

Comparative Transcriptome Analysis of the Differential Effects of Florpyrauxifen-Benzyl Treatment on Phytohormone Transduction between Florpyrauxifen-Benzyl-Resistant and -Susceptible Barnyard Grasses (Echinochloa crus-galli (L.) P. Beauv)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.1.1. Plant Material Description
2.1.2. Dose–Response Experiments of Florpyrauxifen-Benzyl
2.1.3. Seedling Preparation for Transcriptome Analysis
2.2. Statistical Treatment of Dose–Response Curves
2.3. RNA Isolation and Transcriptome Sequencing
2.4. Transcriptome Assembly and Functional Classification
2.5. Differential Gene Expression and Enrichment Analysis
2.6. RT–qPCR Validation
3. Results
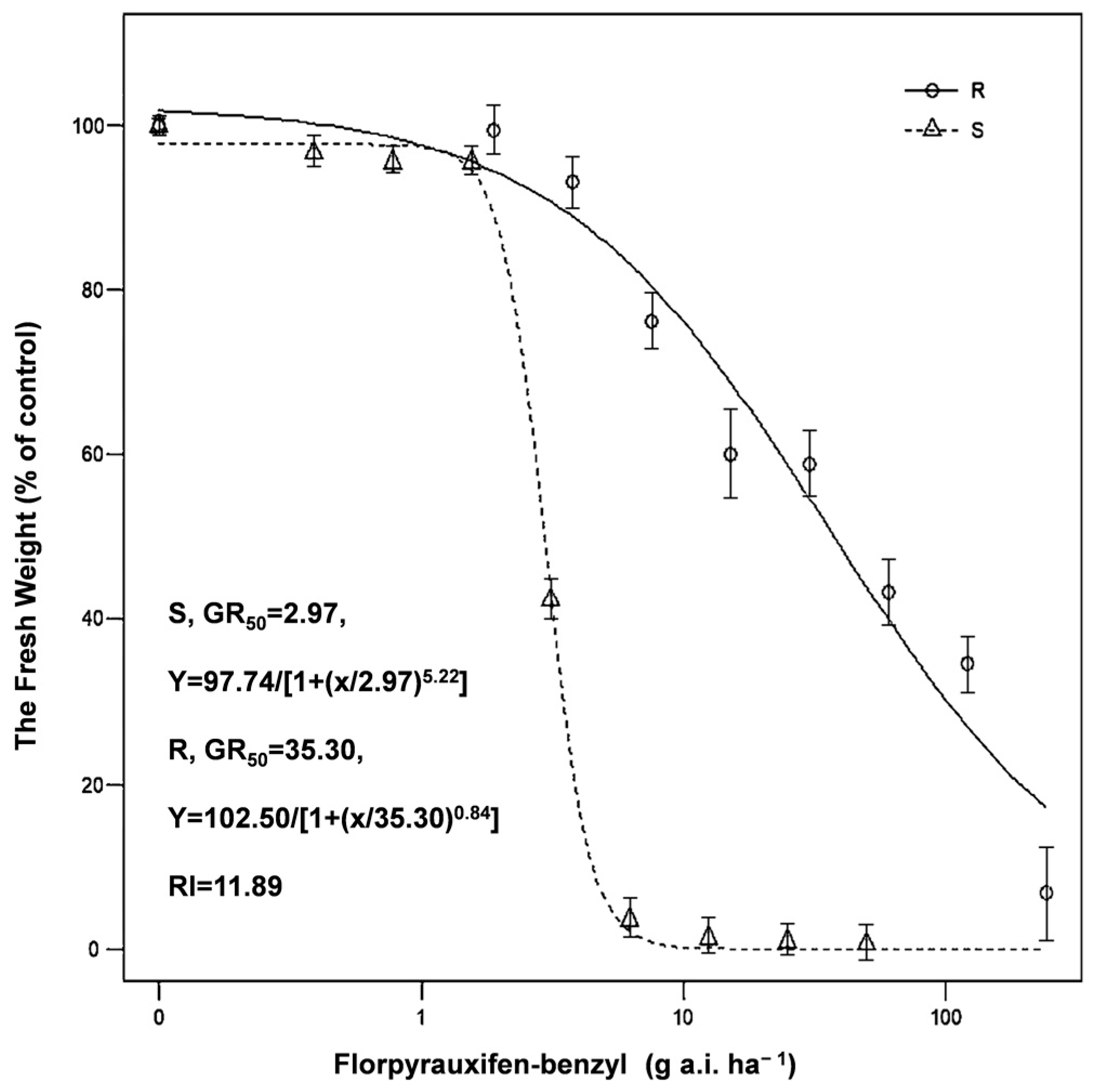
3.1. Florpyrauxifen-Benzyl Efficacy against Resistant and Susceptible E. crus-galli
3.2. De Novo Assembly of E. crus-galli Reference Transcriptome
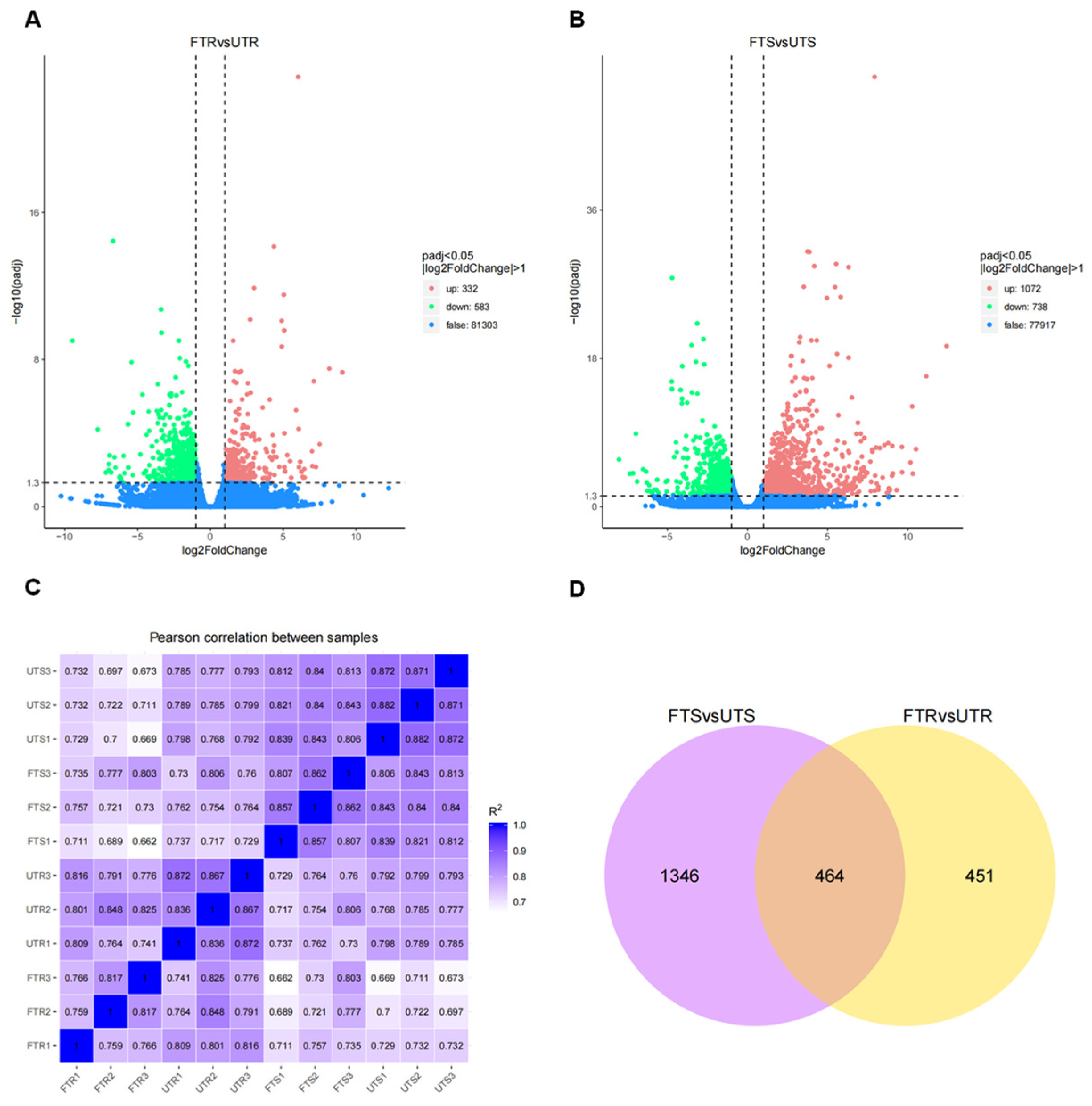
3.3. Identification of Differentially Expressed Genes
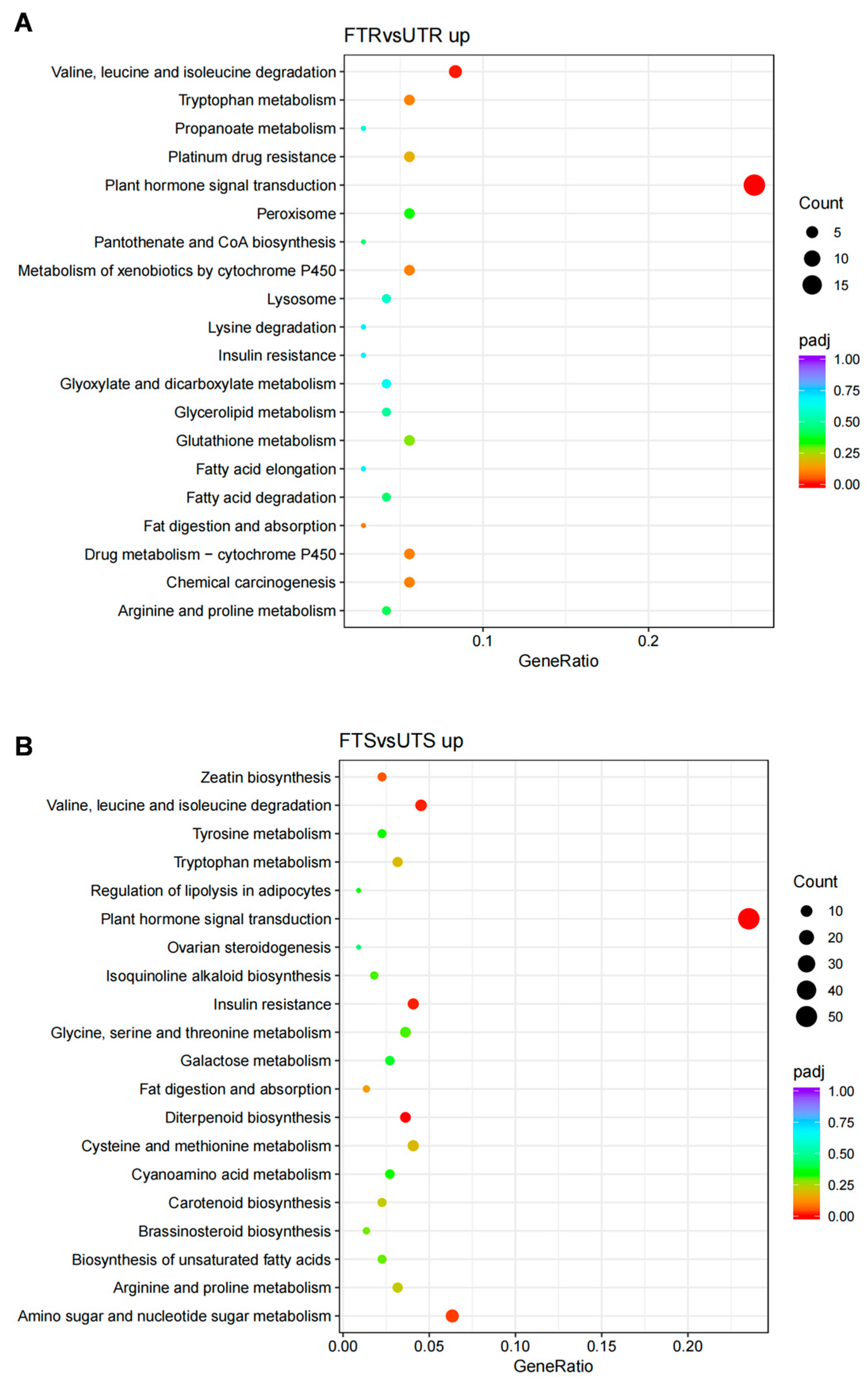
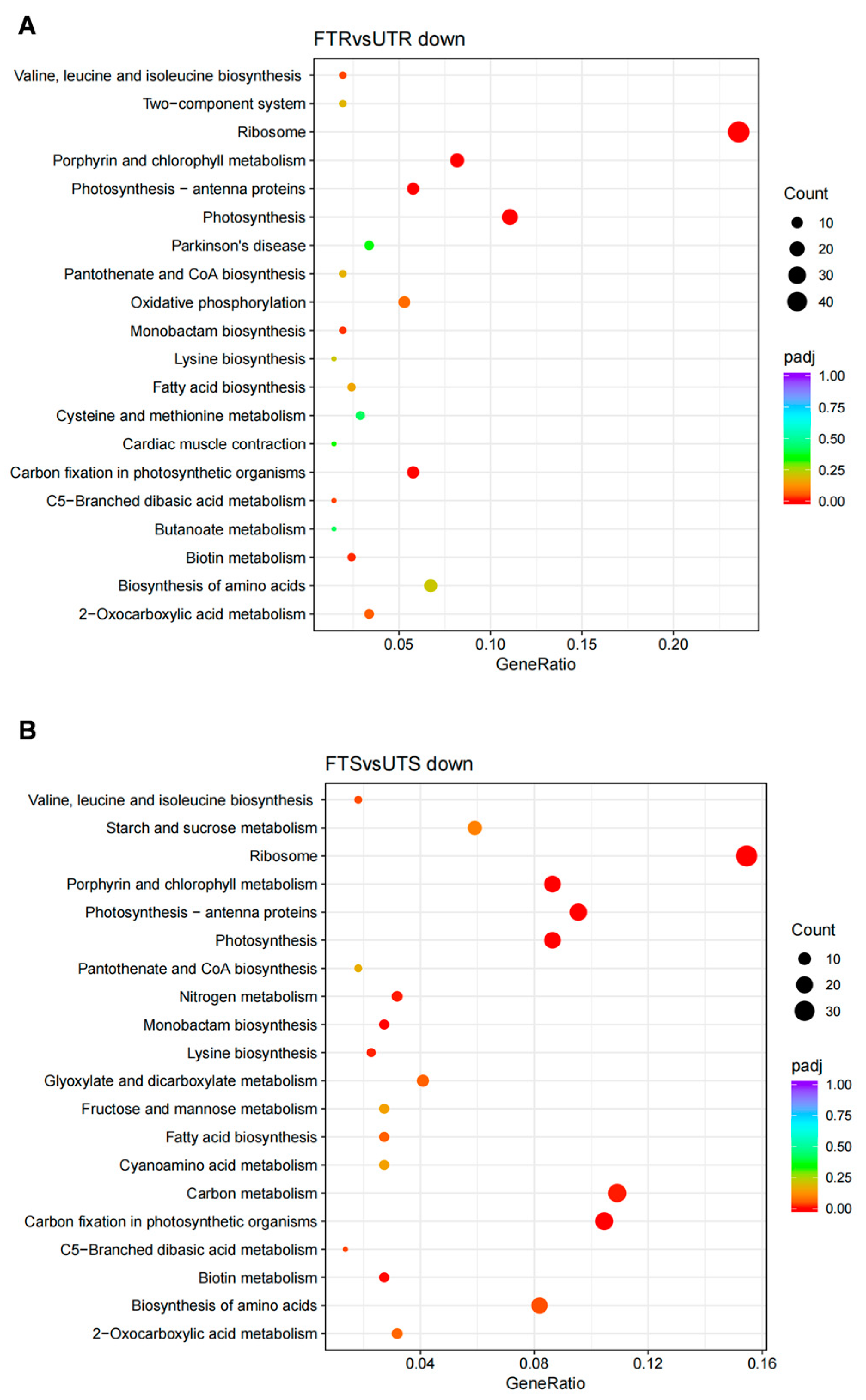
3.4. Enriched GO Terms and KEGG Pathways of DEGs
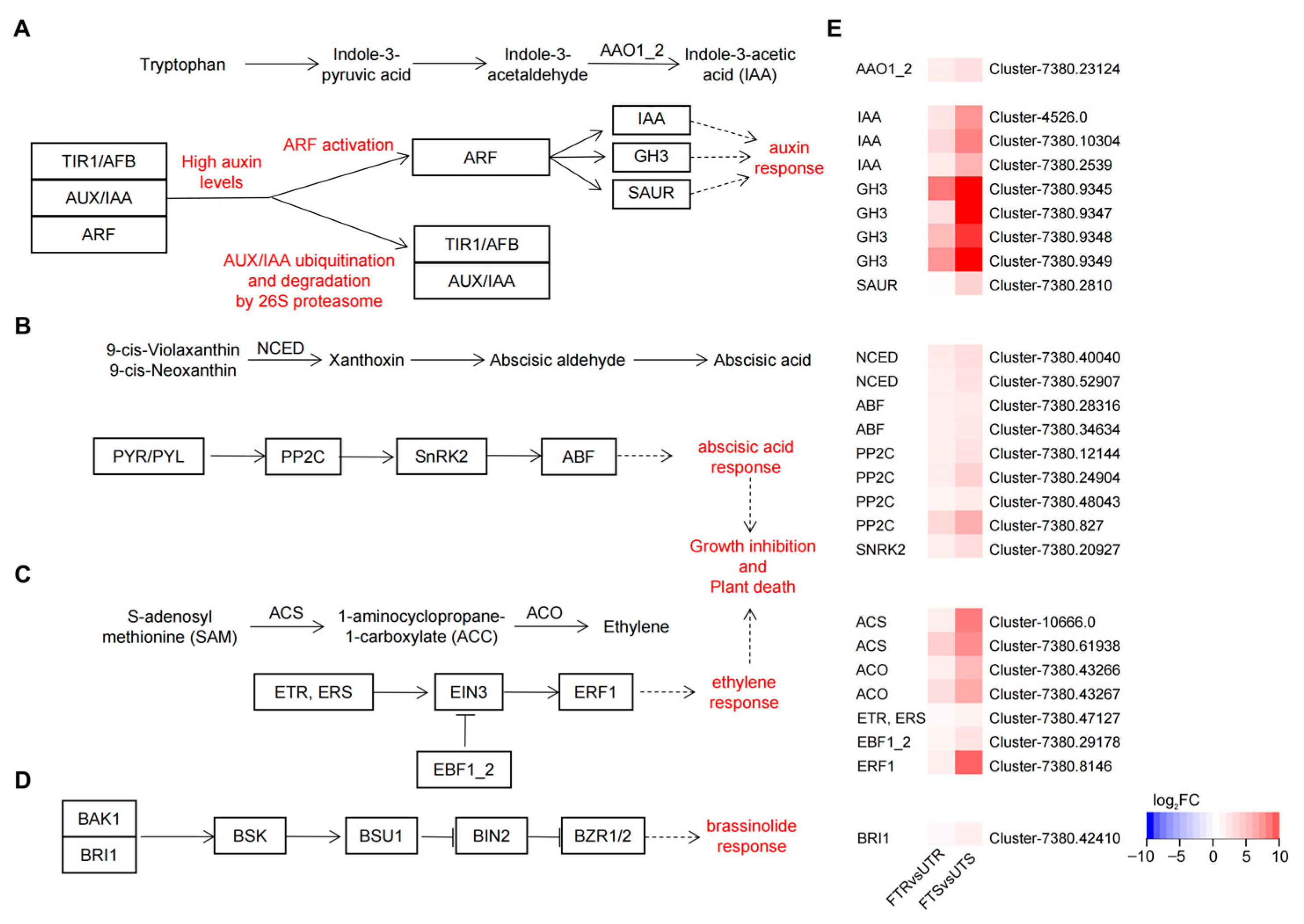
3.5. DEGs Related to Phytohormone Biosynthesis and Signal Transduction
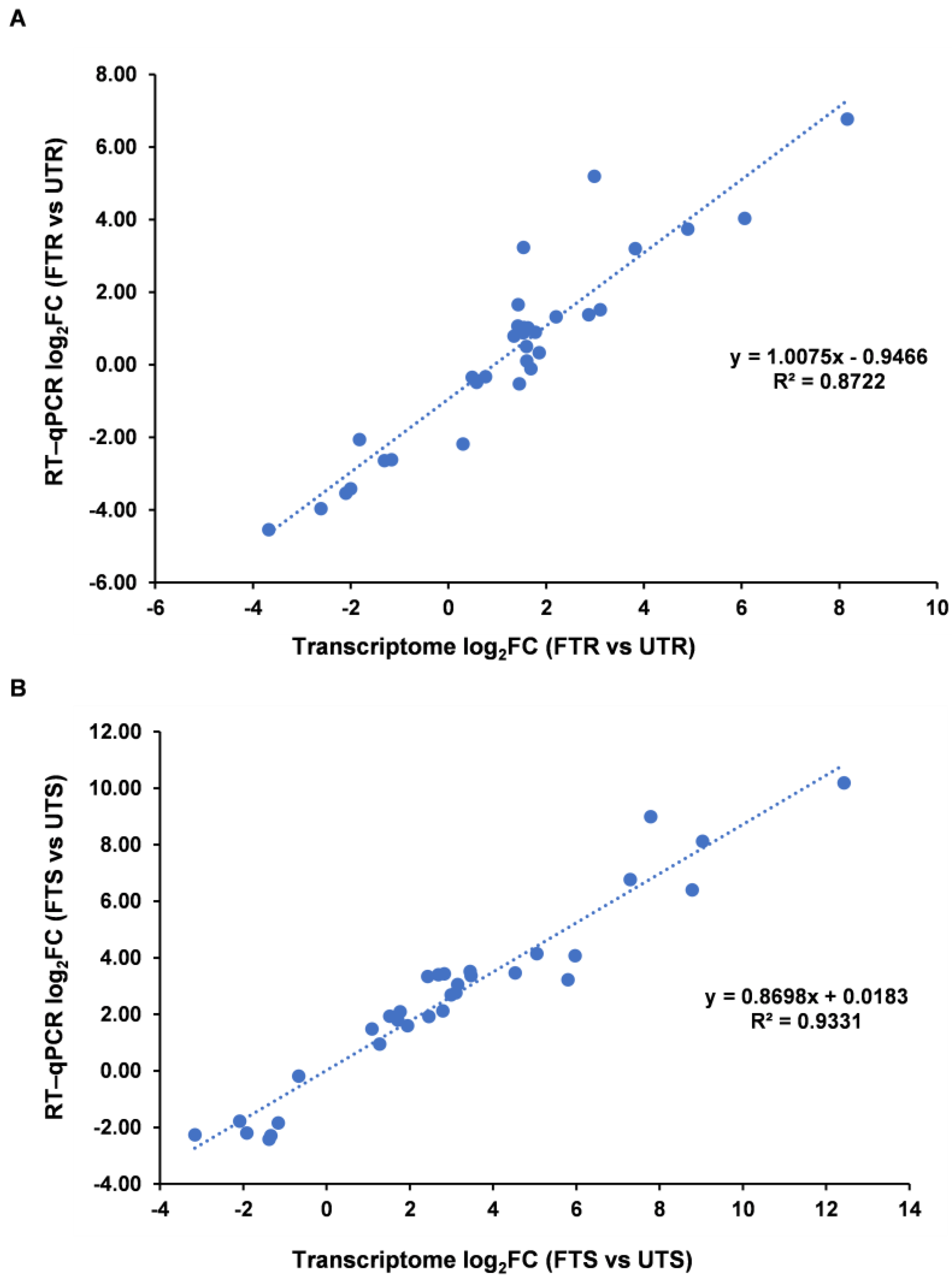
