
Comparative Omics-Based Identification and Expression Analysis of a Two-Component System in Vigna radiata in Drought Stress

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Family Identification of Two-Component System in V. radiata
2.2. Prediction of TCS Gene Structure and Cis Regulatory Elements in V. radiata
2.3. Phylogenetic Analysis, Multiple Sequence Alignment, and Motif Recognition
2.4. Chromosomal Mapping, Gene Duplication, and Evolutionary Analysis of TCS Members in V. radiata
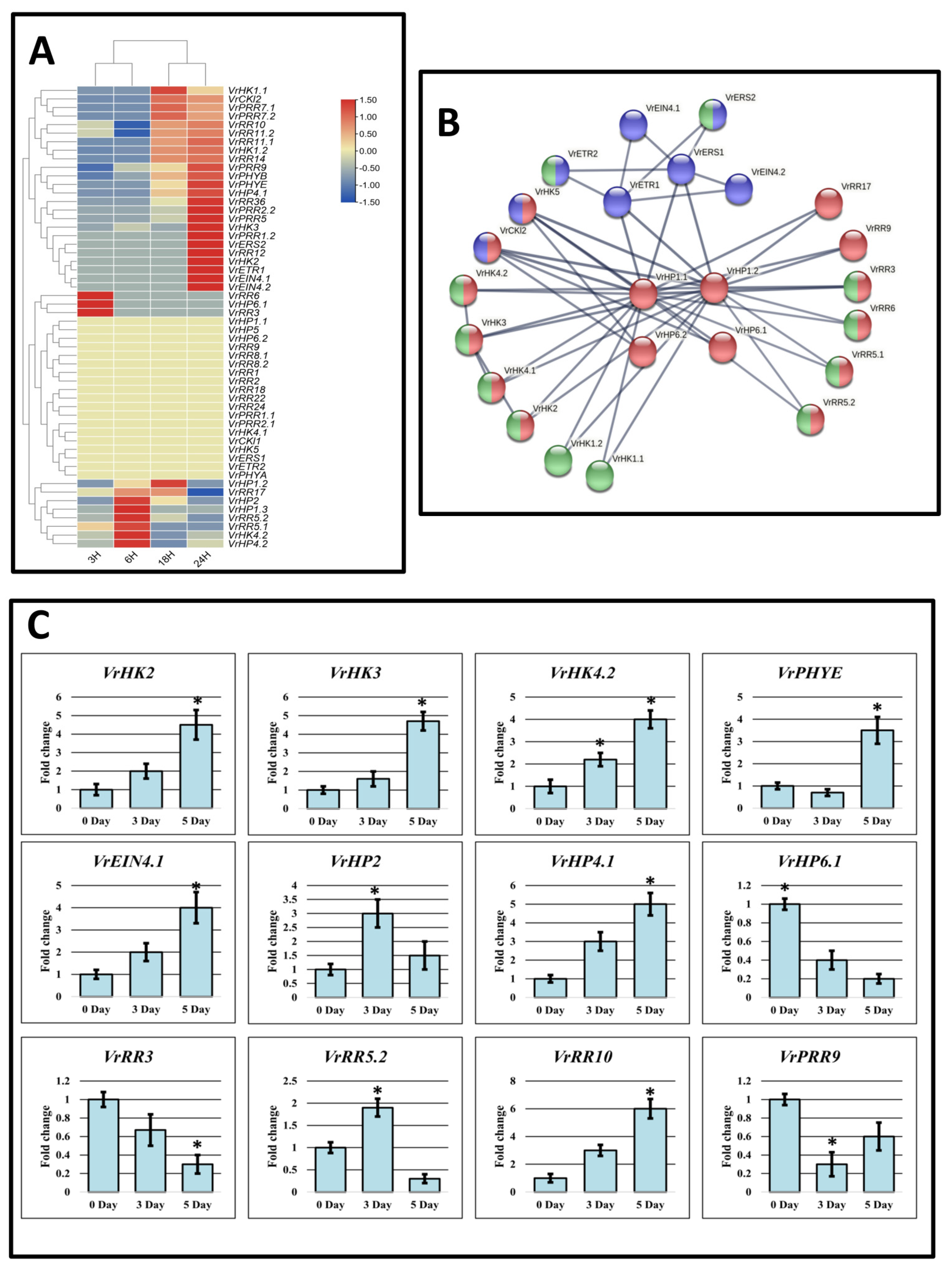
2.5. Two-Component System Gene Expression Patterns and Protein–Protein Interaction PPI in V. radiata
2.6. Plant Growth and Treatments
2.7. Validation of Expression Results Using Quantitative Real-Time PCR
3. Results
3.1. Comprehensive Identification of Two-Component System Genes in V. radiata
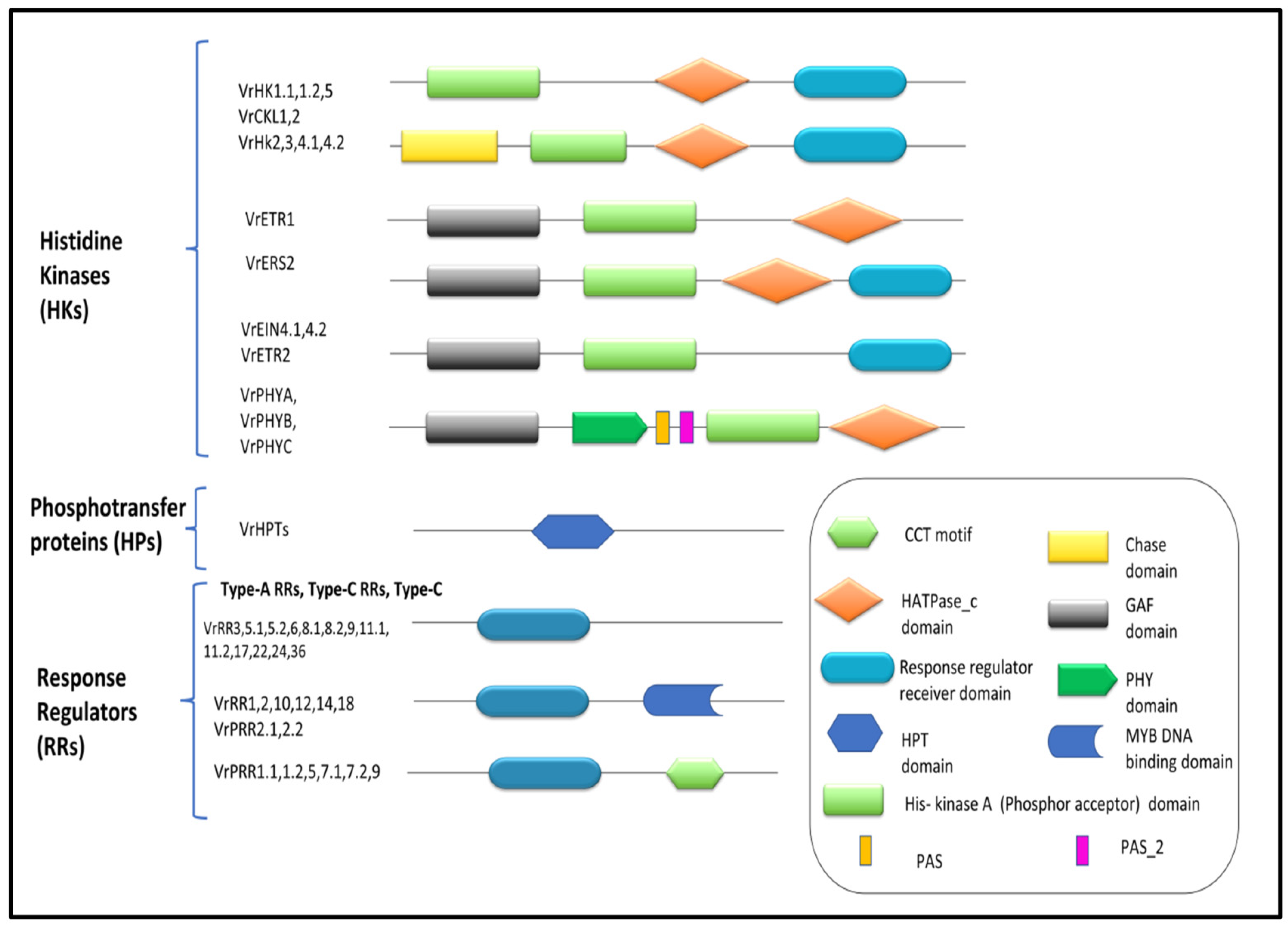
3.2. Histidine Kinase Protein Family in V. radiata
3.3. Histidine Phosphotransfer Protein (HpT) Family in V. radiata
3.4. The Response Regulators (RR) Family in V. radiata

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total No. of TCS Genes | HK | HP (Pseudo HP) | Type-A RRs | Type B RRs | Type C RRs | PRRS | Cotyledons | References |
|---|---|---|---|---|---|---|---|---|---|
| Arabidopsis thaliana | 47 | 8 | 6 (1) | 10 | 12 | 2 | 9 | Dicot | [15] |
| Oryza sativa L. | 37 | 5 | 5 | 15 | 7 | 0 | 5 | Monocot | [60] |
| Cicer arietinum | 51 | 18 | 7 (2) | 7 | 7 | 2 | 10 b | Dicot | [58] |
| Sorghum bicolor | 37 | 13 | 5 (2) | 3 | 7 | 2 | 7 b | Monocot | [59] |
| Glycine max | 98 | 36 | 13 | 18 | 15 | 3 | 13 | Dicot | [10] |
| Citrullus lanatus | 49 | 19 | 6 (2) | 8 | 10 | 6 | 5 | Dicot | [61] |
| Cucumis melo L. | 51 | 17 | 9 | 8 | 11 | 0 | 6 | Eudicots | [62] |
| Cucumis sativus L. | 46 | 18 | 7 (2) | 8 | 8 | 0 | 5 | Dicot | [61] |
| Populus trichocarpa | 49 | 12 | 12 | 9 | 11 | 0 | 5 | Dicot | [63] |
| Solanum lycopersicum | 65 | 20 | 6 (2) | 7 | 23 | 1 | 8 | Dicot | [64] |
| Physcomitrella patens | 39 | 18 | 3 | 7 | 5 | 2 | 4 a | Monocot | [65] |
| Zea mays | 59 | 11 | 9 (2) | 16 | 9 | 3 | 11 a | Monocot | [66] |
| Zizania latifolia | 69 | 25 | 8 | 14 | 14 | 2 | 6 | Monocot | [67] |
| Lotus japonicus | 40 | 14 | 7 | 7 | 11 | 1 | 5 a | Dicot | [20] |
| Brassica rapa | 85 | 20 | 8 (1) | 21 | 17 | 4 | 15 | Dicot | [68] |
| Triticum aestivum | 62 | 7 | 10 | 41 | 2 | 0 | 2 | Monocot | [69] |
| V. radiata | 54 | 18 | 9(2) | 8 | 8 | 3 | 8 | Dicot | Present work |
3.5. Features of VrTCS
3.6. Gene Structure and Conserved Motif Analysis of V. radiata TCS Members
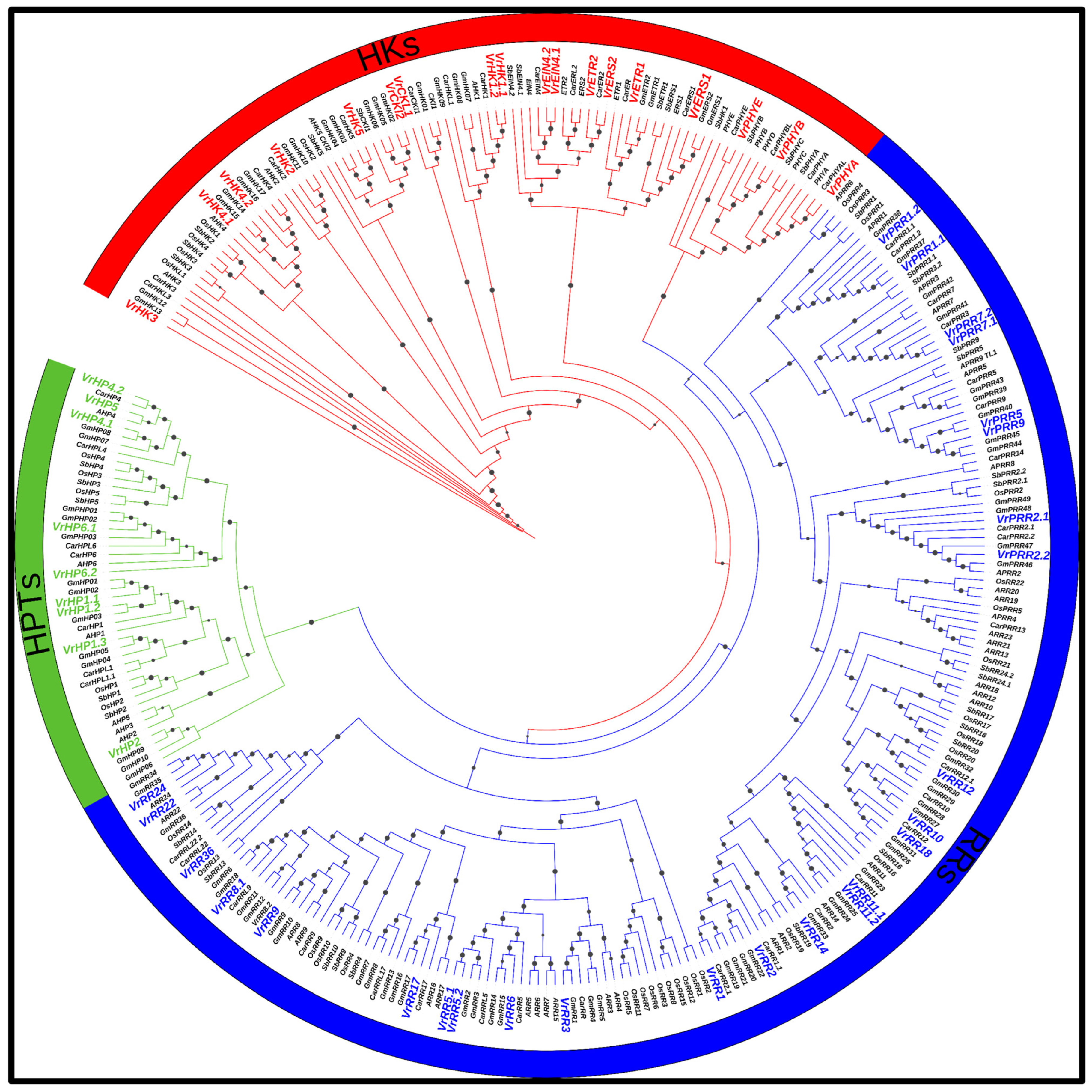
3.7. Phylogenetic Analysis of V. radiata TCS Members
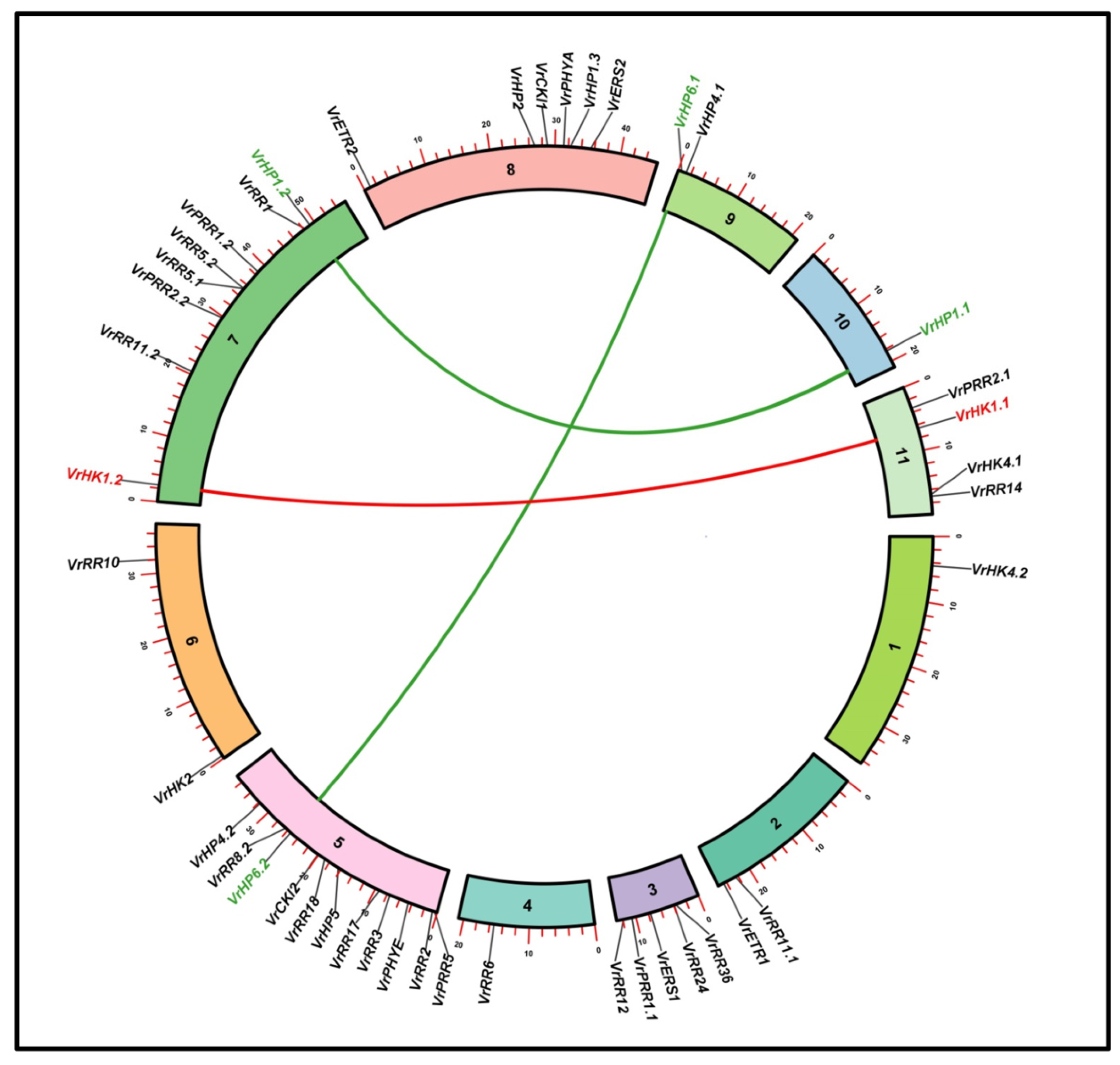
3.8. Synteny Analysis, Gene Distribution, and Duplication of V. radiata TCS Members
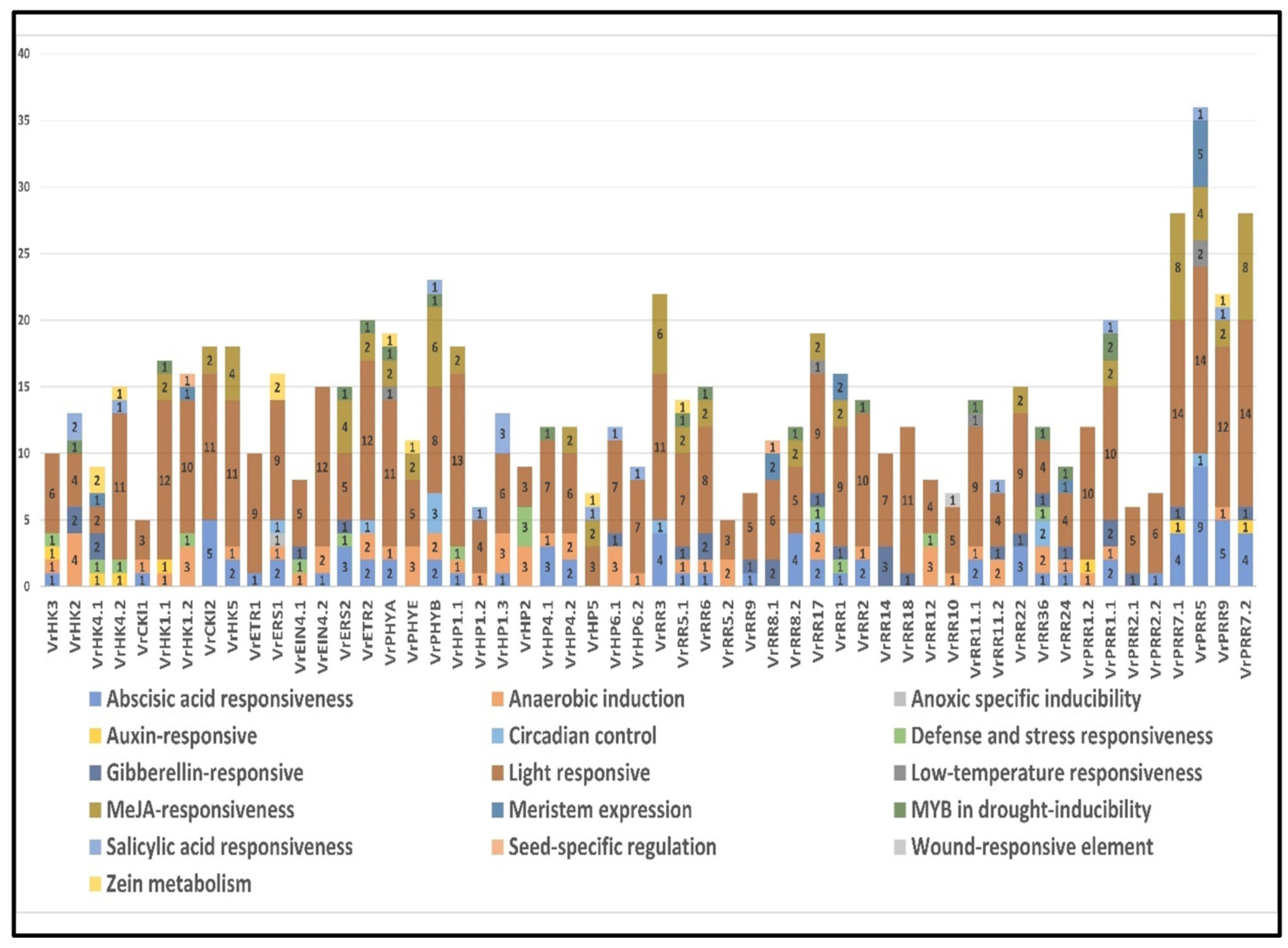
3.9. Promoter Analysis of V. radiata TCS Genes
