

Genetic Diversity of Global Faba Bean Germplasm Resources Based on the 130K TNGS Genotyping Platform
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Extraction and SNP Genotyping
2.3. Data Analysis
3. Results
3.1. The Genetic Diversity Indices of SNP Markers
3.2. Population Structure Analysis
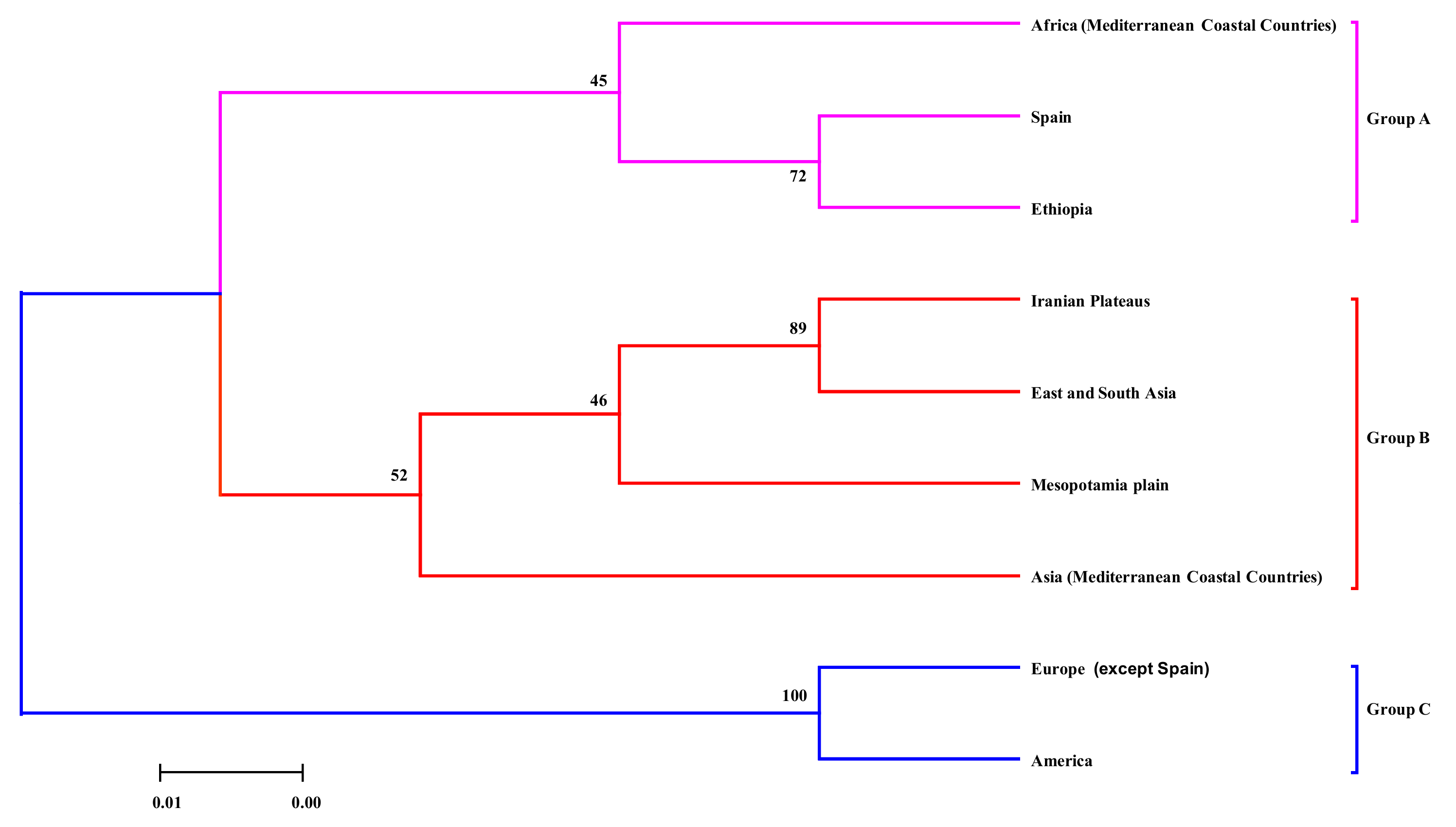
3.3. Clustering Analysis
3.4. Genetic Diversity Analysis of Global Faba Bean Germplasm Resources
3.5. Analysis of Molecular Variance
4. Discussion
4.1. On the Faba Bean SNP Genotyping Platform
4.2. On Genetic Diversity Studies of the Faba Bean Genetic Resources
4.3. On the Origin and Distribution of Faba Bean
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierre, J.; Suso, M.J.; Moreno, M.T.; Eanault, R.; Le Guen, J. Diversity and efficiency of the pollinating entomofauna (Hymenoptera: Apidae) of faba bean (Vicia faba L.) in two locations in France and Spain. Ann. De La Société Entomol. De Fr. 1999, 35, 312–318. [Google Scholar]
- Suso, M.J.; Pierre, J.; Moreno, M.T.; Eanault, R.; Le Guen, J. Variation in outcrossing levels in faba bean cultivars: Role of ecological factors. J. Agric. Sci. 2001, 136, 399–405. [Google Scholar] [CrossRef]
- FAOSTAT. Data from: FAOSTAT Databases, Food and Agriculture Organization. 2020. Available online: https://www.fao.org/faostat/en/#data/TCL (accessed on 3 November 2022).
- Ma, Y.; Bao, S.Y.; Yang, T.; Hu, J.G.; Guan, J.P.; He, Y.H.; Wang, X.J.; Wan, Y.L.; Sun, X.L.; Jiang, J.Y.; et al. Genetic linkage map of Chinese native variety faba bean (Vicia faba L.) based on simple sequence repeat markers. Plant Breed. 2013, 132, 397–400. [Google Scholar] [CrossRef]
- Duc, G. Faba bean (Vicia faba L.). Field Crop. Res. 1997, 53, 99–109. [Google Scholar] [CrossRef]
- Yin, J.; Sui, Z.M.; Li, Y.; Yang, H.J.; Yuan, L.; Huang, J.G. A new function of white-rot fungi Ceriporia lacerata HG2011: Improvement of biological nitrogen fixation of broad bean (Vicia faba). Microbiol. Res. 2022, 256, 126939. [Google Scholar] [CrossRef]
- Ingraffia, R.; Amato, G.; Frenda, A.S.; Giambalvo, D. Impacts of arbuscular mycorrhizal fungi on nutrient uptake, N2 fixation, N transfer, and growth in a wheat/faba bean intercropping system. PLoS ONE 2019, 14, e0213672. [Google Scholar] [CrossRef]
- Kaur, S.; Cogan, N.; Forster, J.; Paull, J. Assessment of genetic diversity in faba bean based on single nucleotide polymorphism. Diversity 2014, 6, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Mulugeta, B.; Tesfaye, K.; Keneni, G.; Ahmed, S. Genetic diversity in spring faba bean (Vicia faba L.) genotypes as revealed by high-throughput KASP SNP markers. Genet. Resour. Crop Evol. 2021, 68, 1971–1986. [Google Scholar] [CrossRef]
