

Effect of NaCl on Morphophysiological and Biochemical Responses in Gossypium hirsutum L.

Abstract
:1. Introduction
2. Materials and Methodology
2.1. Collection of Germplasms
2.2. Experimental Design and Salt Treatment
2.3. Harvesting and Transplanting
2.4. Analysis of Morphophysiological Parameters
Na+ and K+ Contents
2.5. Analysis of Biochemical Parameters
2.5.1. Determination of Proline
2.5.2. Catalase Activity
2.5.3. Peroxidase Activity
2.5.4. Hydrogen Peroxide
2.6. Statistical Analysis
3. Results
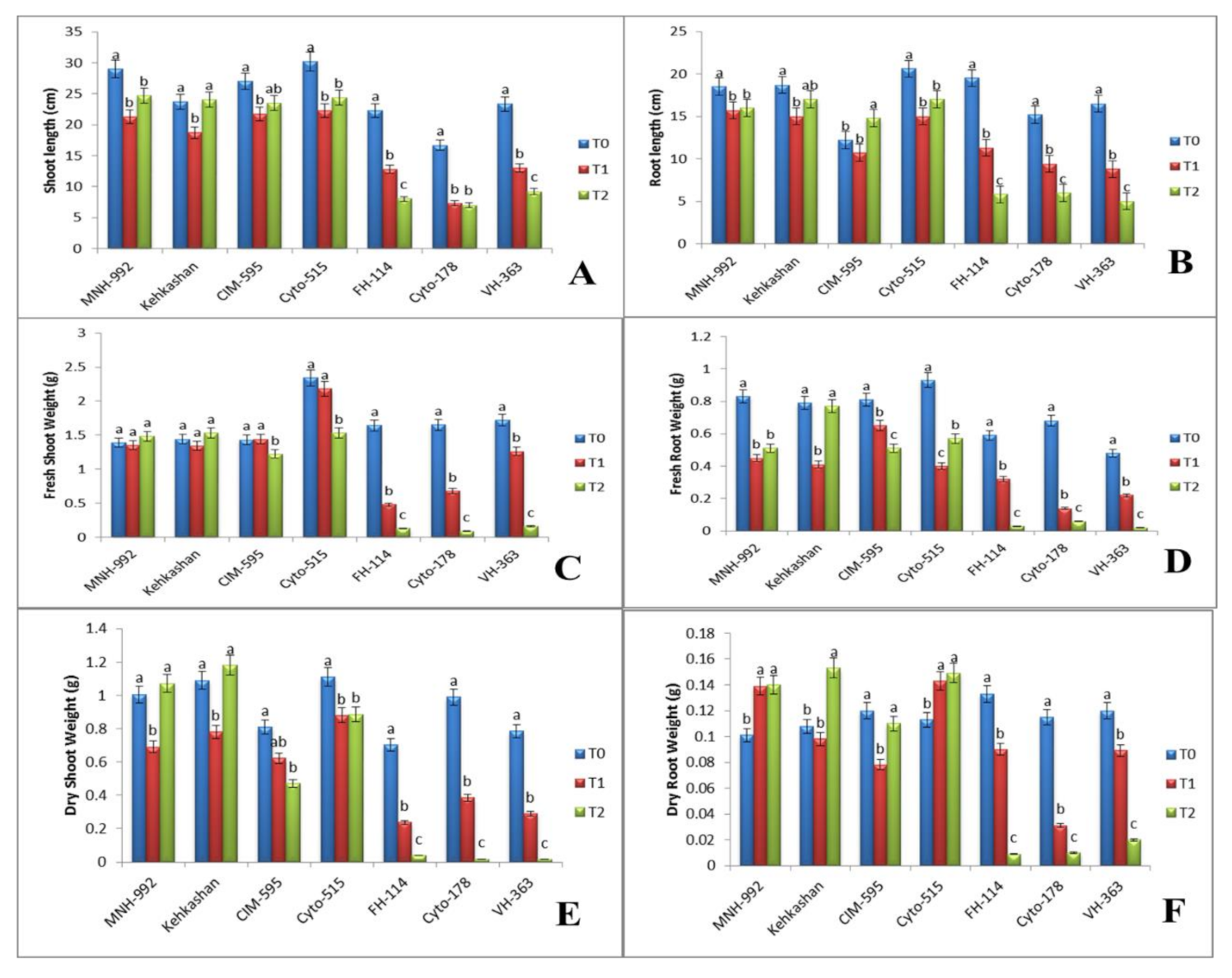
3.1. Effect of Salt Stress on Morphological Properties
3.1.1. Shoot Length (cm)
3.1.2. Root Length (cm)
3.1.3. Fresh Shoot Weight (g)
3.1.4. Fresh Root Weight (g)
3.1.5. Dry Shoot Weight (g)
3.1.6. Dry Root Weight (g)
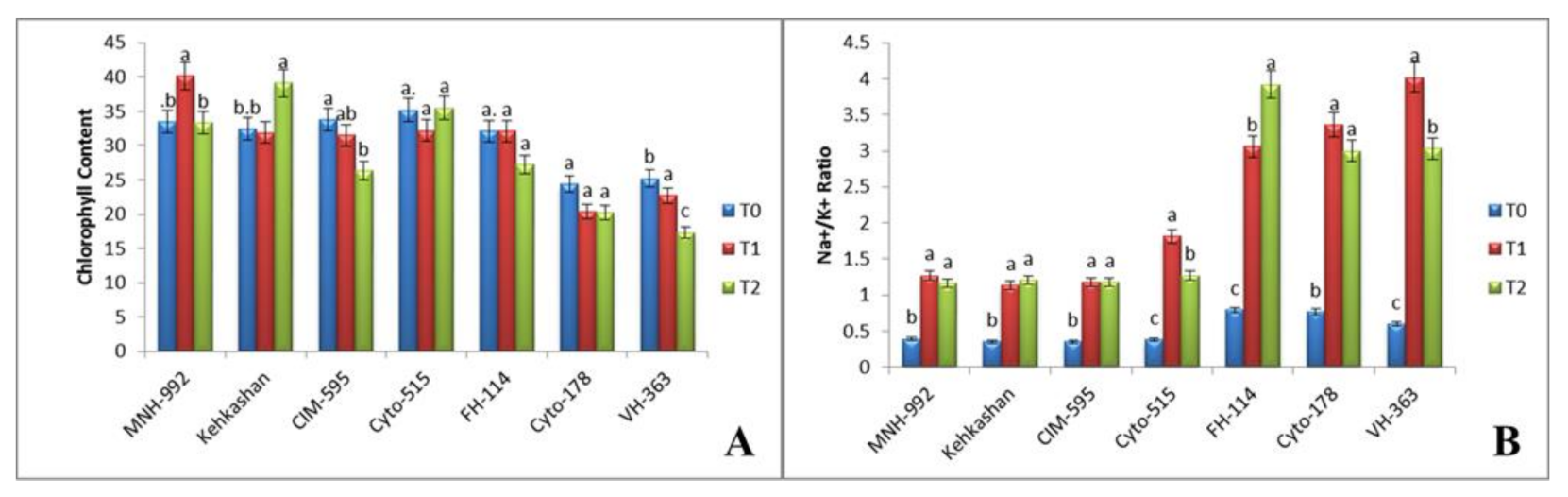
3.2. Effect of Salt Stress on Physiological Properties
3.2.1. Chlorophyll Content
3.2.2. Na+/K+ Ratio
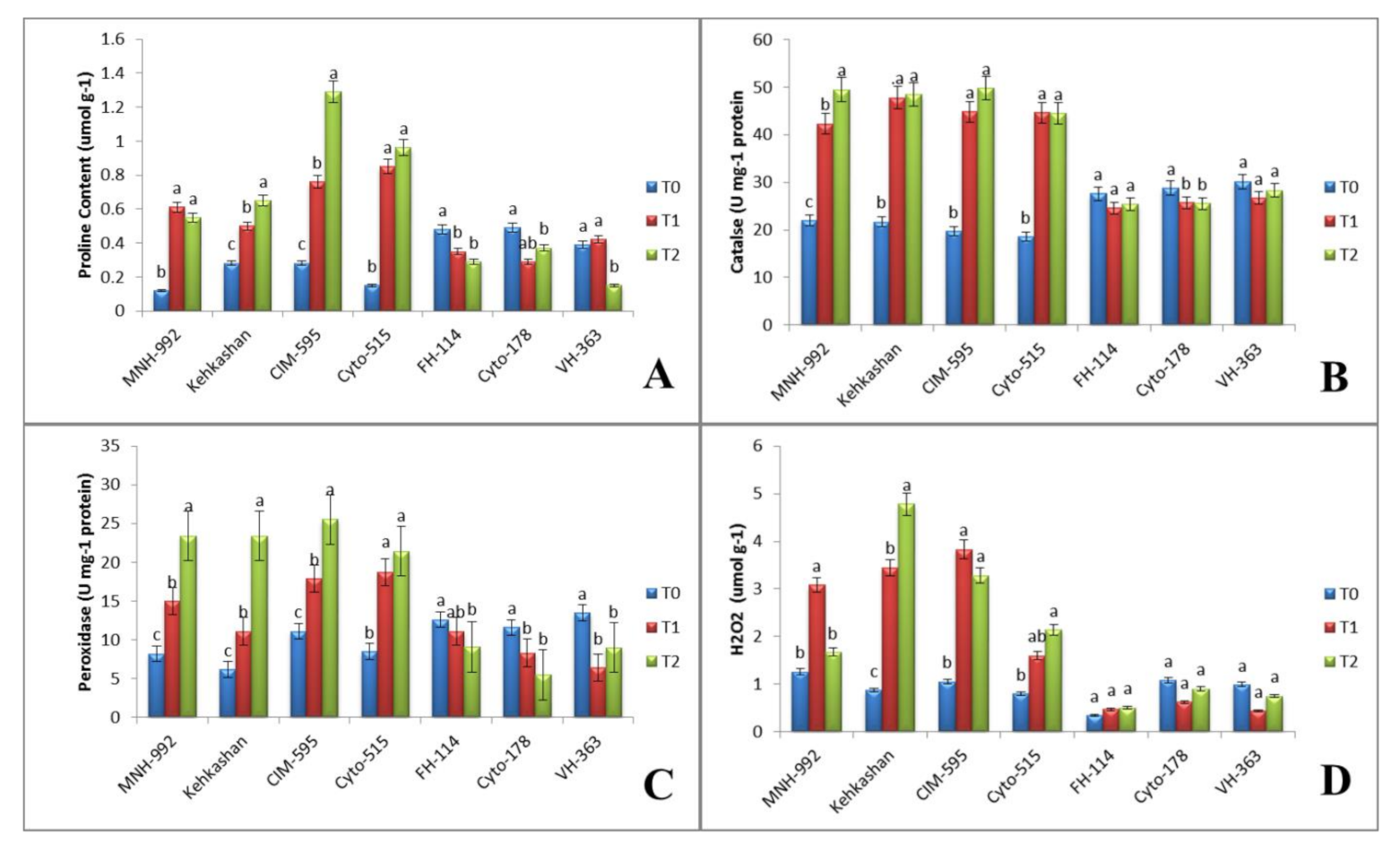
3.3. Effect of Salt Stress on Biochemical Properties
3.3.1. Proline Content
3.3.2. Catalase Activity
3.3.3. Peroxidase Activity
3.3.4. Hydrogen Peroxidase
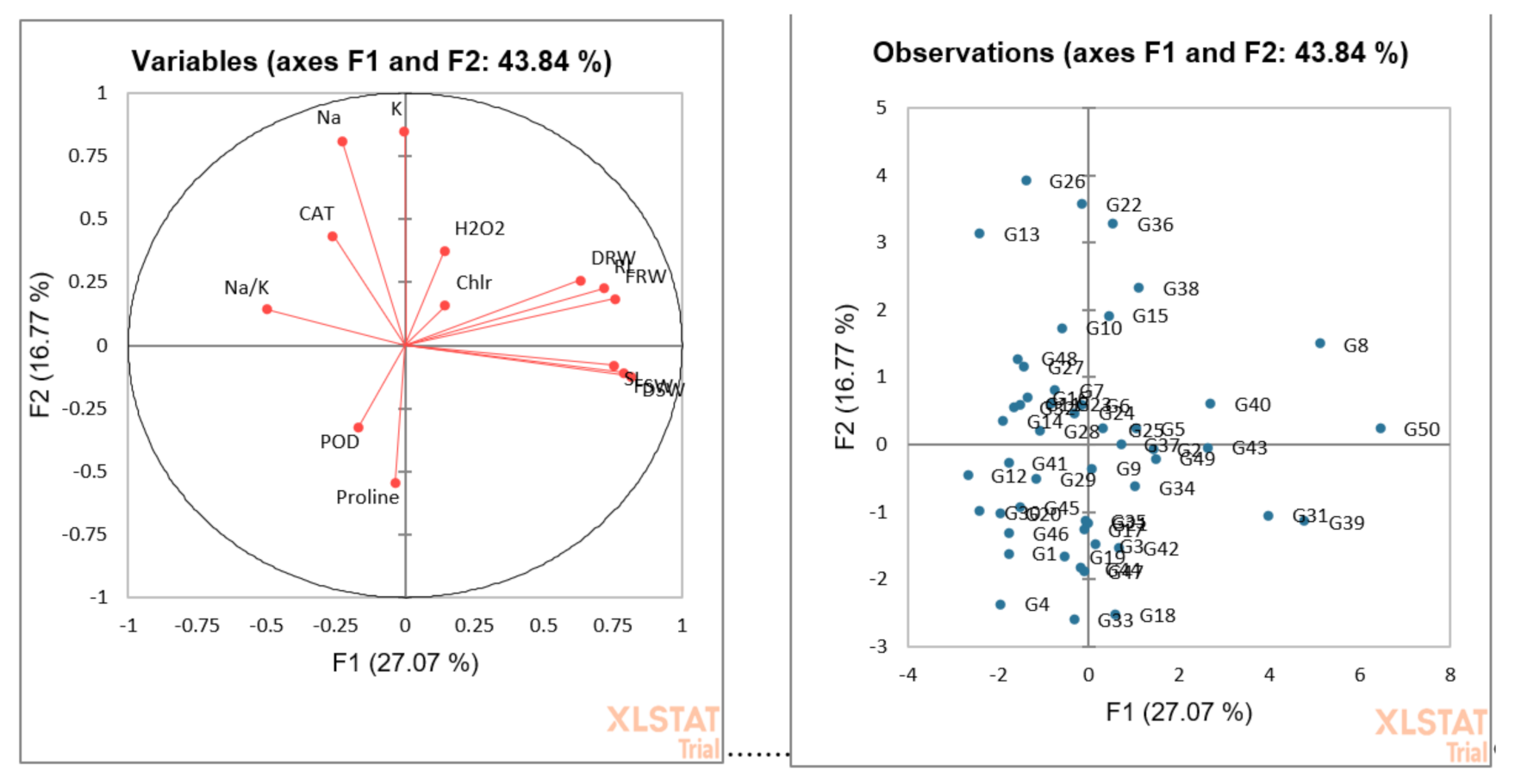
3.4. Cluster Analysis
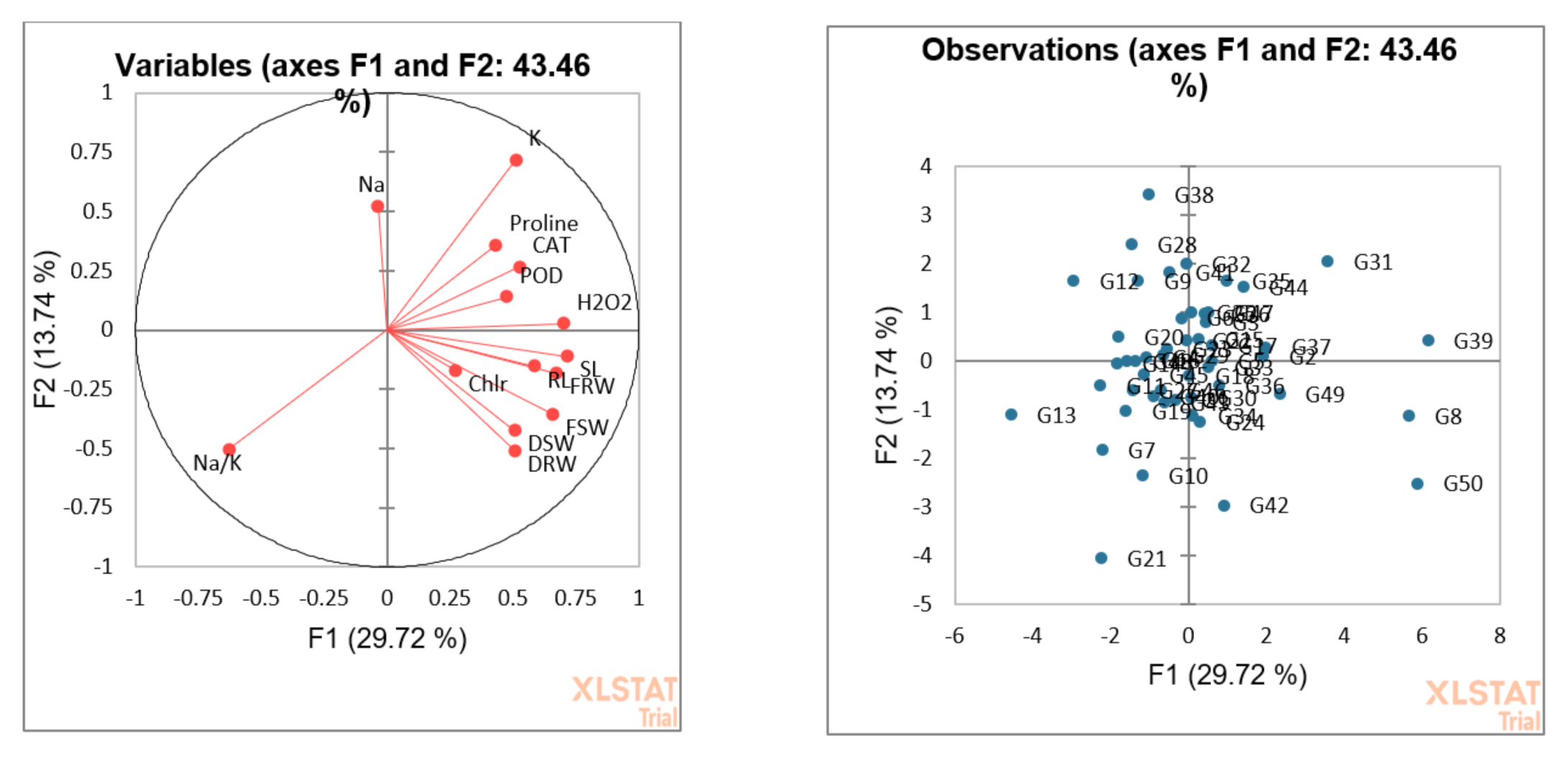
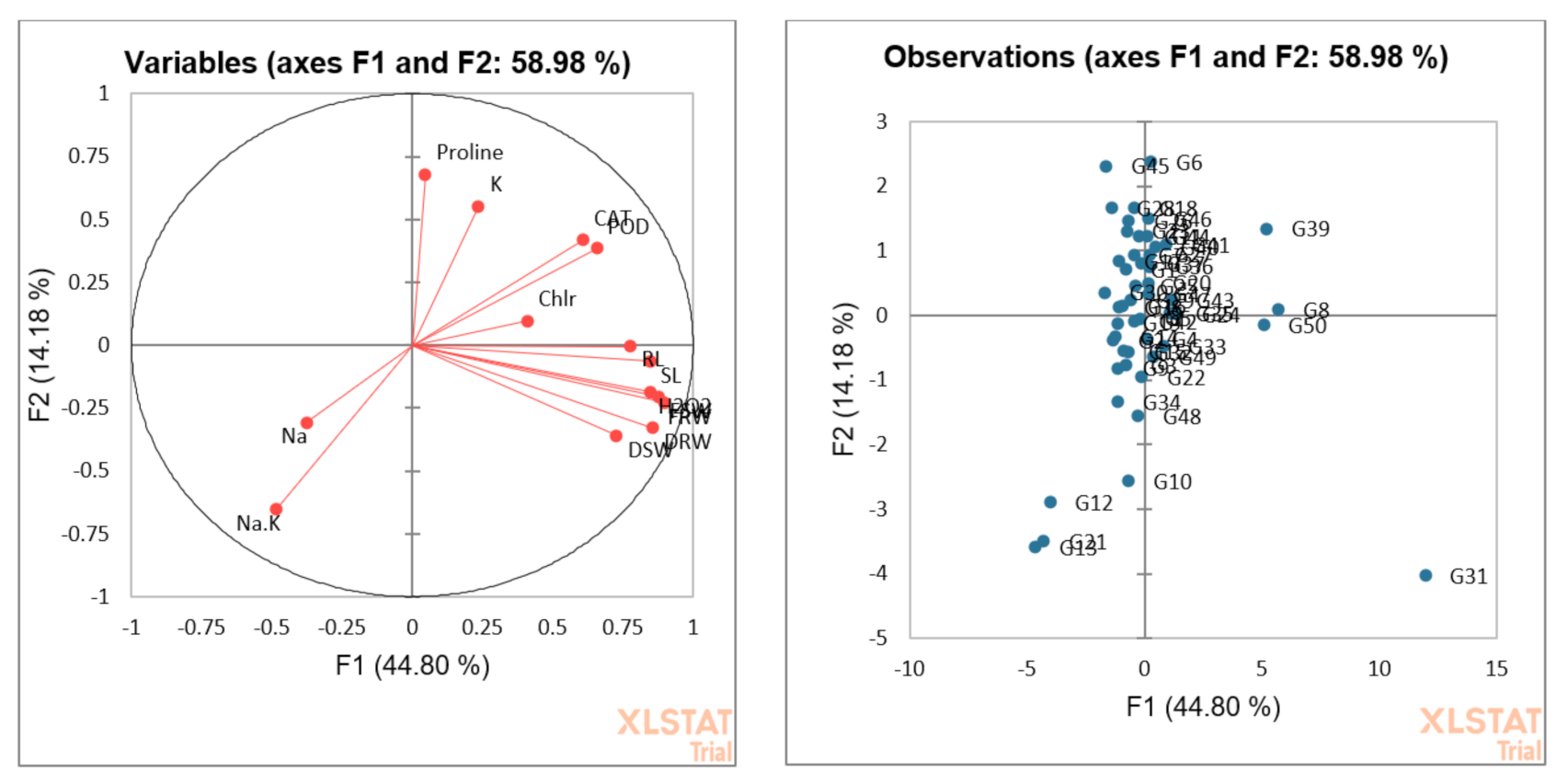
3.5. Principal Component or Biplot Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikandan, A.; Sahu, D.; Blaise, D.; Shukla, P.K. Cotton response to differential salt stress. Int. J. Agri. Sci. 2019, 11, 0975–3710. [Google Scholar]
- Ismail, A.M.; Heue, S.; Thomson, M.J.; Wissuwa, M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity stress in arid and semi-arid climates: Effects and management in field crops. J. Clim. Change Agri. 2019, 13, 197–222. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. J. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. J. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Forghani, A.H.; Almodares, A.; Ehsanpour, A.A. Potential objectives for gibberellic acid and paclobutrazol under salt stress in sweet sorghum (Sorghum bicolor [L.] Moench cv. Sofra). J. Appl. Biol. Chem. 2018, 61, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Al-Farsi, S.M.; Nawaz, A.; Nadaf, S.K.; Al-Sadi, A.M.; Siddique, K.H.; Farooq, M. Effects, tolerance mechanisms and management of salt stress in lucerne (Medicago sativa). J. Crop Pasture Sci. 2020, 71, 411–428. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, R.; Cui, Y.; Ma, Z.; Guo, Y.; Wang, Q.; Xiu, Y.; Zhang, M. Photosynthate-13C allocation in the plant-soil system after 13C–pulse labeling of Phragmites australis in different salt marshes. Geoderma 2019, 347, 252–261. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. J. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- Ding, W.; Song, L.; Wang, X.; Bi, Y. Effect of abscisic acid on heat stress tolerance in the calli from two ecotypes of Phragmites communis. J. Biol. Plant 2010, 54, 607–613. [Google Scholar] [CrossRef]
- SSharif, I.; Aleem, S.; Farooq, J.; Rizwan, M.; Younas, A.; Sarwar, G.; Chohan, S.M. Salinity stress in cotton: Effects, mechanism of tolerance and its management strategies. J. Physiol. Mol. Biol. Plants 2019, 25, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Higbie, S.M.; Wang, F.; Stewart, J.M.; Sterling, T.M.; Lindemann, W.C.; Hughs, E.; Zhang, J. Physiological response to salt (NaCl) stress in selected cultivated tetraploid cottons. Int. J. Agron. 2010, 2010, 643475. [Google Scholar] [CrossRef] [Green Version]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [Green Version]
- Majeed, S.; Malik, T.A.; Rana, I.A.; Azhar, M.T. Antioxidant and physiological responses of upland cotton accessions grown under high-temperature regimes. Iran J. Sci. Technol. Trans. A Sci. 2019, 43, 2759–2768. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Station 1950, 347, 360–361. [Google Scholar]
- Hosseini, G.; Thengane, R.J. Salinity tolerance in cotton (Gossypium hirsutum L.) genotypes. Int. J. Bot. 2007, 3, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Fielding, J.; Hall, J. A biolchemical and cytochemical study of peroxidase activity in roots of Pisum sativum: I. a comparison of DAB-peroxidase and guaiacol-peroxidase with particular emphasis on the properties of cell wall activity. J. Exp. Bot. 1978, 29, 969–981. