
Eco-Physiological Behavior of Five Tunisian Olive Tree Cultivars under Drought Stress



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Relative Water Content
2.3. Stomatal Resistance
2.4. Chlorophyll Fluorescence
2.5. SPAD Index
2.6. Dry Matter Accumulation
2.7. Statistical Analysis
3. Results
3.1. Effect of Drought Stress on Relative Water Content
3.2. Effect of Drought Stress on Stomatal Resistance
3.3. Effect of Drought Stress on PSII Maximal Photochemical Efficiency (FV/FM) and Performance Index (PI)
3.4. Effect of Drought Stress on SPAD Index
3.5. Effect of Drought Stress on Dry Matter Accumulation
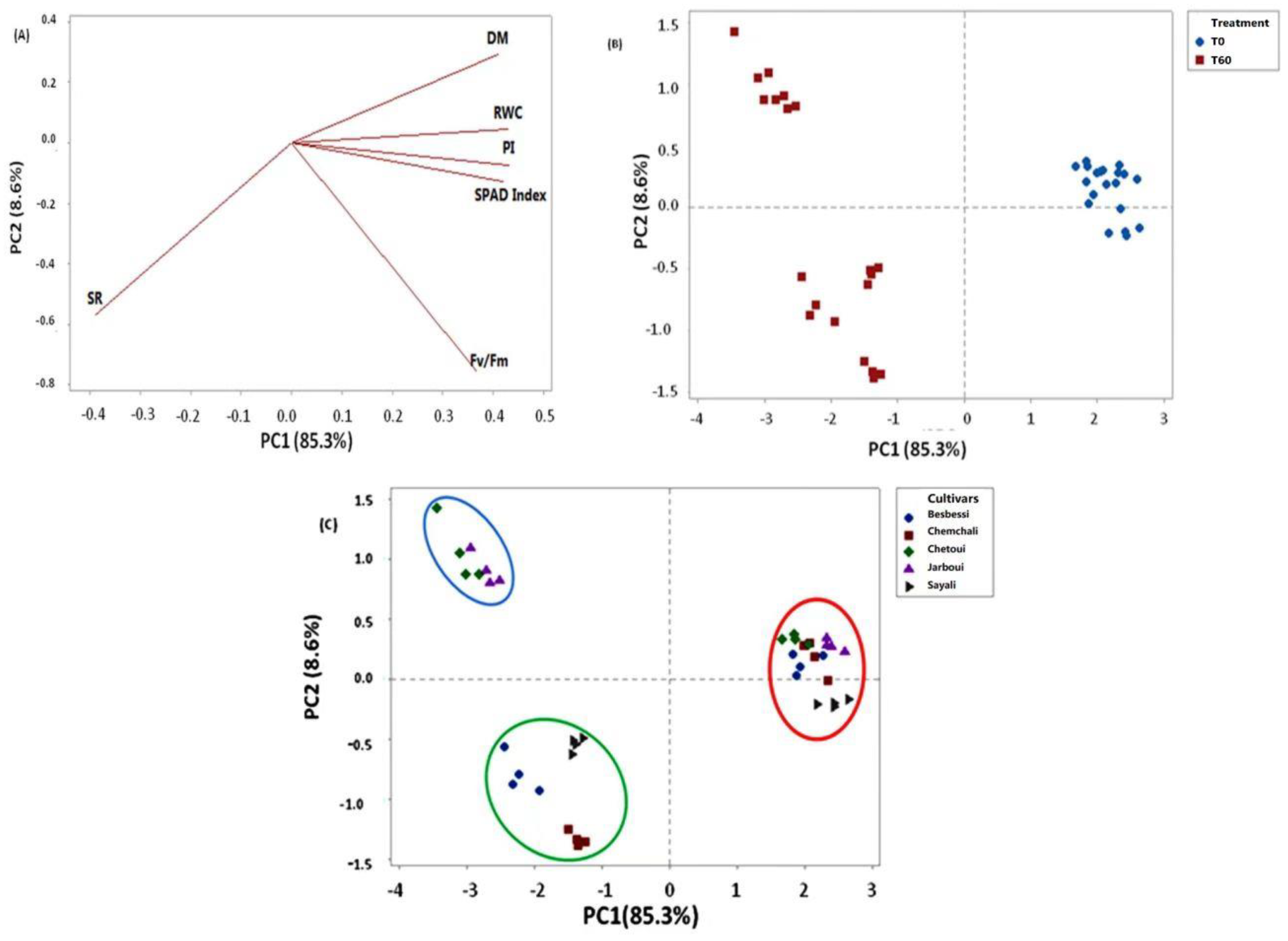
3.6. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lionello, P.; Scarascia, L. The Relation between Climate Change in the Mediterranean Region and Global Warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Abdulla, F.; Eshtawi, T.; Assaf, H. Assessment of the Impact of Potential Climate Change on the Water Balance of a Semi-Arid Watershed. Water Resour. Manag. 2009, 23, 2051–2068. [Google Scholar] [CrossRef]
- Rochdane, S.; Bounoua, L.; Zhang, P.; Imhoff, M.L.; Messouli, M.; Yacoubi-Khebiza, M. Combining Satellite Data and Models to Assess Vulnerability to Climate Change and Its Impact on Food Security in Morocco. Sustainabilty 2014, 6, 1729–1746. [Google Scholar] [CrossRef] [Green Version]
- Melki, A.; Abdollahi, K.; Fatahi, R.; Abida, H. Groundwater Recharge Estimation under Semi Arid Climate: Case of Northern Gafsa Watershed, Tunisia. J. Afr. Earth Sci. 2017, 132, 37–46. [Google Scholar] [CrossRef]
- Souissi, A.; Mtimet, N.; McCann, L.; Chebil, A.; Thabet, C. Determinants of Food Consumption Water Footprint in the MENA Region: The Case of Tunisia. Sustainability 2022, 14, 1539. [Google Scholar] [CrossRef]
- GTZ; MARH. Stratégie Nationale d’Adaptation de l’Agriculture Tunisienne et des Écosystèmes aux Changements Climatiques. 2007. Available online: https://www.giz.de/de/html/index.html (accessed on 12 December 2022).
- Amamou, H.; Ben, M.; Aouadi, H.; Khemiri, H.; Mahouachi, M.; Beckers, Y.; Hammami, H. Climate Risk Management Climate Change-Related Risks and Adaptation Strategies as Perceived in Dairy Cattle Farming Systems in Tunisia. Clim. Risk Manag. 2018, 20, 38–49. [Google Scholar] [CrossRef]
- Boughariou, E.; Allouche, N.; Jmal, I.; Mokadem, N.; Ayed, B.; Hajji, S.; Khanfir, H.; Bouri, S. Modeling Aquifer Behaviour under Climate Change and High Consumption: Case Study of the Sfax Region, Southeast Tunisia. J. Afr. Earth Sci. 2018, 141, 118–129. [Google Scholar] [CrossRef]
- Chebil, A.; Frija, A.; Makhlouf, M.; Thabet, C.; Jebari, S. Effects of Water Scarcity on the Performances of the Agricultural Sector and Adaptation Strategies in Tunisia. In Agricultural Economics; IntechOpen: London, UK, 2019. [Google Scholar]