- Yang, T.; Liu, R.; Luo, Y.F.; Hu, S.N.; Wang, D.; Wang, C.Y.; Pandey, M.K.; Ge, S.; Xu, Q.L.; Li, N.N.; et al. Improved pea reference genome and pan-genome highlight genomic features and evolutionary characteristics. Nat. Genet. 2022, 54, 1553–1563. [Google Scholar] [CrossRef]
- Guan, J.T.; Zhang, J.T.; Gong, D.; Zhang, Z.Q.; Yu, Y.; Luo, G.L.; Somta, P.; Hu, Z.; Wang, S.H.; Yuan, X.X.; et al. Genomic analyses of rice bean landraces reveal adaptation and yield related loci to accelerate breeding. Nat. Commun. 2022, 13, 5707–5722. [Google Scholar] [CrossRef]
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J.; et al. Genome sequence of mungbean and insights into evolution within Vigna species. Nat. Commun. 2014, 5, 5443–5451. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Wang, Y.; Peng, J.X.; Fan, B.J.; Xu, D.X.; Wu, J.; Cao, Z.M.; Gao, Y.Q.; Wang, X.Q.; Li, S.T.; et al. High-quality genome assembly and pan-genome studies facilitate genetic discovery in mung bean and its improvement. Plant Comm. 2022, 3, 100352–100367. [Google Scholar] [CrossRef]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.M.; Rosen, B.D.; Tar’an, B.; et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Chen, W.B.; Li, Y.P.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.A.; Azam, S.; Fan, G.Y.; Whaley, A.M.; et al. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2011, 30, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Garg, G.; Kamphuis, L.G.; Bayer, P.E.; Kaur, P.; Dudchenko, O.; Taylor, C.M.; Frick, K.M.; Foley, R.C.; Gao, L.L.; Aiden, E.L.; et al. A pan-genome and chromosome-length reference genome of narrow-leafed lupin (Lupinus angustifolius) reveals genomic diversity and insights into key industry and biological traits. Plant J. 2022, 111, 1252–1266. [Google Scholar] [CrossRef]
- Johnston, J.S.; Bennett, M.D.; Rayburn, A.L.; Galbraith, D.W.; Price, H.J. Reference standards for determination of DNA content of plant nuclei. Am. J. Bot. 1999, 86, 609–613. [Google Scholar] [CrossRef]
- Hill, C.B.; Wong, D.; Tibbits, J.; Forrest, K.; Hayden, M.; Zhang, X.Q.; Westcott, S.; Angessa, T.T.; Li, C.D. Targeted enrichment by solution-based hybrid capture to identify genetic sequence variants in barley. Sci. Data 2019, 6, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Terracciano, I.; Cantarella, C.; Fasano, C.; Cardi, T.; Mennella, G.; D’Agostino, N. Liquid-phase sequence capture and targeted re-sequencing revealed novel polymorphisms in tomato genes belonging to the MEP carotenoid pathway. Sci. Rep. 2017, 7, 5616–5628. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.F.; Wang, H.W.; Tao, J.J.; Ren, Y.H.; Xu, C.; Wu, K.S.; Zou, C.; Zhang, J.N.; Xu, Y.B. Development of multiple SNP marker panels affordable to breeders through genotyping by target sequencing (GBTS) in maize. Mol. Breed. 2019, 39, 37–48. [Google Scholar] [CrossRef]