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. J. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Torrie, J.H.; Steel, R.G. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Zhao, G.; Song, Y.; Wang, Q.; Yao, D.; Li, D.; Qin, W.; Ge, X.; Yang, Z.; Xu, W.; Su, Z.; et al. Gossypium hirsutum salt tolerance is enhanced by overexpression of G. arboreum JAZ1. Front. Bioeng. Biotechnol. 2020, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Naher, T.; Sarkar, M.; Kabir, A.; Haider, S.; Paul, N. Screening of Zn-efficient rice through hydroponic culture. J. Plant Environ. Dev. 2014, 3, 14–18. [Google Scholar]
- Sharma, N.; Acharya, S.; Kumar, K.; Singh, N.; Chaurasia, O.P. Hydroponics as an advanced technique for vegetable production: An overview. J. Soil Water Conserv. 2018, 17, 364–371. [Google Scholar] [CrossRef]
- Munawar, W.; Hameed, A.; Khan, M.K.R. Differential morphophysiological and biochemical responses of cotton genotypes under various salinity stress levels during early growth stage. Front. Plant Sci. 2021, 12, 622309. [Google Scholar] [CrossRef]
- Wang, N.; Wang, X.; Shi, J.; Liu, X.; Xu, Q.; Zhou, H.; Song, M.; Yan, G. Mepiquat chloride-priming induced salt tolerance during seed germination of cotton (Gossypium hirsutum L.) through regulating water transport and K+/Na+ homeostasis. J. Exp. Bot. 2019, 159, 168–178. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; De Pascale, S. Salt stress response in tomato beyond the salinity tolerance threshold. J. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, P.; Tiwari, R.; Meena, K.K.; Yandigeri, M.; Singh, D.P.; Arora, D.K. Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. J. Biol. Fertil. Soils. 2011, 47, 907–916. [Google Scholar] [CrossRef]
- Zhan, Y.; Wu, Q.; Chen, Y.; Tang, M.; Sun, C.; Sun, J.; Yu, C. Comparative proteomic analysis of okra (Abelmoschus esculentus L.) seedlings under salt stress. BMC Genom. 2019, 20, 381. [Google Scholar] [CrossRef]