3.10. Expression Analysis of V. radiata TCS Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kieber, J.J.; Schaller, G.E. Cytokinin Signaling in Plant Development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, C.; Yang, H.; Jiao, Y. Cytokinin Pathway Mediates APETALA1 Function in the Establishment of Determinate Floral Meristems in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 6840–6845. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Schmülling, T. Cytokinin Action in Plant Development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Sheen, J. Two-Component Circuitry in Arabidopsis Cytokinin Signal Transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Urao, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two-Component Systems in Plant Signal Transduction. Trends Plant Sci. 2000, 5, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Thomason, P.; Kay, R. Eukaryotic Signal Transduction via Histidine-Aspartate Phosphorelay. J. Cell Sci. 2000, 113, 3141–3150. [Google Scholar] [CrossRef]
- Mizuno, T. Plant Response Regulators Implicated in Signal Transduction and Circadian Rhythm. Curr. Opin. Plant Biol. 2004, 7, 499–505. [Google Scholar] [CrossRef]
- Schaller, G.E.; Kieber, J.J.; Shiu, S.-H. Two-Component Signaling Elements and Histidyl-Aspartyl Phosphorelays. Arab. Book 2008, 6, e0112. [Google Scholar] [CrossRef]
- Hwang, I.; Chen, H.-C.; Sheen, J. Two-Component Signal Transduction Pathways in Arabidopsis. Plant Physiol. 2002, 129, 500–515. [Google Scholar] [CrossRef]
- Mochida, K.; Yoshida, T.; Sakurai, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Genome-Wide Analysis of Two-Component Systems and Prediction of Stress-Responsive Two-Component System Members in Soybean. DNA Res. 2010, 17, 303–324. [Google Scholar] [CrossRef]
- Kiba, T.; Naitou, T.; Koizumi, N.; Yamashino, T.; Sakakibara, H.; Mizuno, T. Combinatorial Microarray Analysis Revealing Arabidopsis Genes Implicated in Cytokinin Responses through the His→Asp Phosphorelay Circuitry. Plant Cell Physiol. 2005, 46, 339–355. [Google Scholar] [CrossRef]
- Lohrmann, J.; Buchholz, G.; Keitel, C.; Sweere, U.; Kircher, S.; Bäurle, I.; Kudla, J.; Schäfer, E.; Harter, K. Differential Expression and Nuclear Localization of Response Regulator-Like Proteins from Arabidopsis Thaliana. Plant Biol. 1999, 1, 495–505. [Google Scholar] [CrossRef]
- Mähönen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Törmäkangas, K.; Ikeda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin Signaling and Its Inhibitor AHP6 Regulate Cell Fate During Vascular Development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef]
- Gupta, P.; Yadav, C.; Singh, D.; Nongpiur, R.C.; Singla-Pareek, S.L.; Pareek, A. Two Component Mediated Stress Signaling in Plants: A Comparative Profiling in Monocots and Dicots. In Protein Kinases and Stress Signaling in Plants; Pandey, G.K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 1–19. ISBN 978-1-119-54157-8. [Google Scholar]
- Hutchison, C.E.; Li, J.; Argueso, C.; Gonzalez, M.; Lee, E.; Lewis, M.W.; Maxwell, B.B.; Perdue, T.D.; Schaller, G.E.; Alonso, J.M.; et al. The Arabidopsis Histidine Phosphotransfer Proteins Are Redundant Positive Regulators of Cytokinin Signaling. Plant Cell 2006, 18, 3073–3087. [Google Scholar] [CrossRef] [PubMed]
- West, A.H.; Stock, A.M. Histidine Kinases and Response Regulator Proteins in Two-Component Signaling Systems. Trends Biochem. Sci. 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Tiwari, M.; Yadav, M.; Singh, B.; Pandey, V.; Nawaz, K.; Bhatia, S. Evolutionary and Functional Analysis of Two-component System in Chickpea Reveals CaRR13, a TypeB RR, as Positive Regulator of Symbiosis. Plant Biotechnol. J. 2021, 19, 2415–2427. [Google Scholar] [CrossRef] [PubMed]
