. [Google Scholar] [CrossRef]
- Gondor, O.K.; Janda, T.; Soos, V.; Pal, M.; Majlath, I.; Adak, M.K.; Balazs, E.; Szalai, G. Salicylic Acid Induction of Flavonoid Biosynthesis Pathways in Wheat Varies by Treatment. Front. Plant Sci. 2016, 7, 1447. [Google Scholar] [CrossRef] [Green Version]
- Szalai, G.; Pál, M.; Árendás, T.; Janda, T. Priming seed with salicylic acid increases grain yield and modifies polyamine levels in maize. Cereal Res. Commun. 2016, 44, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Szalai, G.; Horgosi, S.; Soós, V.; Majláth, I.; Balázs, E.; Janda, T. Salicylic acid treatment of pea seeds induces its de novo synthesis. J. Plant Physiol. 2011, 168, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic acid perception and signaling: Structural mechanisms and applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and Mechanism of Jasmonic Acid in Plant Responses to Abiotic and Biotic Stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Effects of jasmonic acid signaling on the wheat microbiome differ between body sites. Sci. Rep. 2017, 7, 41766. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Germination Index | Germination Rate Index | Speed of Germination | Radicle Length on 7th Day (cm) |
|---|---|---|---|---|
| MACC-430@ 0.1 gL−1 | 53 | 45.79 | 4.58 | 1.1 |
| MACC-430@ 0.3gL−1 | 78 | 62.29 | 6.23 | 1.1 |
| MACC-430@ 0.5 gL−1 | 65 | 53.79 | 5.38 | 1.2 |
| MACC-430@ 1 gL−1 | 87 | 68.12 | 6.81 | 1.2 |
| MACC-683@ 0.1 gL−1 | 48 | 42.95 | 4.30 | 1.0 |
| MACC-683@ 0.3 gL−1 | 61 | 51.45 | 5.15 | 0.9 |
| MACC-683@ 0.5 gL−1 | 74 | 59.62 | 5.96 | 1.2 |
| MACC-683@ 1 gL−1 | 77 | 61.79 | 6.18 | 1.2 |
| MACC-922@ 0.1 gL−1 | 68 | 55.79 | 5.58 | 1.2 |
| MACC-922@ 0.3 gL−1 | 74 | 59.62 | 5.96 | 0.9 |
| MACC-922@ 0.5 gL−1 | 62 | 51.95 | 5.20 | 0.9 |
| MACC-922@ 1 gL−1 | 70 | 57.29 | 5.73 | 0.8 |
| MACC-519@ 0.1 gL−1 | 66 | 54.12 | 5.41 | 0.8 |
| MACC-519@ 0.3 gL−1 | 62 | 51.95 | 5.20 | 0.8 |
| MACC-519@ 0.5 gL−1 | 54 | 46.95 | 4.70 | 0.6 |
| MACC-519@ 1 gL−1 | 57 | 48.95 | 4.90 | 0.7 |
| MACC-612@ 0.1 gL−1 | 75 | 60.29 | 6.03 | 1.2 |
| MACC-612@ 0.3 gL−1 | 78 | 62.29 | 6.23 | 1.1 |
| MACC-612@ 0.5 gL−1 | 77 | 61.45 | 6.15 | 1.0 |
| MACC-612@ 1 gL−1 | 62 | 51.95 | 5.20 | 1.1 |
| MACC-677@ 0.1 gL−1 | 57 | 48.29 | 4.83 | 0.7 |
| MACC-677@ 0.3 gL−1 | 61 | 51.12 | 5.11 | 0.7 |
| MACC-677@ 0.5 gL−1 | 56 | 47.79 | 4.78 | 0.6 |
| MACC-677@ 1 gL−1 | 55 | 47.45 | 4.75 | 0.7 |
| MACC-755@ 0.1 gL−1 | 61 | 51.45 | 5.15 | 0.5 |
| MACC-755@ 0.3 gL−1 | 58 | 49.45 | 4.95 | 0.5 |
| MACC-755@ 0.5 gL−1 | 64 | 53.45 | 5.35 | 0.6 |
| MACC-755@ 1 gL−1 | 39 | 36.95 | 3.70 | 0.5 |
| MACC-438@ 0.1 gL−1 | 36 | 34.95 | 3.50 | 0.3 |
| MACC-438@ 0.3 gL−1 | 34 | 33.29 | 3.33 | 0.2 |
| MACC-438@ 0.5 gL−1 | 32 | 31.62 | 3.16 | 0.2 |