- Farooq, M.; Rehman, A.; Al-Alawi, A.K.; Al-Busaidi, W.M.; Lee, D.-J. Integrated use of seed priming and biochar improves salt tolerance in cowpea. J. Sci. Hortic. 2020, 272, 109507. [Google Scholar] [CrossRef]
- Meneguzzo, S.; Navari-Izzo, F.; Izzo, R. NaCl effects on water relations and accumulation of mineral nutrients in shoots, roots and cell sap of wheat seedlings. J. Plant Physiol. 2000, 156, 711–716. [Google Scholar] [CrossRef]
- Akram, M.S.; Athar, H.; Ashraf, M. Improving growth and yield of sunflower (Helianthus annuus L.) by foliar application of potassium hydroxide (KOH) under salt stress. Pak.J. Bot. 2007, 39, 769–776. [Google Scholar]
- Santa-María, G.E.; Epstein, E. Potassium/sodium selectivity in wheat and the amphiploid cross wheat X Lophopyrum elongatum. J. Plant Sci. 2001, 160, 523–534. [Google Scholar] [CrossRef]
- Harinasut, P.; Tsutsui, K.; Takabe, T.; Nomura, M.; Takabe, T.; Kishitani, S. Exogenous glycinebetaine accumulation and increased salt-tolerance in rice seedlings. J. Biosci. Biotechnol. Biochem. 1996, 60, 366–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Mun, B.-G.; Khan, A.L.; Waqas, M.; Kim, H.-H.; Shahzad, R.; Imran, M.; Yun, B.-W.; Lee, I.-J. Regulation of reactive oxygen and nitrogen species by salicylic acid in rice plants under salinity stress conditions. PLoS ONE 2018, 13, e0192650. [Google Scholar] [CrossRef] [Green Version]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Chen, Y.-E.; Mao, J.-J.; Sun, L.-Q.; Huang, B.; Ding, C.-B.; Gu, Y.; Liao, J.-Q.; Hu, C.; Zhang, Z.-W.; Yuan, S.; et al. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. J. Physiol. Plant 2018, 164, 349–363. [Google Scholar] [CrossRef]
- Yassin, M.; El Sabagh, A.; Mekawy, A.M.M.; Islam, M.S.; Hossain, A.; Barutcular, C.; Alharby, H.; Bamagoos, A.; Liu, L.; Ueda, A.; et al. Comparative performance of two bread wheat (Triticum aestivum L.) genotypes under salinity stress. J. Appl. Ecol. Environ. Res. 2019, 17, 5029–5041. [Google Scholar] [CrossRef]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)—Differential response in salt-tolerant and sensitive varieties. J. Plant Sci. 2003, 165, 1411–1418. [Google Scholar] [CrossRef]