- Gattolin, S.; Alandete-Saez, M.; Elliott, K.; Gonzalez-Carranza, Z.; Naomab, E.; Powell, C.; Roberts, J.A. Spatial and Temporal Expression of the Response Regulators ARR22 and ARR24 in Arabidopsis Thaliana. J. Exp. Bot. 2006, 57, 4225–4233. [Google Scholar] [CrossRef]
- Makino, S.; Kiba, T.; Imamura, A.; Hanaki, N.; Nakamura, A.; Suzuki, T.; Taniguchi, M.; Ueguchi, C.; Sugiyama, T.; Mizuno, T. Genes Encoding Pseudo-Response Regulators: Insight into His-to-Asp Phosphorelay and Circadian Rhythm in Arabidopsis Thaliana. Plant Cell Physiol. 2000, 41, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Niwa, Y.; Yamashino, T.; Mizuno, T. A Genome-Wide Compilation of the Two-Component Systems in Lotus Japonicus. DNA Res. 2009, 16, 237–247. [Google Scholar] [CrossRef]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Histidine Kinases in Plants: Cross Talk between Hormone and Stress Responses. Plant Signal. Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Genome-Wide Expression Profiling of Soybean Two-Component System Genes in Soybean Root and Shoot Tissues under Dehydration Stress. DNA Res. 2011, 18, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Großkinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The Role of Cis -Zeatin-Type Cytokinins in Plant Growth Regulation and Mediating Responses to Environmental Interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [PubMed]
- Zdarska, M.; Dobisová, T.; Gelová, Z.; Pernisová, M.; Dabravolski, S.; Hejátko, J. Illuminating Light, Cytokinin, and Ethylene Signalling Crosstalk in Plant Development. J. Exp. Bot. 2015, 66, 4913–4931. [Google Scholar] [CrossRef]
- Singh, D.; Singla-Pareek, S.L.; Pareek, A. Two-Component Signaling System in Plants: Interaction Network and Specificity in Response to Stress and Hormones. Plant Cell Rep. 2021, 40, 2037–2046. [Google Scholar] [CrossRef]
- Tran, L.-S.P.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Role of Cytokinin Responsive Two-Component System in ABA and Osmotic Stress Signalings. Plant Signal. Behav. 2010, 5, 148–150. [Google Scholar] [CrossRef]
- Nair, R.M.; Schafleitner, R.; Lee, S.-H. (Eds.) The Mungbean Genome; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2020; ISBN 978-3-030-20007-7. [Google Scholar]
- Cao, D.; Li, H.; Yi, J.; Zhang, J.; Che, H.; Cao, J.; Yang, L.; Zhu, C.; Jiang, W. Antioxidant Properties of the Mung Bean Flavonoids on Alleviating Heat Stress. PLoS ONE 2011, 6, e21071. [Google Scholar] [CrossRef] [PubMed]
- Uppalwar, S.V.; Garg, V.; Dutt, R. Seeds of Mung Bean (Vigna radiata (L.) R.Wilczek): Taxonomy, Phytochemistry, Medicinal Uses and Pharmacology. Curr. Bioact. Compd. 2021, 17, 220–233. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, M.; Wu, R.; Chen, X.; Liu, F.; Xing, B. Genome-Wide Analysis of OSCA Gene Family Members in Vigna Radiata and Their Involvement in the Osmotic Response. BMC Plant Biol. 2021, 21, 408. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, N.; Yadav, M.; Sharma, A.K.; Kumar, V.; Bhat, K.V. Salt Stress and Mungbean (Vigna radiata (L.) Wilczek): Effects, Physiological Perspective and Management Practices for Alleviating Salinity. Arch. Agron. Soil Sci. 2019, 65, 1287–1301. [Google Scholar] [CrossRef]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomics Data. In Plant Bioinformatics; Edwards, D., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2016; Volume 1374, pp. 115–140. ISBN 978-1-4939-3166-8. [Google Scholar]