3.6. Validation of DEGs by Real-Time Quantitative PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bryson, C.T.; DeFelice, M.S. Weeds of the South; University of Georgia Press: Athens, GA, USA, 2009; pp. 373–374. [Google Scholar]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: http://weedscience.org/Pages/Species.aspx (accessed on 5 December 2022).
- Grossmann, K. Mode of action of auxin herbicides: A new ending to a long, drawn out story. Trends Plant Sci. 2000, 5, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Huang, Y.; Qi, P.; Lian, G.; Hu, X.; Han, N.; Wang, J.; Zhu, M.; Qian, Q.; Bian, H. Functional analysis of auxin receptor OsTIR1/OsAFB family members in rice grain yield, tillering, plant height, root system, germination, and auxinic herbicide resistance. New Phytol. 2021, 229, 2676–2692. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, S.B.; Wang, X.J.; Hagen, G.; Guilfoyle, T.J. AUX/IAA proteins are active repressors, and their stability and activity are modulated by auxin. Plant Cell 2001, 13, 2809–2822. [Google Scholar] [CrossRef] [Green Version]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef]
- Velásquez, J.C.; Bundt, A.D.C.; Camargo, E.R.; Andres, A.; Viana, V.E.; Hoyos, V.; Plaza, G.; de Avila, L.A. Florpyrauxifen-Benzyl Selectivity to Rice. Agriculture 2021, 11, 1270. [Google Scholar] [CrossRef]