- Ben Abdallah, S.; Elfkih, S.; Suárez-Rey, E.M.; Parra-López, C.; Romero-Gámez, M. Evaluation of the Environmental Sustainability in the Olive Growing Systems in Tunisia. J. Clean. Prod. 2021, 282, 124526. [Google Scholar] [CrossRef]
- Omri, A.; Abdelhamid, S.; Ayadi, M.; Araouki, A. The Investigation of Minor and Rare Tunisian Olive Cultivars to Enrich and Diversify the Olive Genetic Resources of the Country. J. Food Compos. Anal. 2021, 95, 103657. [Google Scholar] [CrossRef]
- Daly-Hassen, H.; Annabi, M.; King-Okumu, C. Social and Private Profitability of Tree-Based Adaptation Options to Climate Change in a Dryland Area of Tunisia. New Medit 2019, 18, 89–105. [Google Scholar] [CrossRef]
- Chtourou, F.; Valli, E.; Ben Mansour, A.; Bendini, A.; Gallina Toschi, T.; Bouaziz, M. Characterization of Virgin Olive Oils Obtained from Minor Tunisian Varieties for Their Valorization. J. Food Meas. Charact. 2021, 15, 5060–5070. [Google Scholar] [CrossRef]
- Omri, A.; Abdelhamid, S.; Benincasa, C.; Araouki, A.; Ayadi, M.; Gharsallaoui, M.; Gouiaa, M. Genetic Diversity and Association of Molecular Markers with Biochemical Traits in Tunisian Olive Cultivars. Genet. Resour. Crop Evol. 2021, 68, 1181–1197. [Google Scholar] [CrossRef]
- Debbabi, O.S.; Ben Amar, F.; Rahmani, S.M.; Taranto, F.; Montemurro, C.; Miazzi, M.M. The Status of Genetic Resources and Olive Breeding in Tunisia. Plants 2022, 11, 1759. [Google Scholar] [CrossRef] [PubMed]
- Moriondo, M.; Ferrise, R.; Trombi, G.; Brilli, L.; Dibari, C.; Bindi, M. Modelling Olive Trees and Grapevines in a Changing Climate. Environ. Model. Softw. 2015, 72, 387–401. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2021, 11, 56. [Google Scholar] [CrossRef]
- Avolio, E.; Orlandi, F.; Bellecci, C.; Fornaciari, M.; Federico, S. Assessment of the Impact of Climate Change on the Olive Flowering in Calabria (Southern Italy). Theor. Appl. Climatol. 2012, 107, 531–540. [Google Scholar] [CrossRef]
- Boussadia, O.; Ben Mariem, F.; Mechri, B.; Boussetta, W.; Braham, M.; Hadj, S.B. Response to Drought of Two Olive Tree Cultivars (Cv Koroneki and Meski). Sci. Hortic. 2008, 116, 388–393. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H. Physiological and Anatomical Changes Induced by Drought in Two Olive Cultivars (Cv Zalmati and Chemlali). Acta Physiol. Plant. 2011, 33, 53–65. [Google Scholar] [CrossRef]
- Gholami, R.; Fahadi Hoveizeh, N.; Zahedi, S.M.; Gholami, H.; Carillo, P. Effect of Three Water-Regimes on Morpho-Physiological, Biochemical and Yield Responses of Local and Foreign Olive Cultivars under Field Conditions. BMC Plant Biol. 2022, 22, 477. [Google Scholar] [CrossRef]
- Guyot, G.; Scoffoni, C.; Sack, L. Combined Impacts of Irradiance and Dehydration on Leaf Hydraulic Conductance: Insights into Vulnerability and Stomatal Control. Plant Cell Environ. 2012, 35, 857–871. [Google Scholar] [CrossRef]
