

The Changes of Tolerance, Accumulation and Oxidative Stress Response to Cadmium in Tobacco Caused by Introducing Datura stramonium L. Genes
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Hydroponic Experiment
2.3. Determination of Cd Concentration
2.4. ROS and MDA Assay
2.5. Chlorophyll and Proline Assay
2.6. Antioxidant Enzymes Assays
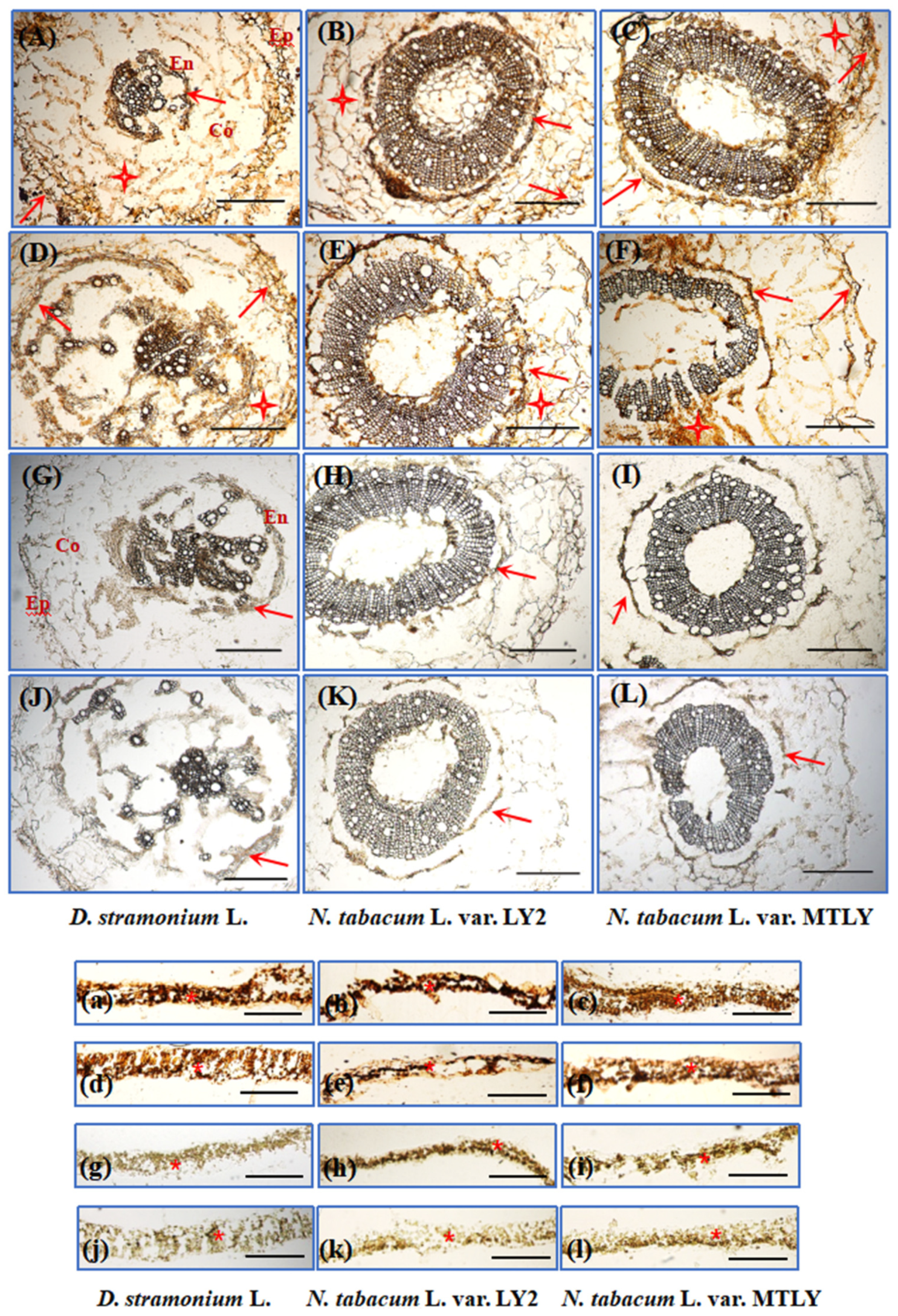
2.7. Histochemical Analysis
2.8. Statistical Analysis
3. Results
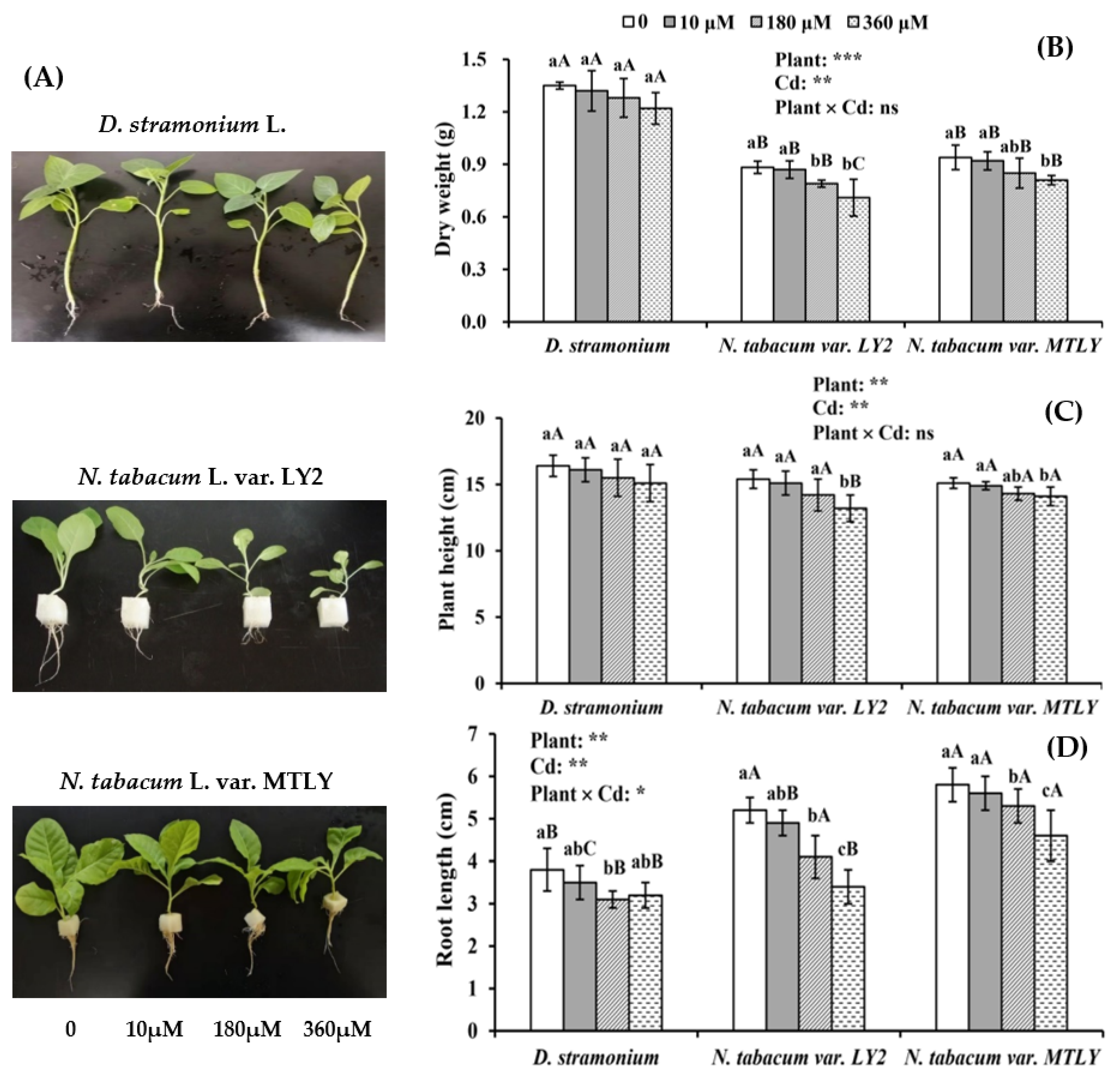
3.1. Effects of Cadmium Stress on Seedlings Growth
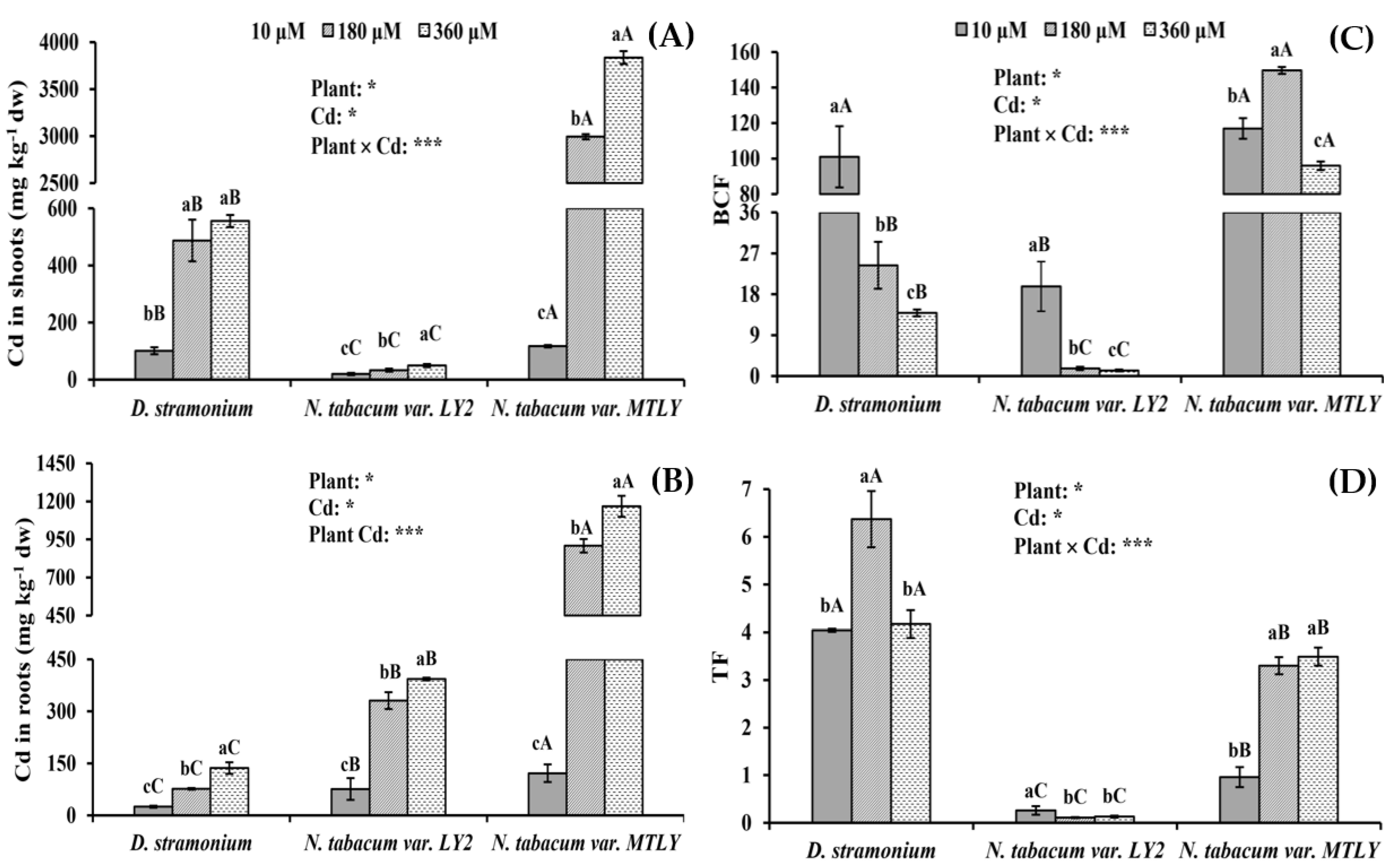
3.2. Cd Accumulation in Seedling Tissues
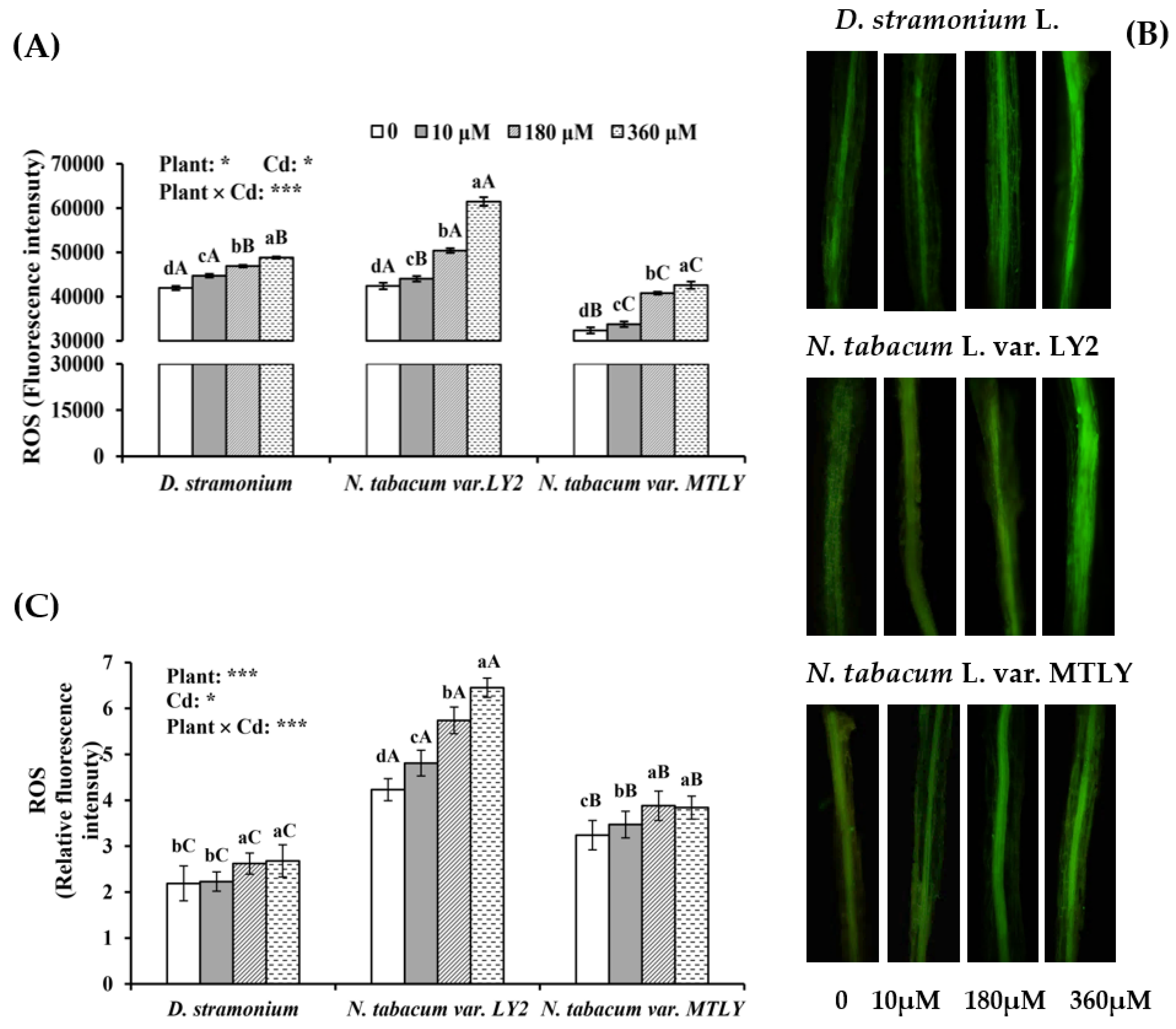
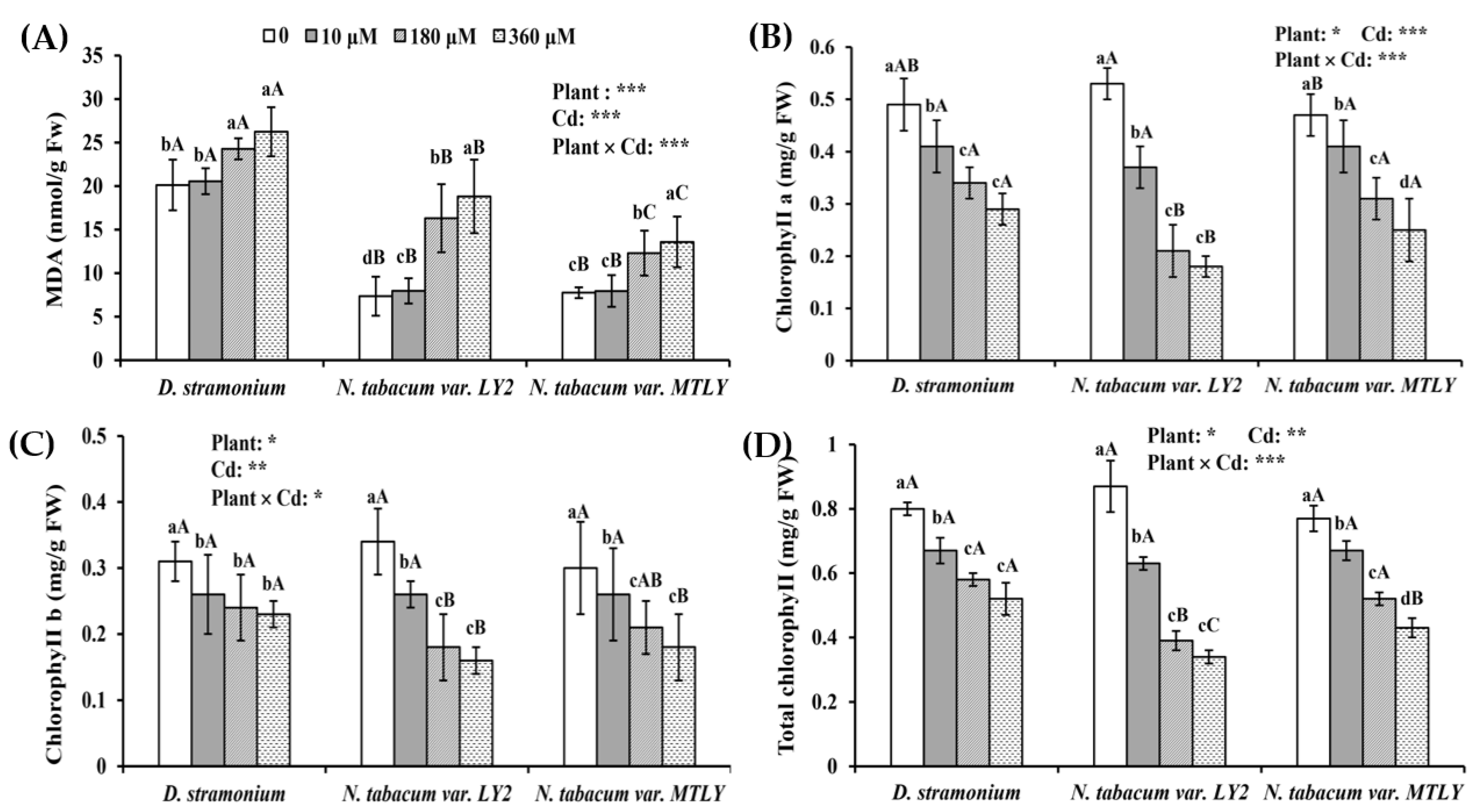
3.3. Cd Exposure Induced Oxidative Stress in Seedlings
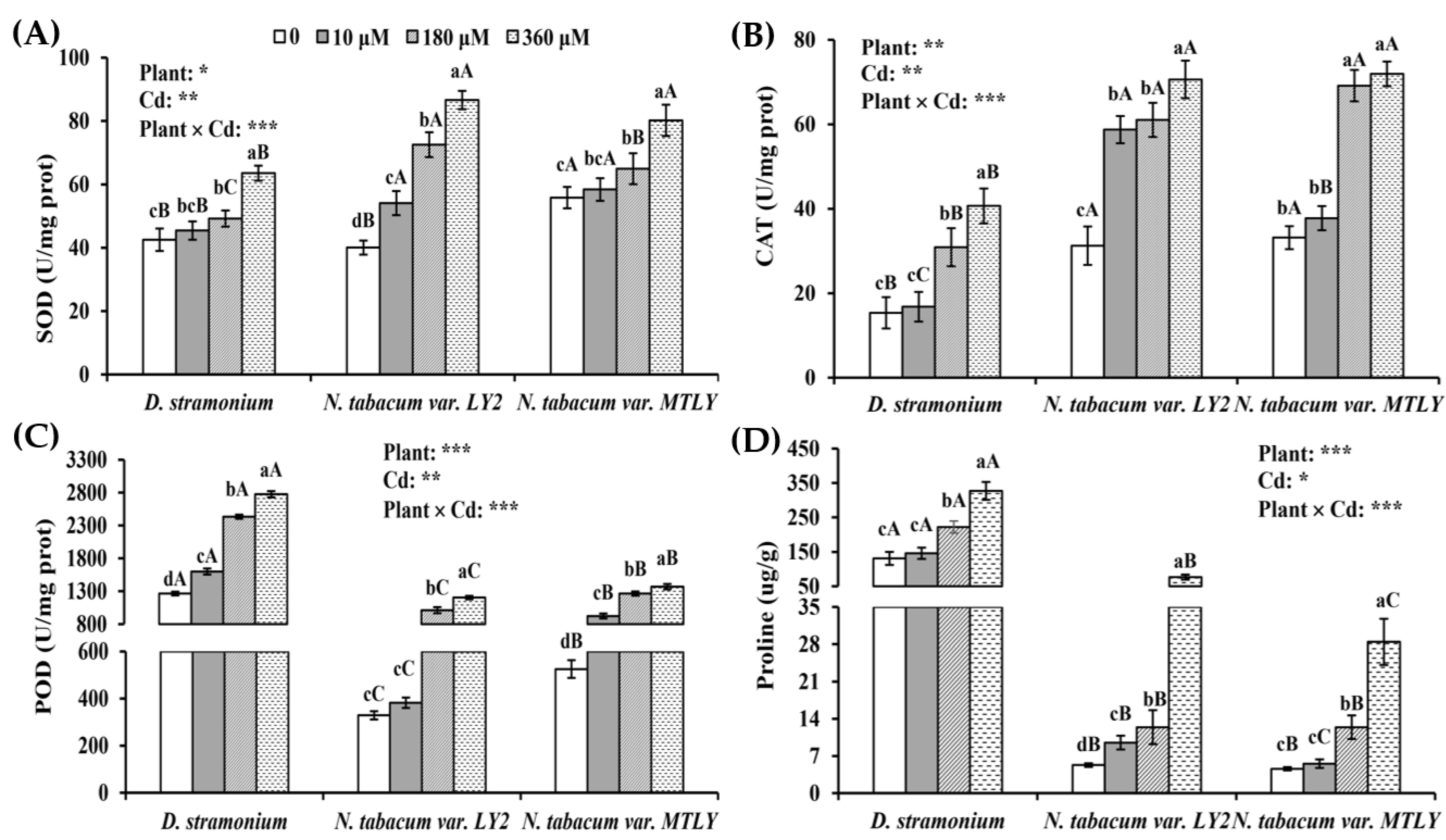
3.4. Antioxidant Response of Seedlings to Cd Stress
3.5. Cd Exposure Changed Secondary Metabolism of Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pouresmaieli, M.; Ataei, M.; Forouzandeh, P.; Azizollahi, P.; Mahmoudifard, M. Recent progress on sustainable phytoremediation of heavy metals from soil. J. Environ. Chem. Eng. 2022, 10, 108482. [Google Scholar] [CrossRef]
- Jia, W.T.; Lv, S.L.; Feng, J.J.; Li, J.H.; Li, Y.X.; Li, S.Z. Morphophysiological characteristic analysis demonstrated the potential of sweet sorghum (Sorghum bicolor (L.) Moench) in the phytoremediation of cadmium-contaminated soils. Environ. Sci. Pollut. Res. 2016, 23, 18823–18831. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Carmona, F.F.; Bhargava, M.; Srivastava, S. Approaches for enhanced phytoextraction of heavy metals. J. Environ. Manag. 2012, 105, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Elango, D.; Devi, K.D.; Jeyabalakrishnan, H.K.; Rajendran, K.; Haridass, V.K.T.; Dharmaraj, D.; Charuchandran, C.V.; Wang, W.Y.; Fakude, M.; Mishra, R.; et al. Agronomic, breeding, and biotechnological interventions to mitigate heavy metal toxicity problems in agriculture. J. Agric. Food Res. 2022, 10, 100374. [Google Scholar] [CrossRef]
- Guo, B.; Liang, Y.C.; Zhu, Y.G.; Zhao, F.J. Role of salicylic acid in alleviating oxidative damage in rice roots (Oryza sativa) subjected to cadmium stress. Environ. Pollut. 2007, 147, 743–749. [Google Scholar] [CrossRef]
- Jean, L.; Bordas, F.; Gautier-Moussard, C.; Vernay, P.; Hitmi, A.; Bollinger, J.C. Effect of citric acid and EDTA on chromium and nickel uptake and translocation by Datura innoxia. Environ. Pollut. 2008, 153, 555–563. [Google Scholar] [CrossRef]