| MACC-438@ 1 gL−1 | 29 | 29.62 | 2.96 | 0.3 |
| CONTROL | 45 | 40.12 | 4.01 | 0.8 |
| CD @1% | 4.5396 | 3.926 | 0.9246 | 0.1444 |
| SE d | 1.7115 | 0.987 | 0.3486 | 0.0544 |
| IAA | SA | pHBA | BA | tCA | |
|---|---|---|---|---|---|
| MACC-438 | n.d. | 21.1 ± 1.3 bc | 86.3 ± 6.6 b | 798.3 ± 84.9 b | n.d. |
| MACC-430 | n.d. | 22.5 ± 0.7 b | 65.8 ± 4.2 c | 971.7 ± 55.4 b | n.d. |
| MACC-612 | 59.3 ± 2.6 | 35.1 ± 1.9 a | 698.3 ± 27.8 a | 2210.0 ± 63.8 a | 38.2 ± 1.0 |
| MACC-922 | n.d. | 14.6 ± 3.5 c | 80.7 ± 3.7 b | 883.3 ± 48.7 b | n.d. |
| Control | MACC-922 | MACC-430 | MACC-612 | MACC-438 | |
|---|---|---|---|---|---|
| Fv/Fm | 0.784 ± 0.01 | 0.788 ± 0.006 | 0.786 ± 0.004 | 0.780 ± 0.011 | 0.778 ± 0.013 |
| Y (II) | 0.541 ± 0.04 | 0.556 ± 0.022 | 0.550 ± 0.020 | 0.530 ± 0.029 | 0.535 ± 0.039 |
| Y(NO) | 0.228 ± 0.03 | 0.219 ± 0.041 | 0.242 ± 0.025 | 0.250 ± 0.033 | 0.253 ± 0.028 |
| Pn | 12.8 ± 2.4 | 14.9 ± 3.9 | 12.6 ± 2.2 | 14.4 ± 1.2 | 9.3 ± 1.2 |
| gs | 337 ± 157 | 525 ± 183 | 358 ± 98 | 438 ± 54 | 246 ± 51 |
| Ci | 195 ± 33 | 210 ± 17 | 202 ± 16 | 200 ± 17 | 197 ± 8 |
| E | 3.85 ± 1.09 | 5.05 ± 0.90 | 4.05 ± 0.67 | 4.68 ± 0.37 | 3.20 ± 0.49 |
| WUE | 3.42 ± 0.49 | 2.95 ± 0.64 | 3.11 ± 0.21 | 3.09 ± 0.29 | 3.20 ± 0.49 |
| Indole-3-acetic-Acid | Salicylic Acid | P-Hydroxybenzoic Acid | p-Coumaric Acid | Jamonic Acid | Abscisic Acid | Neochlorogenic Acid | Rutin | Naringenin | Jasmonic Acid-Leucine/Isoleucine Conjugate | |
|---|---|---|---|---|---|---|---|---|---|---|
| Béres1 | ||||||||||
| Control | 1.03 ±0.21 | 47.2 ±6.5 | 25.9 ±1.0 | 19.7 ±5.0 | 106.4 ±11.5 | 3.92 ±0.26 | 118 ±65 | 27.7 ±3.7 | 1.28 ±0.20 | 50,194 ±7023 |
| 430 | 0.84 ±0.13 | 50.3 ±11.9 | 29.2 ±1.5 * | 16.4 ±4.1 | 62.8 ±12.8 * | 4.36 ±0.30 | 178 ±76 | 30.3 ±2.6 | 1.27 ±0.19 | 21,850 ±3644 * |
| 438 | 1.02 ±0.22 | 49.1 ±10.1 | 29.3 ±2.5 * | 29.0 ±11.4 | 72.8 ±15.4 * | 3.83 ±0.29 | 105 ±17 | 31.9 ±4.2 | 1.61 ±0.36 | 27,942 ±5796 * |
| 612 | 0.99 ±0.20 | 45.8 ±12.8 | 27.6 ±3.2 | 22.9 ±5.8 | 40.4 ±23.7 * | 4.05 ±0.81 | 191 ±57 | 28.4 ±2.2 | 1.97 ±1.28 | 15,112 ±12,476 * |
| 922 | 1.05 ±0.21 | 64.2 ±12.3 * | 26.6 ±2.7 | 18.8 ±3.5 | 57.1 ±11.3 * | 4.20 ±0.50 | 129 ±80 | 31.5 ±3.5 | 1.71 ±0.31 * | 19,417 ±5716 * |
| Nádor | ||||||||||
| Control | 1.04 ±0.20 | 55.8 ±8.7 | 20.5 ±5.4 | 32.7 ±5.1 | 18.1 ±2.9 | 15.24 ±0.73 | 1271 ±405 | 3.4 ±0.8 | 3.94 ±1.40 | 9078 ±1267 |
| 430 | 0.78 ±0.13 | 44.8 ±5.0 | 18.1 ±2.5 | 27.4 ±8.2 | 22.9 ±4.9 | 8.61 ±2.31 * | 1064 ±42 | 3.3 ±0.8 | 3.13 ±0.33 | 7405 ±2070 |
| 438 | 0.75 ±0.04 * | 115.4 ±29.3 * | 15.3 ±2.0 | 25.8 ±1.5 * | 59.5 ±20.1 * | 14.12 ±1.05 | 476 ±157 * | 3.4 ±0.5 | 2.38 ±0.39 | 17,980 ±5702 * |
| 612 | 0.78 ±0.06 * | 50.8 ±9.1 | 19.6 ±0.4 | 29.2 ±5.1 | 41.4 ±7.8 * | 9.56 ±0.85 * | 522 ±277 * | 3.4 ±0.3 | 3.09 ±0.37 | 11,842 ±2164 |