- Hussain, I.; Ali, S.; Liu, W.G.; Awais, M.; Li, J.H.; Liao, Y.L.; Zhu, M.S.; Fu, C.Y.; Liu, D.L.; Wang, F. Identification of heterotic groups and patterns based on genotypic and phenotypic characteristics among rice accessions of diverse origins. Front. Genet. 2022, 13, 811124. [Google Scholar] [CrossRef]
- Li, X.W.; Zheng, H.L.; Wu, W.S.; Liu, H.L.; Wang, J.G.; Jia, Y.; Li, J.M.; Yang, L.M.; Lei, L.; Zou, D.T.; et al. QTL mapping and candidate gene analysis for alkali tolerance in Japonica rice at the bud stage based on linkage mapping and genome-wide association study. Rice 2020, 13, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhang, Y.B.; Ren, H.; Li, X.; Zhang, X.; Zhang, Z.Y.; Zhang, C.L.; Liu, S.J.; Wang, X.T.; Zeng, Q.D.; et al. Epistatic interaction effect between chromosome 1BL (Yr29) and a novel locus on 2AL facilitating resistance to stripe rust in Chinese wheat Changwu 357-9. Theor. Appl. Genet. 2022, 135, 2501–2513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.W.; Qiao, L.; Liu, Y.; Wei, N.C.; Zhao, J.J.; Wu, B.B.; Yang, B.; Wang, J.L.; Zheng, J. Genome-wide association study of grain number in common wheat from Shanxi under different water regimes. Front. Plant Sci. 2022, 12, 806295. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Li, H.L.; Wang, J.; Zhao, J.J.; Zheng, X.W.; Wu, B.B.; Du, W.J.; Wang, J.L.; Zheng, J. Analysis of genetic regions related to field grain number per spike from Chinese wheat founder parent Linfen 5064. Front. Plant Sci. 2022, 12, 808136. [Google Scholar] [CrossRef] [PubMed]
- Link, W.; Dixkens, C.; Singh, M.; Schwall, M.; Melchinger, A.E. Genetic diversity in European and Mediterranean faba bean germplasm revealed by RAPD markers. Theor. Appl. Genet. 1995, 90, 27–32. [Google Scholar] [CrossRef]
- Tufan, H.; Erdoğan, C. Genetic diversity in some faba bean (Vicia faba L.) genotypes assessed by simple sequence repeats. Biotechnol. Biotechnol. Equip. 2016, 31, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Kimber, R.B.E.; Cogan, N.O.I.; Materne, M.; Forster, J.W.; Paull, J.G. SNP discovery and high-density genetic mapping in faba bean (Vicia faba L.) permits identification of QTLs for ascochyta blight resistance. Plant Sci. 2014, 217–218, 47–55. [Google Scholar] [CrossRef]
- El-Esawi, M.A. SSR analysis of genetic diversity and structure of the germplasm of faba bean (Vicia faba L.). Comptes Rendus Biol. 2017, 340, 474–480. [Google Scholar] [CrossRef]
- Göl, Ş.; Doğanlar, S.; Frary, A. Relationship between geographical origin, seed size and genetic diversity in faba bean (Vicia faba L.) as revealed by SSR markers. Mol. Genet. Genom. 2017, 292, 991–999. [Google Scholar] [CrossRef]