- Taghizadeh, N.; Ranjbar, G.A.; Nematzadeh, G.A.; Ramazani Moghaddam, M.R. Salt-related genes expression pattern in salt-tolerant and salt-sensitive cultivars of cotton (Gossypium sp.) Under NaCl stress. J. Plant Mo.l Breed. 2018, 6, 1–15. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Genotypes | Code | Genotypes | Code | Genotypes | Code | Genotypes | Code | Genotypes |
|---|---|---|---|---|---|---|---|---|---|
| G1 | SB-149 | G11 | IUB-13 | G21 | VH-363 | G31 | Kehkshan | G41 | KZ-189 |
| G2 | FH-452 | G12 | FH-114 | G22 | IR-3701 | G32 | Mubarak | G42 | FH-172 |
| G3 | KZ-181 | G13 | Cyto-178 | G23 | FH-170 | G33 | Bahar-2017 | G43 | BS-80 |
| G4 | KZ-191 | G14 | CRS-2007 | G24 | Cyto-124 | G34 | FH-215 | G44 | NS-121 |
| G5 | VH-341 | G15 | S-9 | G25 | AGC-2 | G35 | VH-228 | G45 | FH-118 |
| G6 | AA-703 | G16 | VH-259 | G26 | Ghouri | G36 | FH-142 | G46 | FH-169 |
| G7 | Tipo-1 | G17 | Debal | G27 | VH-339 | G37 | NIAB-777 | G47 | AGC-501 |
| G8 | MNH-992 | G18 | FH-154 | G28 | MNH-888 | G38 | VH-330 | G48 | NIAB-820 |
| G9 | Tarzan | G19 | Cyto-179 | G29 | FH-458 | G39 | CIM-595 | G49 | FH-490 |
| G10 | CRS-2 | G20 | VH-377 | G30 | AA-802 | G40 | Cyto-608 | G50 | Cyto-515 |
| Source of Variation | DF | SL | RL | FSW | FRW | DSW | DRW | Chlr | Na+ | K+ | Na+/K+ | H2O2 | Proln | POD | CAT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Salinity | 2 | 1730.49 ** | 998.7 ** | 18.25 ** | 3.86 ** | 4.29 ** | 0.18 ** | 596.36 ** | 34,853.9 ** | 4387.25 ** | 109.9 ** | 10.8 ** | 21.11 ** | 275.9 ** | 200.5 ** |
| Genotypes | 49 | 82.11 ** | 41.74 ** | 0.798 ** | 0.14 ** | 0.25 ** | 0.007 ** | 85.6 ** | 3484.6 ** | 139.97 ** | 1.55 ** | 1.9 ** | 0.20 ** | 49.11 ** | 44.91 ** |
| Salinity × Genotypes | 98 | 25.15 ** | 8.91 ** | 0.19 ** | 0.047 ** | 0.13 ** | 0.004 * | 60.41 ** | 1963.9 ** | 921.78 ** | 0.83 ** | 0.5 * | 0.20 ** | 23.43 ** | 43.72 ** |
| Error | 300 | 5.43 | 7.47 | 0.096 | 0.045 | 0.045 | 0.003 | 12.53 | 225.6 | 101.82 | 0.06 | 0.4 | 0.020 | 3.56 | 18.39 ** |
| Total | 449 | ||||||||||||||