- Sack, L.; Cowan, P.; Jaikumar, N.; Holbrook, N. The Hydrology of Leaves Co-ordination of Structure and Function in Temperate Woody Species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; Rahemi, M.; Rostami, A.A.; Sedaghat, S. Drought Effects on Growth, Water Content and Osmoprotectants in Four Olive Cultivars with Different Drought Tolerance. Int. J. Fruit Sci. 2018, 18, 254–267. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought Stress Effects and Olive Tree Acclimation under a Changing Climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Santana, V.; Rodriguez-Dominguez, C.M.; Fernández, J.E.; Diaz-Espejo, A. Role of Leaf Hydraulic Conductance in the Regulation of Stomatal Conductance in Almond and Olive in Response to Water Stress. Tree Physiol. 2016, 36, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.M.; Woodruff, D.R.; McCulloh, K.A.; Meinzer, F.C. Leaf Hydraulic Conductance, Measured in Situ, Declines and Recovers Daily: Leaf Hydraulics, Water Potential and Stomatal Conductance in Four Temperate and Three Tropical Tree Species. Tree Physiol. 2009, 29, 879–887. [Google Scholar] [CrossRef]
- Tyree, M.T.; Nardini, A.; Salleo, S.; Sack, L.; El Omari, B. The Dependence of Leaf Hydraulic Conductance on Irradiance during HPFM Measurements: Any Role for Stomatal Response? J. Exp. Bot. 2005, 56, 737–744. [Google Scholar] [CrossRef]
- Flexas, J.; Scoffoni, C.; Gago, J.; Sack, L. Leaf Mesophyll Conductance and Leaf Hydraulic Conductance: An Introduction to Their Measurement and Coordination. J. Exp. Bot. 2013, 64, 3965–3981. [Google Scholar] [CrossRef]
- Bosabalidis, A.M.; Kofidis, G. Comparative Effects of Drought Stress on Leaf Anatomy of Two Olive Cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- Ahumada-Orellana, L.; Ortega-Farías, S.; Poblete-Echeverría, C.; Searles, P.S. Estimation of Stomatal Conductance and Stem Water Potential Threshold Values for Water Stress in Olive Trees (Cv. Arbequina). Irrig. Sci. 2019, 37, 461–467. [Google Scholar] [CrossRef]
- Boussadia, O.; Ben Hassine, M.; Braham, M. Effect of Water Stress on Photosynthetic Assimilation and Biomass Accumulation in Olive Tree. Acta Sci. Agric. 2018, 2, 84–92. [Google Scholar]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H. Drought Stress in Plants: An Overview. In Plant Responses to Drought Stress. From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; p. 466. ISBN 978-3-642-32653-0. [Google Scholar]
- Lawlor, D.W. Limitation to Photosynthesis in Water-Stressed Leaves: Stomata vs. Metabolism and the Role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Cornic, G. Photosynthetic Carbon Assimilation and Associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes-Silva, A.A.; López-Bernal, Á.; Ferreira, T.C.; Villalobos, F.J. Leaf Water Relations and Gas Exchange Response to Water Deficit of Olive (Cv. Cobrançosa) in Field Grown Conditions in Portugal. Plant Soil 2016, 402, 191–209. [Google Scholar] [CrossRef]