- Yang, X.E.; Feng, Y.; He, Z.L.; Stoffella, P.J. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 339–353. [Google Scholar] [CrossRef]
- Seth, C.S. A review on mechanisms of plant tolerance and role of transgenic plants in environmental clean-up. Bot. Rev. 2012, 78, 32–62. [Google Scholar] [CrossRef]
- Liang, H.M.; Lin, T.H.; Chiou, J.M.; Yeh, K.C. Model evaluation of the phytoextraction potential of heavy metal hyperaccumulators and non-hyperaccumulators. Environ. Pollut. 2009, 157, 1945–1952. [Google Scholar] [CrossRef]
- Kumar, G.; Kushwaha, H.R.; Panjabi-Sabharwal, V.; Kumari, S.; Joshi, R.; Karan, R.; Mittal, S.; Pareek, S.L.S.; Pareek, A. Clustered metallothionein genes are co-regulated in rice and ectopic expression of OsMT1e-P confers multiple abiotic stress tolerance in tobacco via ROS scavenging. BMC Plant Bio. 2012, 12, 107. [Google Scholar]
- Kozak, K.; Antosiewicz, D.M. Tobacco as an efficient metal accumulator. BioMetals 2022. [Google Scholar] [CrossRef]
- Zhi, J.K.; Liu, X.; Yin, P.; Yang, R.X.; Liu, J.F.; Xu, J.C. Overexpression of the metallothionein gene PaMT3-1 from Phytolacca americana enhances plant tolerance to cadmium. Plant Cell Tissue Organ Cult. 2020, 143, 211–218. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Anand, U.; Kumar, R.; Ghorai, M.; Aftab, T.; Jha, N.K.; Rajapaksha, A.U.; Bundschuh, J.; Bontempi, E.; Dey, A. Phytoremediation and sequestration of soil metals using the CRISPR/Cas9 technology to modify plants: A review. Environ. Chem. Lett. 2022, 21, 429–445. [Google Scholar] [CrossRef]
- Pilon-Smits, E.; Pilon, M. Phytoremediation of metals using transgenic plants. Crit. Rev. Plant Sci. 2002, 21, 439–456. [Google Scholar] [CrossRef]
- Thu-Hang, P.; Bassie, L.; Safwat, G.; Trung-Nghia, P.; Christou, P.; Capell, T. Expression of a heterologous S-adenosylmethionine decarboxylase cDNA in plants demonstrates that changes in S-adenosyl-L-methionine decarboxylase activity determine levels of the higher polyamines spermidine and spermine. Plant Physiol. 2002, 129, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkorta, I.; Hernandez-Allica, J.; Becerril, J.M.; Amezaga, I.; Albizu, I.; Garbisu, C. Recent findings on the phytoremediation of soils contaminated with environmentally toxic heavy metals and metalloids such as zinc, cadmium, lead, and arsenic. Rev. Environ. Sci. Biotechnol. 2004, 3, 71–90. [Google Scholar] [CrossRef]