- Gish, W.; States, D.J. Identification of Protein Coding Regions by Database Similarity Search. Nat. Genet. 1993, 3, 266–272. [Google Scholar] [CrossRef]
- Bateman, A. The Pfam Protein Families Database. Nucleic Acids Res. 2004, 32, D138–D141. [Google Scholar] [CrossRef] [PubMed]
- Hulo, N. The PROSITE Database. Nucleic Acids Res. 2006, 34, D227–D230. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J. SMART: A Web-Based Tool for the Study of Genetically Mobile Domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; von Heijne, G.; Brunak, S. Feature-Based Prediction of Non-Classical and Leaderless Protein Secretion. Protein Eng. Des. Sel. 2004, 17, 349–356. [Google Scholar] [CrossRef]
- Yu, C.-S.; Cheng, C.-W.; Su, W.-C.; Chang, K.-C.; Huang, S.-W.; Hwang, J.-K.; Lu, C.-H. CELLO2GO: A Web Server for Protein SubCELlular LOcalization Prediction with Functional Gene Ontology Annotation. PLoS ONE 2014, 9, e99368. [Google Scholar] [CrossRef]
- Brown, G.R.; Hem, V.; Katz, K.S.; Ovetsky, M.; Wallin, C.; Ermolaeva, O.; Tolstoy, I.; Tatusova, T.; Pruitt, K.D.; Maglott, D.R.; et al. Gene: A Gene-Centered Information Resource at NCBI. Nucleic Acids Res. 2015, 43, D36–D42. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.-C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-Seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef]
- Mering, C.V. STRING: A Database of Predicted Functional Associations between Proteins. Nucleic Acids Res. 2003, 31, 258–261. [Google Scholar] [CrossRef]
- Bakhtiarizadeh, M.R.; Najaf-Panah, M.J.; Mousapour, H.; Salami, S.A. Versatility of Different Melting Temperature (Tm) Calculator Software for Robust PCR and Real-Time PCR Oligonucleotide Design: A Practical Guide. Gene Rep. 2016, 2, 1–3. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Bas, A.; Forsberg, G.; Hammarstrom, S.; Hammarstrom, M.-L. Utility of the Housekeeping Genes 18S RRNA, Beta-Actin and Glyceraldehyde-3-Phosphate-Dehydrogenase for Normalization in Real-Time Quantitative Reverse Transcriptase-Polymerase Chain Reaction Analysis of Gene Expression in Human T Lymphocytes. Scand. J. Immunol. 2004, 59, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-R.; Nam, H.-Y.; Kim, S.-U.; Kim, S.-I.; Chang, Y.-J. Normalization of Reverse Transcription Quantitative-PCR with Housekeeping Genes in Rice. Biotechnol. Lett. 2003, 25, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Stadler, R. Ethylene Hormone Receptor Action InArabidopsis. Bioessays 2001, 23, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Clack, T.; Mathews, S.; Sharrock, R.A. The Phytochrome Apoprotein Family InArabidopsis Is Encoded by Five Genes: The Sequences and Expression OfPHYD AndPHYE. Plant Mol. Biol. 1994, 25, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Azeem, F.; Ali, M.A.; Nawaz, M.A.; Nadeem, H.; Abbas, A.; Batool, R.; Atif, R.M.; Ijaz, U.; Nieves-Cordones, M.; et al. Genome-Wide Identification and Expression Analysis of Two Component System Genes in Cicer Arietinum. Genomics 2020, 112, 1371–1383. [Google Scholar] [CrossRef]