- Edziri, H.; Chehab, H.; Aissaoui, F.; Boujnah, D.; Mastouri, M. Photosynthetic, Anatomical and Biochemical Responses of Olive Tree (Olea europaea) Cultivars under Water Stress. Plant Biosyst. 2021, 155, 740–746. [Google Scholar] [CrossRef]
- Zushi, K.; Matsuzoe, N. Using of Chlorophyll a Fluorescence OJIP Transients for Sensing Salt Stress in the Leaves and Fruits of Tomato. Sci. Hortic. 2017, 219, 216–221. [Google Scholar] [CrossRef]
- Lepeduš, H.; Brkić, I.; Cesar, V.; Jurković, V.; Antunović, J.; Jambrović, A.; Brkić, J.; Šimić, D. Chlorophyll Fluorescence Analysis of Photosynthetic Performance in Seven Maize Inbred Lines under Water-Limited Conditions. Period. Biol. 2012, 114, 73–76. [Google Scholar]
- Živčák, M.; Brestič, M.; Olšovská, K.; Slámka, P. Performance Index as a Sensitive Indicator of Water Stress in Triticum aestivum L. Plant Soil Environ. 2008, 54, 133–139. [Google Scholar] [CrossRef] [Green Version]
- van Heerden, P.D.R.; Swanepoel, J.W.; Krüger, G.H.J. Modulation of Photosynthesis by Drought in Two Desert Scrub Species Exhibiting C3-Mode CO2 Assimilation. Environ. Exp. Bot. 2007, 61, 124–136. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesi; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transiet as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: London, UK, 2000; pp. 445–483. ISBN 0748408215. [Google Scholar]
- Moula, I.; Boussadia, O.; Koubouris, G.; Ben Hassine, M.; Boussetta, W.; Van Labeke, M.C.; Braham, M. Ecophysiological and Biochemical Aspects of Olive Tree (Olea europaea L.) in Response to Salt Stress and Gibberellic Acid-Induced Alleviation. S. Afr. J. Bot. 2020, 132, 38–44. [Google Scholar] [CrossRef]
- Faraloni, C.; Cutino, I.; Petruccelli, R.; Leva, A.R.; Lazzeri, S.; Torzillo, G. Chlorophyll Fluorescence Technique as a Rapid Tool for in Vitro Screening of Olive Cultivars (Olea europaea L.) Tolerant to Drought Stress. Environ. Exp. Bot. 2011, 73, 49–56. [Google Scholar] [CrossRef]
- Torres, I.; Sánchez, M.T.; Benlloch-González, M.; Pérez-Marín, D. Irrigation Decision Support Based on Leaf Relative Water Content Determination in Olive Grove Using near Infrared Spectroscopy. Biosyst. Eng. 2019, 180, 50–58. [Google Scholar] [CrossRef]
- El Refaey, A.A.; Mohamed, Y.I.; El-Shazly, S.M.; Abd El Salam, A.A. Effect of Salicylic and Ascorbic Acids Foliar Application on Picual Olive Trees Growth under Water Stress Condition. Egypt. J. Soil Sci. 2022, 62, 1–17. [Google Scholar] [CrossRef]
- Dias, M.C.; Correia, S.; Serôdio, J.; Silva, A.M.S.; Freitas, H.; Santos, C. Chlorophyll Fluorescence and Oxidative Stress Endpoints to Discriminate Olive Cultivars Tolerance to Drought and Heat Episodes. Sci. Hortic. 2018, 231, 31–35. [Google Scholar] [CrossRef]