- Wei, Z.Z.; Wei, K.Q.; Yang, J.W. Distant Hybridization Breeding of Tobacco, 2nd ed.; China Agricultural Science and Technology Press: Beijing, China, 2019. [Google Scholar]
- Wei, K.Q.; Yang, J.X.; Wei, Z.Z. Pollination following grafting introduces efficiently Ocimum basilicum L. genes into Nicotiana tabacum L. Span. J. Agric. Res. 2013, 11, 1068–1077. [Google Scholar] [CrossRef] [Green Version]
- Wei, K.Q.; Pang, S.X.; Yang, J.X.; Wei, Z.Z. Enhancement of cadmium tolerance and accumulation by introducing Perilla frutescens (L.) Britt var. frutescens genes in Nicotiana tabacum L. plants. Environ. Sci. Pollut. Res. 2015, 22, 5405–5416. [Google Scholar] [CrossRef]
- Wei, Z.Z.; Yan, X.P.; Deng, Z.F.; Yang, J.X.; Wang, Y.T. Research on distant hybridization between tobacco and D. stramonium. Chin. Tob. Sci. 2005, 1, 1–5. [Google Scholar]
- Wei, K.Q.; Son, X.; Yang, J.X.; Wei, Z.Z. Chemical components in the intergeneric hybrid between tobacco and medicinal plant Datura metel L. Nat. Prod. Res. Dev. 2008, 20, 778–781. [Google Scholar]
- Vaillant, N.; Monnet, F.; Hitmi, A.; Sallanon, H.; Coudret, A. Comparative study of responses in four Datura species to a zinc stress. Chemosphere 2005, 59, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Shirkhani, Z.; Chehregani Rad, A.; Mohsenzadeh, F. Effects of soil and foliar cadmium application on morphological, physiological, genetic and epigenetic characteristics of Datura Stramonium L. Int. J. Environ. Sci. Technol. 2022, 19, 9067–9082. [Google Scholar] [CrossRef]
- Zhou, Z.; Yang, S.Q.; Liu, H.T.; Chen, X.F.; Chen, X.; Zhang, B. Uptake and subcellular distribution of cadmium in Datura stramonium L. J. Agric. Resour. Environ. 2019, 36, 385–391. [Google Scholar]
- Boojar, M.M.A.; Goodarzi, F. The copper tolerance strategies and the role of antioxidative enzymes in three plant species grown on copper mine. Chemosphere 2007, 67, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Banković-Ilić, I.B.; Stamenković, O.S.; Veljković, V.B. Biodiesel production from non-edible plant oils. Renew. Sustain. Energ. Rev. 2012, 16, 3621–3647. [Google Scholar] [CrossRef]
- Varun, M.; D’Souza, R.; Pratas, J.; Paul, M.S. Metal contamination of soils and plants associated with the glass industry in North Central India: Prospects of phytoremediation. Environ. Sci. Pollut. Res. 2012, 19, 269–281. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, W.W.; Hanna, M.A.; Zhang, Y.P.; Bhadury, P.S.; Wang, Y.; Song, B.A.; Yang, S. Biodiesel preparation, optimization, and fuel properties from non-edible feedstock. Datura stramonium L. Fuel 2012, 91, 182–186. [Google Scholar] [CrossRef]