- Lavy, M.; Estelle, M. Mechanisms of auxin signaling. Development 2016, 143, 3226–3229. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.A.; Neal, R.; Merlo, A.O.; Honma, M.; Hicks, G.R.; Wolff, K.; Matsumura, W.; Davies, J.P. Mutations in an auxin receptor homolog afb5 and in sgt1b confer resistance to synthetic picolinate auxins and not to 2,4-dichlorophenoxyacetic acid or indole-3-acetic acid in arabidopsis. Plant Physiol. 2006, 142, 542–552. [Google Scholar] [CrossRef] [Green Version]
- Prigge, M.J.; Platre, M.; Kadakia, N.; Zhang, Y.; Greenham, K.; Szutu, W.; Pandey, B.K.; Bhosale, R.A.; Bennett, M.J.; Busch, W.; et al. Genetic analysis of the Arabidopsis TIR1/AFB auxin receptors reveals both overlapping and specialized functions. eLife 2020, 9, e54740. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Weijers, D.; Lechner, E.; Yamada, M.; Hobbie, L.; Ehrismann, J.S.; Jürgens, G.; Estelle, M. Plant development is regulated by a family of auxin receptor F box proteins. Dev. Cell 2005, 9, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Sundaram, S.; Armitage, L.; Evans, J.P.; Hawkes, T.; Kepinski, S.; Ferro, N.; Napier, R.M. Defining binding efficiency and specificity of auxins for SCF(TIR1/AFB)-Aux/IAA co-receptor complex formation. ACS Chem. Biol. 2014, 9, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Greenham, K.; Zhang, Y.; Santner, A.; Castillejo, C.; Mutka, A.M.; O’Malley, R.C.; Ecker, J.R.; Kunkel, B.N.; Estelle, M. The Arabidopsis auxin receptor F-box proteins AFB4 and AFB5 are required for response to the synthetic auxin picloram. G3 Genes Genomes Genet. 