- Zameer, R.; Sadaqat, M.; Fatima, K.; Fiaz, S.; Rasul, S.; Zafar, H.; Qayyum, A.; Nashat, N.; Raza, A.; Shah, A.N.; et al. Two-Component System Genes in Sorghum Bicolor: Genome-Wide Identification and Expression Profiling in Response to Environmental Stresses. Front. Genet. 2021, 12, 794305. [Google Scholar] [CrossRef]
- Du, L.; Jiao, F.; Chu, J.; Jin, G.; Chen, M.; Wu, P. The Two-Component Signal System in Rice (Oryza sativa L.): A Genome-Wide Study of Cytokinin Signal Perception and Transduction. Genomics 2007, 89, 697–707. [Google Scholar] [CrossRef]
- He, Y.; Liu, X.; Zou, T.; Pan, C.; Qin, L.; Chen, L.; Lu, G. Genome-Wide Identification of Two-Component System Genes in Cucurbitaceae Crops and Expression Profiling Analyses in Cucumber. Front. Plant Sci. 2016, 7, 899. [Google Scholar] [CrossRef]
- Liu, P.; Wang, S.; Wang, X.; Yang, X.; Li, Q.; Wang, C.; Chen, C.; Shi, Q.; Ren, Z.; Wang, L. Genome-Wide Characterization of Two-Component System (TCS) Genes in Melon (Cucumis melo L.). Plant Physiol. Biochem. 2020, 151, 197–213. [Google Scholar] [CrossRef]
- Singh, G.; Kumar, R. Genome-Wide Insilico Analysis of Plant Two Component Signaling System in Woody Model Plant Populus Trichocarpa. Res. Plant Biol. 2012, 2, 13–23. [Google Scholar]
- Firon, N.; Pressman, E.; Meir, S.; Khoury, R.; Altahan, L. Ethylene Is Involved in Maintaining Tomato (Solanum lycopersicum) Pollen Quality under Heat-Stress Conditions. AoB Plants 2012, 2012, pls024. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Yamashino, T.; Nakanishi, H.; Mizuno, T. Classification of the Genes Involved in the Two-Component System of the Moss Physcomitrella patens. Biosci. Biotechnol. 2010, 74, 2542–2545. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.X.; Ma, Q.; Lin, Y.X.; Tang, X.L.; Zhou, Y.Q.; Zhu, S.W.; Fan, J.; Cheng, B.J. Genome-Wide Identification, Classification, and Analysis of Two-Component Signal System Genes in Maize. Genet. Mol. Res. 2011, 10, 3316–3330. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, F.; Wu, X.; Hu, Y.; Dong, L.; Dewitte, W.; Wen, B. Genome-Wide Characterization and Expression of Two-Component System Genes in Cytokinin-Regulated Gall Formation in Zizania Latifolia. Plants 2020, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, M.; Kong, L.; Lv, Y.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Genome-Wide Identification, Phylogeny, Duplication, and Expression Analyses of Two-Component System Genes in Chinese Cabbage (Brassica rapa ssp. Pekinensis). DNA Res. 2014, 21, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Gahlaut, V.; Mathur, S.; Dhariwal, R.; Khurana, J.P.; Tyagi, A.K.; Balyan, H.S.; Gupta, P.K. A Multi-Step Phosphorelay Two-Component System Impacts on Tolerance against Dehydration Stress in Common Wheat. Funct. Integr. Genomics. 2014, 14, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Urao, T.; Yakubov, B.; Satoh, R.; Yamaguchi-Shinozaki, K.; Seki, M.; Hirayama, T.; Shinozaki, K. A Transmembrane Hybrid-Type Histidine Kinase in Arabidopsis Functions as an Osmosensor. Plant Cell 1999, 11, 1743–1754. [Google Scholar] [CrossRef]