- Shirkhani, Z.; Rad, A.C.; Mohsenzadeh, F. Improving Cd-phytoremediation ability of Datura stramonium L. by chitosan and chitosan nanoparticles. Biologia 2021, 76, 2161–2171. [Google Scholar] [CrossRef]
- Liu, W.X.; Shang, S.H.; Xue, F.; Zhang, G.P.; Wu, F.B. Modulation of exogenous selenium in cadmium-induced changes in antioxidative metabolism, cadmium uptake, and photosynthetic performance in the 2 tobacco genotypes differing in cadmium tolerance. Environ. Toxicol. Chem. 2015, 34, 92–99. [Google Scholar] [CrossRef]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ. 2008, 54, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, Y.X.; Huang, Z.B.; Huang, L. Antioxidative response of metal-accumulator and non-accumulator plants under cadmium stress. Plant Soil. 2008, 310, 137–149. [Google Scholar] [CrossRef]
- Kavuliová, J.; Kaduková, J.; Ivánová, D. The evaluation of heavy metal toxicity in plants using the biochemical tests. Nova Biotechnol. Chim. 2012, 11, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Cadmium induces different biochemical responses in wild type and catalase-deficient tobacco plants. Environ. Exp. Bot. 2015, 109, 201–211. [Google Scholar] [CrossRef]
- Muradoglu, F.; Gundogdu, M.; Ercisli, S.; Encu, T.; Balta, F.; Jaafar, H.Z.E.; Zia-Ul-Haq, M. Cadmium toxicity affects chlorophyll a and b content, antioxidant enzyme activities and mineral nutrient accumulation in strawberry. Biol. Res. 2015, 48, 11. [Google Scholar] [CrossRef] [Green Version]
- Eapen, S.; D’Souza, S.F. Prospects of genetic engineering of plants for phytoremediation of toxic metals. Biotechnol. Adv. 2005, 23, 97–114. [Google Scholar] [CrossRef]
- Sarma, H. Metal hyperaccumulation in plants: A review focusing on phytoremediation technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.G.; Zhu, Q.S.; Zhang, Z.J.; Xu, J.K.; Yang, J.C.; Wong, M.H. Variations in cadmium accumulation among rice cultivars and types and the selection of cultivars for reducing cadmium in the diet. J. Sci. Food. Agric. 2005, 85, 147–153. [Google Scholar] [CrossRef]
- Liu, Y.G.; Wang, X.; Zeng, G.M.; Qu, D.; Gu, J.J.; Zhou, M.; Chai, L.Y. Cadmium-induced oxidative stress and response of the ascorbate–glutathione cycle in Bechmeria nivea (L.) Gaud. Chemosphere 2007, 69, 99–107. [Google Scholar] [CrossRef]