2016, 6, 1383–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, J.; Schmitzer, P.; Weimer, M.; Napier, R.; Prusinska, J. Mode-of-action analysis of a new arylpicolinate herbicide from Dow AgroSciences. In Proceedings of the Weed Science Society of America Annual Meeting; Weed Science Society of America: Lexington, KY, USA, 2015. [Google Scholar]
- Miller, M.R.; Norsworthy, J.K.; Scott, R.C. Evaluation of florpyrauxifen-benzyl on herbicide-resistant and herbicide-susceptible barnyardgrass accessions. Weed Technol. 2018, 32, 126–134. [Google Scholar] [CrossRef]
- Lim, S.H.; Kim, H.; Noh, T.K.; Lim, J.S.; Yook, M.J.; Kim, J.W.; Yi, J.H.; Kim, D.S. Baseline Sensitivity of Echinochloa crus-gall and E. oryzicola to Florpyrauxifen-Benzyl, a New Synthetic Auxin Herbicide, in Korea. Front. Plant Sci. 2021, 12, 656642. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.E.; Norsworthy, J.K.; Roberts, T.L.; Scott, R.; Hardke, J.; Gbur, E.E. Rice Cultivar Response to Florpyrauxifen-Benzyl When Applied with Imazethapyr and a Cytochrome P450 Inhibitor. Int. J. Agron. 2020, 2020, 2769610. [Google Scholar] [CrossRef]