- Yang, T.; Jiang, J.Y.; Zhang, H.Y.; Liu, R.; Strelkov, S.; Hwang, S.; Chang, K.F.; Yang, F.; Miao, Y.M.; He, Y.H.; et al. Density Enhancement of a Faba Bean Genetic Linkage Map (Vicia Faba) Based on Simple Sequence Repeats Markers. Plant Breed. 2019, 138, 207–215. [Google Scholar] [CrossRef]
- Sokol, E.; Nijenhuis, M.; Sjollema, K.A.; Jonkman, M.F.; Pas, H.H.; Giepmans, B.N. Particle bombardment of ex vivo skin to deliver DNA and express proteins. In Inflammation: Methods and Protocols; Clausen, B.E., Laman, J.D., Eds.; Humana Press: New York, NY, USA, 2017; pp. 107–118. [Google Scholar] [CrossRef]
- Gentile, F.; Arcaro, A.; Pizzimenti, S.; Daga, M.; Cetrangolo, G.P.; Dianzani, C.; Lepore, A.; Graf, M.; Ames, P.R.J.; Barrera, G. DNA damage by lipid peroxidation products: Implications in cancer, inflammation and autoimmunity. AIMS Genet. 2017, 04, 103–137. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, R.; Liu, Y.J.; Hou, W.W.; Wang, X.J.; Miao, Y.M.; He, Y.H.; Ma, Y.; Li, G.; Wang, D.; et al. Development and application of the Faba_bean_130K targeted next-generation sequencing SNP genotyping platform based on transcriptome sequencing. Theor. Appl. Genet. 2021, 134, 3195–3207. [Google Scholar] [CrossRef]
- Deng, Y.Q.; Xu, H.D.; Su, Y.L.; Liu, S.L.; Xu, L.W.; Guo, Z.X.; Wu, J.J.; Cheng, C.H.; Feng, J. Horizontal gene transfer contributes to virulence and antibiotic resistance of Vibrio harveyi 345 based on complete genome sequence analysis. BMC Genom. 2019, 20, 761–779. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.S.; Gore, M.; Buckler, E.S.; Yu, J.M. Status and prospects of association mapping in plants. Plant Genome 2008, 1, 5–20. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Si, Z.F.; Jin, S.K.; Li, J.Y.; Han, Z.G.; Li, Y.Q.; Wu, X.N.; Ge, Y.X.; Fang, L.; Zhang, T.Z.; Hu, Y. The design, validation, and utility of the “ZJU CottonSNP40K” liquid chip through genotyping by target sequencing. Ind. Crop. Prod. 2022, 188, 115629. [Google Scholar] [CrossRef]
- Liu, Y.C.; Liu, S.L.; Zhang, Z.F.; Ni, L.B.; Chen, X.M.; Ge, Y.X.; Zhou, G.A.; Tian, Z.X. GenoBaits Soy40K: A highly flexible and low-cost SNP array for soybean studies. Sci. China Life Sci. 2022, 65, 1898–1901. [Google Scholar] [CrossRef]
- Terzopoulos, P.J.; Bebeli, P.J. Genetic diversity analysis of Mediterranean faba bean (Vicia faba L.) with ISSR markers. Field Crop. Res. 2008, 108, 39–44. [Google Scholar] [CrossRef]
- Zong, X.X.; Liu, X.J.; Guan, J.P.; Wang, S.M.; Liu, Q.C.; Paull, J.G.; Redden, R. Molecular variation among Chinese and global winter faba bean germplasm. Theor. Appl. Genet. 2009, 118, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.X.; Ren, J.Y.; Guan, J.P.; Wang, S.M.; Liu, Q.C.; Paull, J.G.; Redden, R. Molecular variation among Chinese and global germplasm in spring faba bean areas. Plant Breed. 2009, 129, 508–513. [Google Scholar] [CrossRef]