- Grant, C.A.; Clarke, J.M.; Duguid, S.; Chaney, R.L. Selection and breeding of plant cultivars to minimize cadmium accumulation. Sci. Total Environ. 2008, 390, 301–310. [Google Scholar] [CrossRef]
- Jin, X.F.; Yang, X.E.; Mahmood, Q.; Islam, E.; Liu, D.; Li, H. Response of antioxidant enzymes, ascorbate and glutathione metabolism towards cadmium in hyperaccumulator and nonhyperaccumulator ecotypes of Sedum alfredii H. Environ. Toxicol. 2008, 23, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Su, L.Y.; Yang, J.Z.; Yuan, J.G.; Yin, A.G.; Qiu, Q.; Zhang, K.; Yang, Z.Y. Comparisons of cadmium subcellular distribution and chemical forms between low-Cd and high-Cd accumulation genotypes of watercress (Nasturtium officinale L. R. Br.). Plant Soil. 2015, 396, 325–337. [Google Scholar] [CrossRef]
- Deng, X.P.; Yan, X.; Wei, H.; Zhang, H.X.; Shen, Z.G. Cadmium-induced oxidative damage and protective effects of N-acetyl-L-cysteine against cadmium toxicity in Solanum nigrum L. J. Hazard Mater. 2010, 180, 722–729. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, P.; da Silva, J.A.T.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 872875. [Google Scholar] [CrossRef] [Green Version]
- Emamverdian, A.; Ding, Y.L.; Mokhberdoran, F.; Xie, Y.F. Heavy Metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelova, Z.; Georgiev, S.; Roos, W. Elicitation of plants. Biotechnol. Biotechnol. Equip. 2006, 20, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Hoque, M.A.; Okuma, E.; Banu, M.N.A.; Shimoishi, Y.; Nakamur, Y.; Murata, Y. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef]
- Shukla, D.; Huda, K.M.K.; Banu, M.S.A.; Gill, S.S.; Tuteja, R.; Tuteja, N. OsACA6, a P-type 2B Ca2+ ATPase functions in cadmium stress tolerance in tobacco by reducing the oxidative stress load. Planta 2014, 240, 809–824. [Google Scholar] [CrossRef]
- da Silva Magedans, Y.V.; Matsuura, H.N.; Tasca, R.A.J.C.; Wairich, A.; de Oliveira Junkes, C.F.; de Costa, F.; Fett-Neto, A.G. Accumulation of the antioxidant alkaloid brachycerine from Psychotria brachyceras Müll. Arg. is increased by heat and contributes to oxidative stress mitigation. Environ Exp Bot. 2017, 143, 185–193. [Google Scholar] [CrossRef]