- Sofo, A.; Manfreda, S.; Fiorentino, M.; Dichio, B.; Xiloyannis, C. The Olive Tree: A Paradigm for Drought Tolerance in Mediterranean Climates. Hydrol. Earth Syst. Sci. Discuss. 2007, 4, 2811–2835. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; Yadollahi, A.; Arzani, K.; Imani, A.; Aghaalikhani, M. Gas-Exchange Response of Almond Genotypes to Water Stress. Photosynthetica 2015, 53, 29–34. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Ferreira, H.F.; Correia, C.M. Immediate Responses and Adaptative Strategies of Three Olive Cultivars under Contrasting Water Availability Regimes: Changes on Structure and Chemical Composition of Foliage and Oxidative Damage. Plant Sci. 2006, 170, 596–605. [Google Scholar] [CrossRef]
- Vasques, A.R.; Pinto, G.; Dias, M.C.; Correia, C.M.; Moutinho-Pereira, J.M.; Vallejo, V.R.; Santos, C.; Keizer, J.J. Physiological Response to Drought in Seedlings of Pistacia Lentiscus (Mastic Tree). New For. 2016, 47, 119–130. [Google Scholar] [CrossRef]
- Boussadia, O.; Steppe, K.; Zgallai, H.; Ben El Hadj, S.; Braham, M.; Lemeur, R.; Van Labeke, M.C. Nondestructive Determination of Nitrogen and Chlorophyll Content in Olive Tree Leaves and the Relation with Photosynthesis and Fluorescence Parameters. Photosynthetica 2011, 49, 149–153. [Google Scholar] [CrossRef]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-Oxidative Stress Markers as a Measure of Abiotic Stress-Induced Leaf Senescence: Advantages and Limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Srivastava, A. Govindjee Polyphasic Chlorophyll a Fluorescence Transient in plants and Cyanobacteria. Am. Soc. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivar | T | RWC (%) | SR (s·cm−1) | FV/FM | PI | SPAD Index |
|---|---|---|---|---|---|---|
| Besbessi | T0 | 92.67 ± 1.31 A,a | 1.39 ± 0.21 B,d | 0.828 ± 0.003 CD,a | 12.27 ± 0.41 A,a | 90.41 ± 1.58 A,a |
| T15 | 92.84 ± 1.27 A,a | 2.03 ± 0.34 B,d | 0.828 ± 0.004 A,a | 11.68 ± 0.65 A,a | 88.63 ± 2.80 A,ab | |
| T30 | 88.00 ± 0.99 A,b | 3.76 ± 0.46 B,c | 0.827 ± 0.006 AB,a | 8.39 ± 0.68 B,b | 84.03 ± 1.89 B,bc | |
| T45 | 78.99 ± 1.66 A,c | 6.86 ± 0.52 B,b | 0.801 ± 0.011 A,b | 5.20 ± 0.65 BC,c | 84.96 ± 1.22 A,abc | |
| T60 | 55.00 ± 1.20 A,d | 13.73 ± 0.71 A,a | 0.790 ± 0.01 B,b | 3.72 ± 0.04 D,d | 82.39 ± 1.30 AB,c | |
| Chemchali | T0 | 92.17 ± 0.33 AB,a | 1.37 ± 0.15 B,d | 0.834 ± 0.004 BC,a | 12.54 ± 0.67 A,a | 89.34 ± 2.30 A,ab |
| T15 | 90.15 ± 0.56 AB,a | 2.03 ± 0.59 B,d | 0.832 ± 0.007 A,a | 11.33 ± 0.24 A,ab | 90.39 ± 3.09 A,a | |
| T30 | 79.95 ± 2.81 B,b | 44.9 ± 0.47 AB,c | 0.830 ± 0.0006 AB,a | 10.58 ± 0.31 A,b | 89.13 ± 2.20 A,ab | |
| T45 | 73.33 ± 3.21 A,b | 7.75 ± 0.65 AB,b | 0.817 ± 0.005 A,b | 8.056 ± 0.92 A,c | 86.41 ± 2.09 A,ab | |