- Wang, H.; Sun, X.; Yu, J.; Li, J.; Dong, L. The phytotoxicity mechanism of florpyrauxifen-benzyl to Echinochloa crus-galli (L.) P. Beauv and weed control effect. Pestic. Biochem. Physiol. 2021, 179, 104978. [Google Scholar] [CrossRef]
- Miller, M.R.; Norsworthy, J.K. Influence of Soil Moisture on Absorption, Translocation, and Metabolism of Florpyrauxifen-benzyl. Weed Sci. 2018, 66, 418–423. [Google Scholar] [CrossRef]
- Epp, J.B.; Alexander, A.L.; Balko, T.W.; Buysse, A.M.; Brewster, W.K.; Bryan, K.; Daeuble, J.F.; Fields, S.C.; Gast, R.E.; Green, R.A.; et al. The discovery of Arylex™ active and Rinskor™ active: Two novel auxin herbicides. Bioorg. Med. Chem. 2016, 24, 362–371. [Google Scholar] [CrossRef]
- Hwang, J.I.; Norsworthy, J.K.; González-Torralva, F.; Priess, G.L.; Barber, L.T.; Butts, T.R. Non-target-site resistance mechanism of barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.] to florpyrauxifen-benzyl. Pest Manag. Sci. 2022, 78, 287–295. [Google Scholar] [CrossRef]
- Hwang, J.I.; Norsworthy, J.K.; González-Torralva, F.; Piveta, L.B.; Priess, G.L.; Barber, L.T.; Butts, T.R. Absorption, translocation, and metabolism of florpyrauxifen-benzyl and cyhalofop-butyl in cyhalofop-butyl-resistant barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.]. Pestic. Biochem. Physiol. 2022, 180, 104999. [Google Scholar] [CrossRef]
- Giacomini, D.A.; Gaines, T.; Beffa, R.; Tranel, P.J. Optimizing RNA-seq studies to investigate herbicide resistance. Pest Manag. Sci. 2018, 74, 2260–2264. [Google Scholar] [CrossRef]