- Lu, M.; Yu, S.; Lian, J.P.; Wang, Q.; He, Z.L.; Feng, Y.; Yang, X.E. Physiological and metabolomics responses of two wheat (Triticum aestivum L.) genotypes differing in grain cadmium accumulation. Sci. Total Environ. 2021, 769, 145345. [Google Scholar] [CrossRef]
- Djemal, R.; Khoudi, H. The ethylene-responsive transcription factor of durum wheat, TdSHN1, confers cadmium, copper, and zinc tolerance to yeast and transgenic tobacco plants. Protoplasma 2022, 259, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Berkova, V.; Berka, M.; Griga, M.; Kopecka, R.; Prokopova, M.; Luklova, M.; Horacek, J.; Smykalova, I.; Cicmanec, P.; Novak, J.; et al. Molecular Mechanisms underlying Flax (Linum usitatissimum L.) tolerance to cadmium: A case study of proteome and metabolome of four different Flax genotypes. Plants 2022, 11, 2931. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Zhang, D.Z.; Luo, Y.; Zhang, Y.B.; Xu, L.; Chen, P.L.; Wu, E.G.; Ma, Q.; Wang, H.L.; Zhao, L.; et al. Cadmium tolerance and accumulation from the perspective of metal ion absorption and root exudates in broomcorn millet. Ecotoxicol. Environ. Saf. 2023, 250, 114506. [Google Scholar] [CrossRef]
- Xie, L.P.; He, X.Y.; Shang, S.H.; Zheng, W.T.; Liu, W.X.; Zhang, G.P.; Wu, F.B. Comparative proteomic analysis of two tobacco (Nicotiana tabacum) genotypes differing in Cd tolerance. Biometals 2014, 27, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lu, X.Y.; Guo, X.R.; Pan, Y.J.; Yu, B.F.; Tang, Z.H.; Guo, Q.X. Differential responses to Cd stress induced by exogenous application of Cu, Zn or Ca in the medicinal plant Catharanthus roseus. Ecotoxicol. Environ. Saf. 2018, 157, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Touati, M.; Bottega, S.; Castiglione, M.R.; Sorce, C.; Béjaoui, Z.; Spanò, C. Modulation of the defence responses against Cd in willow species through a multifaceted analysis. Plant Physiol. Biochem. 2019, 142, 125–136. [Google Scholar] [CrossRef]
- Khator, K.; Saxena, I.; Shekhawat, G.S. Nitric oxide induced Cd tolerance and phytoremediation potential of B. juncea by the modulation of antioxidant defense system and ROS detoxification. Biometals 2021, 34, 15–32. [Google Scholar] [CrossRef]