- Gong, Y.M.; Xu, S.C.; Mao, W.H.; Li, Z.Y.; Hu, Q.Z.; Zhang, G.W.; Ding, J. Genetic diversity analysis of faba bean (Vicia faba L.) based on EST-SSR markers. Agric. Sci. China 2011, 10, 838–844. [Google Scholar] [CrossRef]
- Wang, H.F.; Zong, X.X.; Guan, J.P.; Yang, T.; Sun, X.L.; Ma, Y.; Redden, R. Genetic diversity and relationship of global faba bean (Vicia faba L.) germplasm revealed by ISSR markers. Theor. Appl. Genet. 2011, 124, 789–797. [Google Scholar] [CrossRef]
- Ladizinsky, G. On the origin of the broad bean, Vicia faba L. Isr. J. Bot. 1975, 24, 80–88. [Google Scholar]
- Zheng, Z.J. Food Legumes in China; Chinese Agricultural Press: Beijing, China, 1997; ISBN 7-109-04257-X. [Google Scholar]
- Cubero, J.I. On the evolution of Vicia faba L. Theor. Appl. Genet. 1974, 45, 47–51. [Google Scholar] [CrossRef]
- Zeid, M.; Schön, C.C.; Link, W. Genetic diversity in recent elite faba bean lines using AFLP markers. Theor. Appl. Genet. 2003, 107, 1304–1314. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Origin (Continent) | Origin (Country or Province) and No. of Accessions |
|---|---|
| Africa | Algeria (5), Egypt (7), Tunisia (5), Sudan (6), Morocco (2), Ethiopia (7) |
| North America | Canada (24), USA (1) |
| South America | Uruguay (1) |
| Europe | Russia (7), Poland (1), Hungary (2), Bulgaria (1), Spain (12), Greece (1), Britain (29), Holland (9), France (12), Germany (39) |
| Asia (except China) | Japan (3), Nepal (1), India (1), Afghanistan (17), Iran (2), Lebanon (9), Cyprus (1), Syria (46), Iraq (20), Jordan (3), Turkey (20) |
| Asia (spring sowing area of China) | Gansu (16), Hebei (1), Inner Mongolia (6), Ningxia (4), Qinghai (8), Shanxi (4), Xinjiang (4) |
| Asia (winter sowing area of China) | Anhui (3), Guangxi (3), Guizhou (8), Hubei (5), Hunan (4), Jiangsu (11), Jiangxi (3), Sichuan (12), Yunnan (11), Zhejiang (9), Chongqing (4) |
| Populations | Accession No. | Origin (Country or Province) and No. of Accessions |
|---|---|---|
| Subpop 1 | 57 | Africa (4), Asia (except China) (24), Europe (17), spring sowing area of China (8), winter sowing area of China (4) |
| Subpop 2 | 74 | Africa (3), Asia (except China) (12), Europe (4), spring sowing area of China (4), winter sowing area of China (51) |
| Subpop 3 | 67 | Asia (except China) (17), Europe (15), America (5), spring sowing area of China (24), winter sowing area of China (6) |
| Subpop 4 | 113 | Africa (15), Asia (except China) (28), Europe (50), America (15), spring sowing area of China (2), winter sowing area of China (3) |