- Wright, A.A.; Sasidharan, R.; Koski, L.; Rodriguez-Carres, M.; Peterson, D.G.; Nandula, V.K.; Ray, J.D.; Bond, J.A.; Shaw, D.R. Transcriptomic changes in Echinochloa colona in response to treatment with the herbicide imazamox. Planta 2018, 247, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Romero-Puertas, M.C.; Peláez-Vico, M.Á.; Pazmiño, D.M.; Rodríguez-Serrano, M.; Terrón-Camero, L.; Bautista, R.; Gómez-Cadenas, A.; Claros, M.G.; León, J.; Sandalio, L.M. Insights into ROS-dependent signalling underlying transcriptomic plant responses to the herbicide 2,4-D. Plant Cell Environ. 2022, 45, 572–590. [Google Scholar] [CrossRef] [PubMed]
- Rangani, G.; Rouse, C.E.; Saski, C.; Noorai, R.E.; Shankar, V.; Lawton-Rauh, A.L.; Werle, I.S.; Roma-Burgos, N. High Resistance to Quinclorac in Multiple-Resistant Echinochloa colona Associated with Elevated Stress Tolerance Gene Expression and Enriched Xenobiotic Detoxification Pathway. Genes 2022, 13, 515. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinf. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Jiao, C.; Sun, H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.; Banf, M.; Dai, X.; Martin, G.B.; Giovannoni, J.J.; et al. iTAK: A program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Y.; Li, J.; Fang, J.; Liu, T.; Dong, L. Transcriptome profiling to identify cytochrome P450 genes involved in penoxsulam resistance in Echinochloa glabrescens. Pestic. Biochem. Physiol. 