- Joo, J.H.; Wang, S.Y.; Chen, J.G.; Jones, A.M.; Fedoroff, N.V. Different signaling and cell death roles of heterotrimeric G protein α and β subunits in the Arabidopsis oxidative stress response to ozone. Plant Cell 2005, 17, 957–970. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Liang, Y.P.; Wu, Z.N.; Zhang, L.; Liu, Z.W.; Li, Q.F.; Chen, X.J.; Guo, W.L.; Jiang, L.N.; Pan, F.F.; et al. Effects of tetracycline on growth, oxidative stress response, and metabolite pattern of ryegrass. J. Hazard. Mater. 2019, 380, 120885. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Reprint of: Photoperoxidation in isolated chloroplast I.: Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 2022, 726, 109248. [Google Scholar] [CrossRef]
- Sun, R.L.; Zhou, Q.L.; Sun, F.H.; Jin, C.X. Antioxidative defense and proline/phytochelatin accumulation in a newly discovered Cd-hyperaccumulator, Solanum nigrum L. Environ. Exp. Bot. 2007, 60, 468–476. [Google Scholar] [CrossRef]
- Zhang, D.Z.; Wang, P.H.; Zhao, H.X. Determination of the content of free proline in wheat leaves. Plant Physiol. Commun. 1990, 4, 62–65. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Argyropoulou, C.; Akoumianaki-Ioannidou, A.; Christodoulakis, N.S.; Fasseas, C. Leaf anatomy and histochemistry of Lippia citriodora (Verbenaceae). Aust. J. Bot. 2010, 58, 398–409. [Google Scholar] [CrossRef]
- Janošević, D.; Budimir, S.; Alimpić, A.; Marin, P.; Sheef, M.A.; Giweli, A.; Duletić-Laušević, S. Micromorphology and histochemistry of leaf trichomes of Salvia aegyptiaca (Lamiaceae). Arch Biol. Sci. 2016, 68, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Tozin, L.R.S.; Rodrigues, T.M. Morphology and histochemistry of glandular trichomes in Hyptis villosa Pohl ex Benth. (Lamiaceae) and differential labeling of cytoskeletal elements. Acta Bot. Bras. 2017, 31, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Gomez, A.A.; Mercado, M.I.; Belizán, M.M.E.; Ponessa, G.; Vattuone, M.A.; Sampietro, D.A. In situ histochemical localization of alkaloids in leaves and pods of Prosopis ruscifolia. Flora 2019, 256, 1–6. [Google Scholar] [CrossRef]
- Gorinova, N.; Nedkovska, M.; Todorovska, E.; Simova-Stoilova, L.; Stoyanova, Z.; Georgieva, K.; Demirevska-Kepova, K.; Atanassov, A.; Herzig, R. Improved phytoaccumulation of cadmium by genetically modified tobacco plants (Nicotiana tabacum L.). Physiological and biochemical response of the transformants to cadmium toxicity. Environ. Pollut. 2007, 145, 161–170. [Google Scholar] [CrossRef]
- Golia, E.E.; Dimirkou, A.; Mitsios, I.K. Accumulation of metals on tobacco leaves (Primings) grown in an agricultural area in relation to soil. Bull. Environ. Contam. Toxicol. 2007, 79, 158–162. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Manquián-Cerda, K.; Cruces, E.; Escudey, M.; Zúñiga, G.; Calderónd, R. Effect of cadmium on phenolic compounds, antioxidant enzyme activity and oxidative stress in blueberry (Vaccinium corymbosum L.) plantlets grown in vitro. Ecotoxicol. Environ. Safe. 2016, 133, 316–326. [Google Scholar] [CrossRef]