| 922 | 0.69 ±0.06 * | 67.0 ±6.3 | 19.1 ±3.1 | 26.4 ±3.6 | 20.9 ±3.2 | 7.34 ±0.30 * | 760 ±155 * | 3.5 ±0.5 | 2.82 ±0.35 | 9934 ±1857 |
| Béres2 | ||||||||||
| Control | 0.98 ±0.12 | 142.2 ±8.7 | 19.8 ±1.1 | 14.0 ±1.4 | 17.8 ±3.2 | 6.51 ±0.42 | 302 ±101 | 22.7 ±5.3 | 2.26 ±0.25 | 5709 ±1081 |
| 430 | 1.05 ±0.11 | 114.3 ±29.3 | 25.1 ±5.4 | 16.8 ±2.6 | 16.7 ±4.3 | 7.28 ±0.22 * | 481 ±285 | 24.4 ±8.6 | 3.12 ±0.83 | 4873 ±1436 |
| 438 | 0.99 ±0.03 | 98.7 ±14.6 * | 19.4 ±1.2 | 14.6 ±5.1 | 12.4 ±3.2 * | 6.75 ±0.36 | 605 ±319 | 19.9 ±6.8 | 2.23 ±0.21 | 3782 ±829 * |
| 612 | 0.94 ±0.12 | 91.1 ±18.2 | 23.5 ±2.6 | 15.0 ±3.1 | 16.5 ±5.2 | 5.97 ±0.18 | 115 ±32 * | 16.7 ±2.8 | 2.60 ±0.33 | 4202 ±1266 |
| 922 | 0.85 ±0.11 | 93.8 ±18.7 * | 18.3 ±1.1 * | 12.3 ±1.8 | 17.1 ±5.9 | 6.56 ±0.30 | 399 ±107 | 21.4 ±2.6 | 2.02 ±0.11 | 7795 ±4907 |
| Salicylic Acid | p-Hydroxybenzoic Acid | Benzoic Acid | |
|---|---|---|---|
| Germination index | 0.262 NS | 0.101 NS | 0.303 NS |
| Radicle length | 0.415 NS | 0.388 NS | 0.412 NS |
| Net photosynthesis | 0.045 NS | 0.384 NS | 0.345 NS |
| Stomatal conductivity | 0.140 NS | 0.315 NS | 0.370 NS |
| Intercellular CO2 | 0.178 NS | −0.010 NS | 0.244 NS |
| Transpiration | 0.215 NS | 0.407 NS | 0.450 NS |
| Salicylic Acid | p-Hydroxybenzoic Acid | Benzoic Acid | |
|---|---|---|---|
| Indole-3-acetic acid | |||
| Béres1 | −0.345 NS | 0.013 NS | −0.184 NS |
| Nádor | −0.660 NS | −0.231 NS | −0.560 NS |
| Béres2 | 0.043 NS | −0.190 NS | −0.169 NS |
| Abscisic acid | |||
| Béres1 | 0.226 NS | −0.028 NS | 0.224 NS |
| Nádor | −0.500 NS | −0.289 NS | −0.552 NS |
| Béres2 | −0.229 NS | −0.732 NS | −0.478 NS |
| Jasmonic acid | |||
| Béres1 | −0.903 * | −0.707 NS | −0.917 * |
| Nádor | 0.544 NS | 0.341 NS | 0.382 NS |
| Béres2 | −0.330 NS | 0.033 NS | 0.089 NS |
| Rutin | |||
| Béres1 | 0.172 NS | −0.362 NS | −0.035 NS |
| Nádor | −0.218 NS | −0.018 NS | −0.039 NS |
| Béres2 | −0.625 NS | −0.847 NS | −0.734 NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutum, L.; Janda, T.; Darkó, É.; Szalai, G.; Hamow, K.Á.; Molnár, Z. Outcome of Microalgae Biomass Application on Seed Germination and Hormonal Activity in Winter Wheat Leaves. Agronomy 2023, 13, 1088. https://doi.org/10.3390/agronomy13041088
Mutum L, Janda T, Darkó É, Szalai G, Hamow KÁ, Molnár Z. Outcome of Microalgae Biomass Application on Seed Germination and Hormonal Activity in Winter Wheat Leaves. Agronomy. 2023; 13(4):1088. https://doi.org/10.3390/agronomy13041088
Chicago/Turabian StyleMutum, Lamnganbi, Tibor Janda, Éva Darkó, Gabriella Szalai, Kamirán Áron Hamow, and Zoltán Molnár. 2023. "Outcome of Microalgae Biomass Application on Seed Germination and Hormonal Activity in Winter Wheat Leaves" Agronomy 13, no. 4: 1088. https://doi.org/10.3390/agronomy13041088