| T60 | 54.99 ± 6.61 A,c | 11.69 ± 0.46 B,a | 0.830 ± 0.0006 A,a | 6.60 ± 0.44 A,c | 82.97 ± 2.12 AB,b | |
| Chetoui | T0 | 90.14 ± 0.22 B,a | 1.25 ± 0.14 B,d | 0.821 ± 0.002 D,a | 12.52 ± 0.23 A,a | 89.38 ± 1.74 A,a |
| T15 | 87.91 ± 1.06 B,a | 3.25 ± 0.41 AB,c | 0.825 ± 0.002 A,a | 10.89 ± 0.44 AB,b | 89.47 ± 2.54 A,a | |
| T30 | 72.77 ± 2.25 C,b | 6,00 ± 0.87 A,b | 0.817 ± 0.010 B,ab | 8.56 ± 0.58 B,c | 84.18 ± 1.62 B,ab | |
| T45 | 60.70 ± 2.46 C,c | 8.42 ± 0.32 A,a | 0.799 ± 0.002 A,b | 5.40 ± 0.16 B,d | 79.45 ± 1.22 B,b | |
| T60 | 49.71 ± 1.33 A,d | 9.66 ± 0.42 C,a | 0.722 ± 0.012 D,c | 1.57 ± 0.03 C,e | 79.24 ± 2.58 B,b | |
| Jarboui | T0 | 91.49 ± 0.77 AB,a | 2.00 ± 0.14 A,c | 0.841 ± 0.004 B,a | 11.66 ± 0.45 a,a | 89.11 ± 1.88 A,a |
| T15 | 92.11 ± 2.00 A,a | 4.48 ± 0.66 A,b | 0.831 ± 0.004 A,a | 10.21 ± 0.18 b,b | 87.18 ± 1.22 A,a | |
| T30 | 77.27 ± 0.61 BC,b | 5.94 ± 0.83 A,a | 0.836 ± 0.014 AB,a | 9.13 ± 0.23 b,c | 86.87 ± 0.23 AB,a | |
| T45 | 55.00 ± 1.70 C,c | 6.81 ± 0.20 B,a | 0.800 ± 0.007 A,b | 3.68 ± 0.06 c,d | 85.43 ± 2.43 A,ab | |
| T60 | 37.00 ± 1.00 B,d | 7.10 ± 0.40 D,a | 0.745 ± 0.005 C,c | 1.51 ± 0.33 c,e | 81.07 ± 1.89 AB,b | |
| Sayali | T0 | 92.50 ± 0.68 A,a | 1.56 ± 0.13 B,d | 0.851 ± 0.001 A,a | 12.30 ± 0.128 A,a | 91.14 ± 1.57 A,a |
| T15 | 92.02 ± 2.11 A,a | 2.46 ± 0.30 B,d | 0.839 ± 0.010 A,ab | 10.99 ± 0.19 AB,b | 88.52 ± 0.92 A,a | |
| T30 | 86.49 ± 2.80 A,b | 3.53 ± 0.26 B,c | 0.842 ± 0.005 A,a | 10.46 ± 0.15 A,b | 90.10 ± 1.10 A,a | |
| T45 | 67.30 ± 0.64 B,c | 5.00 ± 0.55 C,b | 0.795 ± 0.021 A,c | 4.92 ± 0.67 BC,c | 88.60 ± 0.436 A,a | |
| T60 | 53.00 ± 1.090 A,d | 8.03 ± 0.39 D,a | 0.811 ± 0.002 A,bc | 3.14 ± 0.52 B,d | 85.03 ± 0.97 A,b | |
| p-value | Cultivar | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Cultivar × Treatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.009 |
| Cultivar | Treatment | |
|---|---|---|
| T0 | T60 | |
| Besbessi | 139.69 ± 19.21 D,a | 27,91 ± 0.38 C,b |
| Chemchali | 177.01 ± 8.54 B,a | 31.075 ± 0.79 B,b |
| Chetoui | 149.13 ± 0.85 CD,a | 27.03 ± 1.98 C,b |
| Jarboui | 251.78 ± 2.47 A,a | 37.130 ± 0.36 A,b |
| Sayali | 174.80 ± 15.81 BC,a | 32.11 ± 0.43 B,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boussadia, O.; Omri, A.; Mzid, N. Eco-Physiological Behavior of Five Tunisian Olive Tree Cultivars under Drought Stress. Agronomy 2023, 13, 720. https://doi.org/10.3390/agronomy13030720
Boussadia O, Omri A, Mzid N. Eco-Physiological Behavior of Five Tunisian Olive Tree Cultivars under Drought Stress. Agronomy. 2023; 13(3):720. https://doi.org/10.3390/agronomy13030720
Chicago/Turabian StyleBoussadia, Olfa, Amal Omri, and Nada Mzid. 2023. "Eco-Physiological Behavior of Five Tunisian Olive Tree Cultivars under Drought Stress" Agronomy 13, no. 3: 720. https://doi.org/10.3390/agronomy13030720