- Soukupová, J.; Cvikrová, M.; Albrechtová, J.; Rock, B.N.; Eder, J. Histochemical and biochemical approaches to the study of phenolic compounds and peroxidases in needles of Norway spruce (Picea abies). New Phytol. 2000, 146, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Mesjasz-Przybylowicz, J.; Barnabas, A.; Przybyiowicz, W. Comparison of cytology and distribution of nickel in roots of Ni-hyperaccumulating and non-hyperaccumulating genotypes of Senecio coronatus. Plant Soil 2007, 293, 61–78. [Google Scholar] [CrossRef]
- Zheng, Z.G.; Wu, M. Cadmium treatment enhances the production of alkaloid secondary metabolites in Catharanthus roseus. Plant Sci. 2004, 166, 507–514. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissues | Secondary Metabolites | Cd Treatments | D. stramonium | N. tabacum L. var. LY2 | N. tabacum L. var. MTLY |
|---|---|---|---|---|---|
| Root | Phenolic compounds | 0 | +++ | ++ | +++ |
| 10 µM | +++ | + | +++ | ||
| 180 µM | ++ | + | +++ | ||
| 360 µM | ++ | + | ++ | ||
| Alkaloids | 0 | +++ | ++ | +++ | |
| 10 µM | ++ | +++ | ++ | ||
| 180 µM | ++ | ++ | +++ | ||
| 360 µM | +++ | + | ++++ | ||
| Leaf | Phenolic compounds | 0 | +++ | ++ | +++ |
| 10 µM | ++++ | ++ | ++++ | ||
| 180 µM | +++ | +++ | ++++ | ||
| 360 µM | +++ | + | +++ | ||
| Alkaloids | 0 | +++ | ++ | +++ | |
| 10 µM | ++++ | ++ | +++ | ||
| 180 µM | +++ | + | +++ | ||
| 360 µM | +++ | + | +++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, K.; Guo, T. The Changes of Tolerance, Accumulation and Oxidative Stress Response to Cadmium in Tobacco Caused by Introducing Datura stramonium L. Genes. Agronomy 2023, 13, 882. https://doi.org/10.3390/agronomy13030882
Wei K, Guo T. The Changes of Tolerance, Accumulation and Oxidative Stress Response to Cadmium in Tobacco Caused by Introducing Datura stramonium L. Genes. Agronomy. 2023; 13(3):882. https://doi.org/10.3390/agronomy13030882
Chicago/Turabian StyleWei, Keqiang, and Tingting Guo. 2023. "The Changes of Tolerance, Accumulation and Oxidative Stress Response to Cadmium in Tobacco Caused by Introducing Datura stramonium L. Genes" Agronomy 13, no. 3: 882. https://doi.org/10.3390/agronomy13030882