- Franklin, K.A.; Larner, V.S.; Whitelam, G.C. The Signal Transducing Photoreceptors of Plants. Int. J. Dev. Biol. 2005, 49, 653–664. [Google Scholar] [CrossRef]
- Filiault, D.L.; Wessinger, C.A.; Dinneny, J.R.; Lutes, J.; Borevitz, J.O.; Weigel, D.; Chory, J.; Maloof, J.N. Amino Acid Polymorphisms in Arabidopsis Phytochrome B Cause Differential Responses to Light. Proc. Natl. Acad. Sci. USA 2008, 105, 3157–3162. [Google Scholar] [CrossRef]
- Farré, E.M.; Liu, T. The PRR Family of Transcriptional Regulators Reflects the Complexity and Evolution of Plant Circadian Clocks. Curr. Opin. Plant Biol. 2013, 16, 621–629. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ruan, J.; Ho, T.-H.D.; You, Y.; Yu, T.; Quatrano, R.S. Cis-Regulatory Element Based Targeted Gene Finding: Genome-Wide Identification of Abscisic Acid- and Abiotic Stress-Responsive Genes in Arabidopsis Thaliana. Bioinformatics 2005, 21, 3074–3081. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-P.; Jiang, X.-L.; Zhang, B.-Y.; Su, X.-H. Involvement of MicroRNA-Mediated Gene Expression Regulation in the Pathological Development of Stem Canker Disease in Populus Trichocarpa. PLoS ONE 2012, 7, e44968. [Google Scholar] [CrossRef] [PubMed]






| Locus ID | Gene Name | Isoelectric Point (PI) | Molecular Weight (Da) | Instability Index | Aliphatic Index | GRAVY | Protein Length (Amino Acids) | Chr No. | No. of Exon | G Start | G End | Cellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HK Family | ||||||||||||
| LOC106755439 | VrHK3 | 6.7 | 114,682 | 37.61 | 93.55 | −0.107 | 1028 | Un | 10 | 110,644 | 118,425 | Plasma Membrane |
| LOC106764676 | VrHK2 | 7.2 | 134,098 | 43.76 | 87.55 | −0.22 | 1190 | C6 | 14 | 479,116 | 488,795 | Plasma Membrane |
| LOC106777844 | VrHK4.1 | 6.67 | 110,950 | 36.19 | 89.42 | −0.124 | 990 | C11 | 14 | 16,816,322 | 1,683,310 | Cytoplasmic |
| LOC106770782 | VrHK4.2 | 6.82 | 111,430 | 33.66 | 86.49 | −0.174 | 999 | C1 | 12 | 4,438,961 | 4,447,994 | Cytoplasmic |
| LOC106770468 | VrCKl1 | 6.82 | 111,430 | 33.98 | 90.75 | −0.115 | 1060 | C8 | 7 | 28,809,958 | 28,814,950 | Plasma Membrane |
| LOC106777758 | VrHK1.1 | 6.18 | 141,439 | 41.26 | 90.86 | −0.223 | 1265 | C11 | 13 | 6,536,759 | 6,543,727 | Plasma Membrane |
| LOC106768916 | VrHK1.2 | 8.31 | 137,838 | 40.22 | 89.76 | −0.253 | 1234 | C7 | 13 | 2,505,103 | 2,511,396 | Plasma Membrane |
| LOC106761316 | VrCKl2 | 4.99 | 109,374 | 56.97 | 82.21 | −0.445 | 972 | C5 | 13 | 20,096,528 | 20,101,731 | Nuclear |
| LOC106763060 | VrHK5 | 5.15 | 114,127 | 54.95 | 77.67 | −0.567 | 1011 | C6 | 13 | 315,594,338 | 31,567,102 | Nuclear |
| LOC106756118 | VrETR1 | 8.31 | 83,132.2 | 40.46 | 107.2 | 0.122 | 742 | C2 | 7 | 24,449,047 | 24,457,030 | Plasma Membrane |
| LOC106757732 | VrERS1 | 6.36 | 70,848.1 | 41.01 | 102.4 | 0.031 | 636 | C3 | 7 | 7,678,537 | 7,683,030 | Plasma Membrane |
| LOC106754157 | VrEIN4.1 | 6.91 | 85,246.9 | 35.96 | 101 | 0.053 | 760 | Un | 2 | 1,092,832 | 1,096,427 | Plasma Membrane |
| LOC106754157 | VrEIN4.2 | 6.65 | 85,181.1 | 36.55 | 103.3 | 0.086 | 759 | Un | 2 | 1,098,989 | 1,102,570 | Plasma Membrane |