| Mixed subpop | 99 | Africa (10), Asia (except China) (42), Europe (27), America (6), spring sowing area of China (5), winter sowing area of China (9) |
| Populations | Accession No. | Origin (Country or Province) and No. of Accessions |
|---|---|---|
| Group 1 | 79 | Africa (3), Asia (except China) (39), Europe (26), spring sowing area of China (9), winter sowing area of China (2) |
| Group 2 | 102 | Africa (4), Asia (except China) (27), Europe (9), America (1), spring sowing area of China (5), winter sowing area of China (56) |
| Group 3 | 82 | Africa (5), Asia (except China) (25), Europe (19), America (3), spring sowing area of China (25), winter sowing area of China (5) |
| Group 4 | 147 | Africa (20), Asia (except China) (32), Europe (59), America (22), spring sowing area of China (4), winter sowing area of China (10) |
| Populations | Sample Size | Nei’s Gene Diversity | PIC |
|---|---|---|---|
| Africa | 32 | 0.4541 | 0.3495 |
| America | 26 | 0.4386 | 0.3397 |
| Asia (except China) | 123 | 0.4721 | 0.3603 |
| Europe | 113 | 0.4629 | 0.3550 |
| Spring sowing area of China | 43 | 0.4549 | 0.3500 |
| Winter sowing area of China | 73 | 0.4334 | 0.3368 |
| Populations | Sample Size | Nei’s Gene Diversity | PIC |
|---|---|---|---|
| Mixed subpop | 99 | 0.4723 | 0.3604 |
| Subpop 1 | 57 | 0.4467 | 0.3450 |
| Subpop 2 | 74 | 0.4156 | 0.3254 |
| Subpop 3 | 67 | 0.4439 | 0.3433 |
| Subpop 4 | 113 | 0.4426 | 0.3426 |
| Source | df | SS | MS | Est. Var. | Percentage of Variation |
|---|---|---|---|---|---|
| Among Pops | 5 | 954.546 | 190.909 | 1.083 | 1% |
| Within Pops | 404 | 49,195.268 | 121.770 | 121.770 | 99% |
| Total | 409 | 50,149.815 | 122.853 | 100% |
| Source | df | SSD | MSD | Est. Var. | Percentage of Variation |
|---|---|---|---|---|---|
| Among Pops | 3 | 754.650 | 251.550 | 1.311 | 1% |
| Within Pops | 406 | 49,395.165 | 121.663 | 121.663 | 99% |
| Total | 409 | 50,149.815 | 122.974 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, Y.; Zong, X.; Teng, C.; Hou, W.; Li, P.; Du, D. Genetic Diversity of Global Faba Bean Germplasm Resources Based on the 130K TNGS Genotyping Platform. Agronomy 2023, 13, 811. https://doi.org/10.3390/agronomy13030811
Zhang H, Liu Y, Zong X, Teng C, Hou W, Li P, Du D. Genetic Diversity of Global Faba Bean Germplasm Resources Based on the 130K TNGS Genotyping Platform. Agronomy. 2023; 13(3):811. https://doi.org/10.3390/agronomy13030811
Chicago/Turabian StyleZhang, Hongyan, Yujiao Liu, Xuxiao Zong, Changcai Teng, Wanwei Hou, Ping Li, and Dezhi Du. 2023. "Genetic Diversity of Global Faba Bean Germplasm Resources Based on the 130K TNGS Genotyping Platform" Agronomy 13, no. 3: 811. https://doi.org/10.3390/agronomy13030811