| Normal | 150 mM | 200 mM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Traits | Genotypes | Coding | Max. Value Min. Value | Genotypes | Coding | Max. Value Min. Value | Genotypes | Coding | Max. Value Min. Value |
| SL | Cyto-515 | G50 | 30.20 | Cyto-515 | G50 | 22.33 | MNH-992 | G8 | 24.67 |
| VH-339 | G27 | 11.00 | Cyto-178 | G13 | 7.33 | Cyto-178 | G13 | 7.00 | |
| RL | Cyto-515 | G50 | 20.63 | MNH-992 | G8 | 15.67 | Cyto-515 | G50 | 17.00 |
| MNH-888 | G28 | 9.33 | VH-377 | G20 | 6.67 | VH-363 | G21 | 5.00 | |
| FSW | Cyto-515 | G50 | 2.34 | Cyto-515 | G50 | 2.18 | Cyto-515 | G50 | 1.53 |
| Ghouri | G26 | 0.48 | IUB-13 | G11 | 0.23 | Cyto-178 | G13 | 0.08 | |
| FRW | Cyto-515 | G50 | 0.93 | CIM-595 | G39 | 0.66 | Kehkshan | G31 | 0.77 |
| Mubarak | G32 | 0.12 | FH-118 | G45 | 0.05 | AA-802 | G30 | 0.02 | |
| DSW | Cyto-608 | G40 | 1.05 | Cyto-515 | G50 | 0.88 | Kehkshan | G31 | 1.19 |
| VH-339 | G27 | 0.70 | FH-114 | G12 | 0.23 | FH-114 | G12 | 0.04 | |
| DRW | VH-330 | G38 | 0.22 | AA-802 | G30 | 0.17 | Kehkshan | G31 | 0.15 |
| KZ-191 | G4 | 0.02 | KZ-191 | G4 | 0.01 | Ghouri | G26 | 0.00 | |
| Chlr | Mubarak | G32 | 47.77 | FH-452 | G2 | 40.80 | Kehkshan | G31 | 39.10 |
| FH-118 | G45 | 21.50 | Tarzan | G9 | 15.13 | FH-118 | G45 | 11.73 | |
| Na | FH-142 | G36 | 67.33 | FH-114 | G12 | 195.33 | Tarzan | G9 | 242.33 |
| CIM-595 | G39 | 15.00 | FH-172 | G42 | 38.33 | VH-377 | G20 | 45.67 | |
| K | Ghouri | G26 | 118.00 | Kehkshan | G31 | 92.33 | FH-169 | G9 | 94.67 |
| VH-228 | G35 | 42.67 | FH-172 | G42 | 20.33 | KZ-191 | G4 | 24.67 | |
| Na/K | Cyto-124 | G24 | 0.80 | VH-363 | G21 | 4.02 | FH-114 | G12 | 3.92 |
| CIM-595 | G39 | 0.35 | Kehkshan | G31 | 1.13 | FH-142 | G36 | 1.16 | |
| H2O2 | Tipo-1 | G7 | 1.40 | CIM-595 | G39 | 3.83 | Kehkshan | G31 | 4.78 |
| Bahar-2017 | G33 | 0.22 | VH-363 | G21 | 0.44 | FH-114 | G12 | 0.50 | |
| Proln | KZ-191 | G4 | 0.65 | FH-118 | G31 | 1.18 | AA-703 | G6 | 2.12 |
| CRS-2007 | G14 | 0.02 | Ghouri | G26 | 0.26 | VH-363 | G21 | 0.15 | |
| POD | AGC-501 | G47 | 15.13 | Cyto-515 | G50 | 18.67 | CIM-595 | G39 | 25.50 |
| MNH-888 | G28 | 3.70 | CRS-2 | G10 | 4.67 | Cyto-178 | G13 | 5.47 | |
| CAT | Tipo-1 | G7 | 31.80 | Kehkshan | G31 | 47.77 | CIM-595 | G39 | 49.83 |