| LOC106772158 | VrERS2 | 8.05 | 83,779.4 | 41.93 | 103.9 | 0.085 | 757 | C8 | 5 | 35,402,755 | 35,407,049 | Plasma Membrane |
| LOC106772500 | VrETR2 | 6.28 | 84,271.9 | 33.78 | 97.53 | 0.111 | 760 | C8 | 3 | 1,031,509 | 1,035,564 | Plasma Membrane |
| LOC106771983 | VrPHYA | 6.09 | 123,961 | 47.23 | 93.94 | −0.079 | 1123 | C8 | 7 | 31,264,701 | 31,270,514 | Cytoplasmic |
| LOC106760765 | VrPHYE | 5.69 | 125,312 | 46.74 | 90.79 | −0.173 | 1121 | C5 | 4 | 4,573,654 | 4,579,012 | Cytoplasmic, Plasma Membrane, |
| LOC106754114 | VrPHYB | 5.69 | 125,677 | 45.85 | 93.98 | −0.116 | 1131 | Un | 4 | 10,541 | 17,443 | Cytoplasmic |
| HPTs | ||||||||||||
| LOC106775067 | VrHP1.1 | 4.8 | 17,631 | 42.53 | 88.65 | −0.254 | 155 | C10 | 7 | 18,301,338 | 16,303,340 | Cytoplasmic, Extracellular |
| LOC106767217 | VrHP1.2 | 4.74 | 17,736.2 | 43.93 | 90.45 | −0.223 | 154 | C7 | 6 | 49,140,073 | 49,141,539 | Cytoplasmic, Nuclear |
| LOC106771462 | VrHP1.3 | 5.03 | 16,565.7 | 37.7 | 94.73 | −0.34 | 146 | C8 | 6 | 32,351,139 | 32,352,533 | Nuclear |
| LOC106771501 | VrHP2 | 5.45 | 17,067.4 | 43.33 | 74.8 | −0.361 | 150 | C8 | 6 | 26,939,465 | 26,942,841 | Nuclear |
| LOC106773913 | VrHP4.1 | 8.68 | 17,413.7 | 62.07 | 71.85 | −0.746 | 151 | C9 | 6 | 1,290,483 | 1,292,013 | Nuclear |
| LOC106761115 | VrHP4.2 | 8.59 | 23,040.5 | 36.03 | 79.29 | −0.365 | 198 | C5 | 6 | 31,782,312 | 31,785,137 | Nuclear |
| LOC106760377 | VrHP5 | 4.53 | 12,940.5 | 49.06 | 70.27 | −0.48 | 111 | C5 | 4 | 16,465,489 | 16,467,894 | Nuclear, Cytoplasmic |
| LOC106773969 | VrHP6.1 | 5.94 | 17,971.5 | 44.3 | 90.71 | −0.271 | 156 | C9 | 6 | 547,257 | 548,917 | Extracellular |
| LOC106761820 | VrHP6.2 | 7.59 | 25,968.9 | 48.28 | 89.69 | −0.116 | 223 | C5 | 5 | 25,414,244 | 25,415,627 | Extracellular, Plasma Membrane |
| Type-A RRs | ||||||||||||
| LOC106760963 | VrRR3 | 4.81 | 25,479.8 | 66.93 | 88.06 | −0.309 | 232 | C5 | 5 | 7,566,240 | 7,568,017 | Nuclear |
| LOC106766897 | VrRR5.1 | 7.63 | 22,881.7 | 55.32 | 103.1 | −0.143 | 206 | C7 | 5 | 35,245,726 | 35,247,771 | Nuclear |
| LOC106758083 | VrRR6 | 7.7 | 21,905.2 | 57.57 | 91.09 | −0.116 | 202 | C4 | 5 | 15,357,967 | 15,359,998 | Nuclear, Chloroplast |
| LOC106767155 | VrRR5.2 | 6.76 | 22,762.5 | 55.51 | 102.1 | −0.121 | 206 | C7 | 5 | 35,254,959 | 35,257,043 | Nuclear, Cytoplasmic |
| LOC106752532 | VrRR9 | 5.69 | 27,500.9 | 71.26 | 74.98 | −0.828 | 243 | Un | 5 | 359,726 | 361,726 | Nuclear |
| LOC106779256 | VrRR8.1 | 5.72 | 20,486.6 | 59.82 | 88.72 | −0.401 | 179 | Un | 5 | 344,091 | 345,589 | Nuclear |
| LOC106760951 | VrRR8.2 | 4.95 | 24,924.7 | 68.98 | 93.93 | −0.276 | 219 | C5 | 4 | 26,430,472 | 26,431,946 | Nuclear, Plasma Membrane |
| LOC106759802 | VrRR17 | 8.69 | 15,785.5 | 47.54 | 94.69 | −0.208 | 143 | C5 | 5 | 9,332,686 | 9,333,984 | Nuclear, Extracellular |
| Type-B RRs | ||||||||||||
| LOC106768205 | VrRR1 | 5.84 | 77,305.4 | 45.05 | 82.15 | −0.393 | 706 | C7 | 7 | 48,240,050 | 48,244,152 | Nuclear |
| LOC106760138 | VrRR2 | 6.07 | 73,630.9 | 46.03 | 81.08 | −0.441 | 673 | C5 | 6 | 734,320 | 738,195 | Nuclear |
| LOC106777889 | VrRR14 | 6.15 | 68,215.7 | 56.73 | 74.84 | −0.49 | 620 | C11 | 7 | 17,020,135 | 17,027,339 | Nuclear |
| LOC106762533 | VrRR18 | 6.21 | 73,998.2 | 41.95 | 77.94 | −0.562 | 670 | C5 | 6 | 18,903,242 | 18,907,979 | Nuclear |
| LOC106757890 | VrRR12 | 5.44 | 73,702.2 | 48.21 | 76.48 | −0.516 | 677 | C3 | 6 | 11,915,194 | 11,920,109 | Nuclear |
| LOC106763713 | VrRR10 | 6.4 | 67,500.7 | 41.87 | 76.44 | −0.504 | 613 | C6 | 9 | 32,044,562 | 32,048,926 | Nuclear |
| LOC106777203 | VrRR11.1 | 5.57 | 64,930 | 48.37 | 76.71 | −0.527 | 578 | C2 | 6 | 22,436,393 | 22,440,444 | Nuclear |
| LOC106766713 | VrRR11.2 | 5.06 | 65,044.1 | 56.94 | 76.23 | −0.503 | 578 | C7 | 5 | 20,462,824 | 20,467,554 | Nuclear |
| Type-C RRs | ||||||||||||
| LOC106754035 | VrRR22 | 6.9 | 15,514.9 | 24.28 | 72.1 | −0.395 | 138 | Un | 2 | 964,858 | 965,390 | Cytoplasmic |
| LOC106757536 | VrRR36 | 8.49 | 16,082.7 | 26.09 | 84.48 | −0.275 | 143 | C3 | 3 | 3,714,256 | 3,715,525 | Chloroplast, Mitochondrial |
| LOC106756903 | VrRR24 | 7.94 | 14,994.3 | 47.14 | 88.65 | −0.382 | 133 | C3 | 2 | 3,954,702 | 3,955,257 | Nuclear, Mitochondrial |
| PRRs | ||||||||||||
| LOC106769035 | VrPRR1.2 | 5.67 | 63,861.3 | 53.14 | 73.33 | −0.642 | 570 | C7 | 7 | 38,576,824 | 38,582,446 | Nuclear |
| LOC106756669 | VrPRR1.1 | 5.95 | 62,408.7 | 57.49 | 66.68 | −0.724 | 557 | C3 | 6 | 10,571,692 | 10,576,727 | Nuclear |
| LOC106777717 | VrPRR2.1 | 5.89 | 62,525.5 | 56.81 | 67.57 | −0.668 | 560 | C11 | 13 | 3,330,688 | 3,336,996 | Nuclear |
| LOC106769136 | VrPRR2.2 | 6.22 | 62,034.5 | 54 | 71.81 | −0.573 | 554 | C7 | 13 | 29,778,486 | 29,785,007 | Nuclear |
| LOC106754416 | VrPRR7.1 | 7.54 | 80,057.9 | 43.73 | 68.81 | −0.734 | 737 | Un | 9 | 279,157 | 313,750 | Nuclear |
| LOC106760140 | VrPRR5 | 6.5 | 72,478.7 | 42.36 | 66.99 | −0.656 | 655 | C5 | 8 | 404,026 | 408,131 | Nuclear |
| LOC106780351 | VrPRR9 | 6.05 | 74,267.3 | 44.64 | 67.32 | −0.656 | 675 | Un | 10 | 15,087 | 20,882 | Nuclear |
| LOC106754416 | VrPRR7.2 | 7.54 | 81,386.4 | 43.67 | 69.52 | −0.721 | 749 | Un | 9 | 279,157 | 313,750 | Nuclear |
| Gene Duplication Pairs | Ka | Ks | Ka/Ks | Divergence Time Mya (Million Years Ago) | Type of Duplication |
|---|---|---|---|---|---|
| VrHK1./VrHK1.2 | 3.463 | 1.585 | 2.185 | 120.80 | Segmental |
| VrHP1.1/VrHP1.2 | 6.098 | 2.545 | 2.396 | 193.99 | Segmental |
| VrHP6.1/VrHP6.2 | 2.294 | 2.499 | 0.9180 | 190.46 | Segmental |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afzal, M.A.; Azeem, F.; Afzal, S.; Afzal, N.; Rizwan, M.; Seo, H.; Shah, A.A.; Nawaz, M.A. Comparative Omics-Based Identification and Expression Analysis of a Two-Component System in Vigna radiata in Drought Stress. Agronomy 2023, 13, 989. https://doi.org/10.3390/agronomy13040989
Afzal MA, Azeem F, Afzal S, Afzal N, Rizwan M, Seo H, Shah AA, Nawaz MA. Comparative Omics-Based Identification and Expression Analysis of a Two-Component System in Vigna radiata in Drought Stress. Agronomy. 2023; 13(4):989. https://doi.org/10.3390/agronomy13040989
Chicago/Turabian StyleAfzal, Muhammad Amin, Farrukh Azeem, Shumaila Afzal, Naila Afzal, Muhammad Rizwan, Hyojin Seo, Asad Ali Shah, and Muhammad Amjad Nawaz. 2023. "Comparative Omics-Based Identification and Expression Analysis of a Two-Component System in Vigna radiata in Drought Stress" Agronomy 13, no. 4: 989. https://doi.org/10.3390/agronomy13040989