| Bahar-2017 | G33 | 15.63 | Cyto-178 | G13 | 25.7 | Cyto-178 | G13 | 25.53 | |
| Cluster No. | No. of Genotypes | Coding |
|---|---|---|
| 1 | 20 | G1, G2, G3, G4, G12, G14, G19, G20, G21, G24, G25, G28, G29, G30, G33, G35, G37, G45, G46, G47 |
| 2 | 1 | G32 |
| 3 | 7 | G10, G13, G22, G26, G36, G38, G48 |
| 4 | 14 | G6, G8, G9, G11, G15, G16, G23, G27, G34, G40, G41, G43, G49, G50 |
| 5 | 8 | G5, G7, G17, G18, G31, G39, G42, G44 |
| Cluster No. | No. of Genotypes | Coding |
|---|---|---|
| 1 | 13 | G3, G5, G6, G9, G12, G14, G18, G28, G32, G38, G45, G47, G48 |
| 2 | 10 | G2, G8, G17, G26, G31, G35, G39, G41, G44, G49 |
| 3 | 12 | G7, G13, G20, G21, G24, G27, G29, G33, G34, G40, G43, G46 |
| 4 | 4 | G4, G10, G16, G42 |
| 5 | 11 | G1, G11, G15, G19, G22, G23, G25, G30, G36, G37, G50 |
| Cluster No. | No. of Genotypes | Coding |
|---|---|---|
| 1 | 5 | G3, G9, G12, G15, G48 |
| 2 | 5 | G1, G14, G19, G31, G50 |
| 3 | 21 | G6, G7, G8, G11, G17, G18, G23, G24, G26, G27, G28, G36, G37, G39, G40, G41, G43, G44, G45, G46, G49 |
| 4 | 2 | G4, G20 |
| 5 | 17 | G2, G5, G10, G13, G16, G21, G22, G25, G29, G30, G32, G33, G34, G35, G38, G42, G47 |
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | Cluster 5 | |
|---|---|---|---|---|---|
| SL | 19.52 | 11.33 | 18.12 | 21.88 | 21.33 |
| RL | 13.25 | 14.83 | 15.01 | 15.59 | 15.61 |
| FSW | 0.99 | 1.01 | 0.90 | 1.29 | 1.20 |
| FRW | 0.38 | 0.12 | 0.53 | 0.53 | 0.57 |
| DSW | 0.30 | 0.13 | 0.25 | 0.50 | 0.60 |
| DRW | 0.07 | 0.04 | 0.11 | 0.11 | 0.09 |
| Chlr | 31.23 | 47.77 | 31.37 | 30.88 | 30.54 |
| Na+ | 29.98 | 36.00 | 59.19 | 35.76 | 19.71 |
| K+ | 45.43 | 57.00 | 91.38 | 65.21 | 45.92 |
| Na+/K+ | 0.66 | 0.64 | 0.65 | 0.55 | 0.43 |
| H2O2 | 0.74 | 0.91 | 0.83 | 0.85 | 0.84 |
| Proln | 0.30 | 0.16 | 0.21 | 0.23 | 0.29 |
| POD | 9.80 | 6.77 | 8.50 | 8.89 | 8.55 |
| CAT | 20.98 | 17.27 | 22.93 | 21.50 | 22.53 |
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | Cluster 5 | |
|---|---|---|---|---|---|
| SL | 16.69 | 18.68 | 15.54 | 17.03 | 18.00 |
| RL | 10.64 | 12.54 | 11.41 | 11.79 | 11.32 |
| FSW | 0.62 | 0.90 | 0.68 | 0.65 | 0.80 |
| FRW | 0.18 | 0.33 | 0.19 | 0.18 | 0.25 |
| DSW | 0.13 | 0.17 | 0.12 | 0.09 | 0.15 |
| DRW | 0.04 | 0.05 | 0.04 | 0.05 | 0.06 |
| Chlr | 31.17 | 35.41 | 32.24 | 32.59 | 33.04 |
| Na+ | 148.82 | 106.73 | 80.92 | 53.33 | 102.52 |
| K+ | 67.31 | 81.87 | 34.64 | 26.08 | 53.12 |
| Na+/K+ | 2.33 | 1.32 | 2.51 | 2.15 | 2.04 |
| H2O2 | 0.78 | 1.61 | 0.92 | 1.03 | 1.02 |
| Proln | 0.62 | 0.59 | 0.50 | 0.44 | 0.56 |
| POD | 11.99 | 13.96 | 12.26 | 11.66 | 12.56 |
| CAT | 38.45 | 42.45 | 37.64 | 41.57 | 40.66 |
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | Cluster 5 | |
|---|---|---|---|---|---|
| SL | 11.55 | 16.02 | 13.79 | 13.25 | 12.52 |
| RL | 7.49 | 11.23 | 9.93 | 9.50 | 8.88 |
| FSW | 0.35 | 0.64 | 0.40 | 0.37 | 0.33 |
| FRW | 0.11 | 0.32 | 0.17 | 0.13 | 0.13 |
| DSW | 0.03 | 0.29 | 0.06 | 0.05 | 0.04 |
| DRW | 0.02 | 0.05 | 0.02 | 0.03 | 0.02 |
| Chlr | 28.36 | 33.26 | 28.21 | 29.68 | 28.42 |
| Na+ | 202.87 | 86.53 | 112.10 | 47.83 | 140.29 |
| K+ | 67.60 | 50.40 | 81.62 | 27.83 | 55.55 |
| Na+/K+ | 3.13 | 1.92 | 1.39 | 1.74 | 2.60 |
| H2O2 | 1.14 | 2.12 | 1.33 | 1.18 | 1.20 |
| Proln | 0.72 | 1.06 | 1.10 | 1.00 | 0.96 |
| POD | 16.04 | 19.45 | 18.18 | 17.33 | 16.85 |
| CAT | 43.71 | 44.31 | 43.76 | 45.42 | 41.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaheen, S.; Baber, M.; Aslam, S.; Aslam, S.; Shaheen, M.; Waheed, R.; Seo, H.; Azhar, M.T. Effect of NaCl on Morphophysiological and Biochemical Responses in Gossypium hirsutum L. Agronomy 2023, 13, 1012. https://doi.org/10.3390/agronomy13041012
Shaheen S, Baber M, Aslam S, Aslam S, Shaheen M, Waheed R, Seo H, Azhar MT. Effect of NaCl on Morphophysiological and Biochemical Responses in Gossypium hirsutum L. Agronomy. 2023; 13(4):1012. https://doi.org/10.3390/agronomy13041012
Chicago/Turabian StyleShaheen, Sabahat, Muhammad Baber, Sidra Aslam, Seema Aslam, Mehak Shaheen, Raheela Waheed, Hyojin Seo, and Muhammad Tehseen Azhar. 2023. "Effect of NaCl on Morphophysiological and Biochemical Responses in Gossypium hirsutum L." Agronomy 13, no. 4: 1012. https://doi.org/10.3390/agronomy13041012