2019, 158, 112–120. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Koga, J.; Adachi, T.; Hidaka, H. Purification and characterization of indolepyruvate decarboxylase. A novel enzyme for indole-3-acetic acid biosynthesis in Enterobacter cloacae. J. Biol. Chem. 1992, 267, 15823–15828. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Akaba, S.; Oritani, T.; Delarue, M.; Bellini, C.; Caboche, M.; Koshiba, T. Higher activity of an aldehyde oxidase in the auxin-overproducing superroot1 mutant of Arabidopsis thaliana. Plant Physiol. 1998, 116, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Woeste, K.E.; Vogel, J.P.; Kieber, J.J. Factors regulating ethylene biosynthesis in etiolated Arabidopsis thaliana seedlings. Physiol. Plant. 1999, 105, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Abel, S.; Nguyen, M.D.; Chow, W.; Theologis, A. ACS4, a primary indoleacetic acid-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis thaliana. Structural characterization, expression in Escherichia coli, and expression characteristics in response to auxin. J. Biol. Chem. 1995, 270, 19093–19099. [Google Scholar] [CrossRef] [Green Version]
- Yamagami, T.; Tsuchisaka, A.; Yamada, K.; Haddon, W.F.; Harden, L.A.; Theologis, A. Biochemical diversity among the 1-amino-cyclopropane-1-carboxylate synthase isozymes encoded by the Arabidopsis gene family. J. Biol. Chem. 2003, 278, 49102–49112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraft, M.; Kuglitsch, R.; Kwiatkowski, J.; Frank, M.; Grossmann, K. Indole-3-acetic acid and auxin herbicides up-regulate 9-cis-epoxycarotenoid dioxygenase gene expression and abscisic acid accumulation in cleavers (Galium aparine): Interaction with ethylene. J. Exp. Bot. 2007, 58, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Gleason, C.; Foley, R.C.; Singh, K.B. Mutant analysis in Arabidopsis provides insight into the molecular mode of action of the auxinic herbicide dicamba. PLoS ONE 2011, 6, e17245. [Google Scholar] [CrossRef] [Green Version]
- Pettinga, D.J.; Ou, J.; Patterson, E.L.; Jugulam, M.; Westra, P.; Gaines, T.A. Increased chalcone synthase (CHS) expression is associated with dicamba resistance in Kochia scoparia. Pest Manag. Sci. 2018, 74, 2306–2315. [Google Scholar] [CrossRef]
- Grossmann, K.; Kwiatkowski, J.; Tresch, S. Auxin herbicides induce H2O2 overproduction and tissue damage in cleavers (Galium aparine L.). J. Exp. Bot. 2001, 52, 1811–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Zeevaart, J.A. The 9-cis-epoxycarotenoid cleavage reaction is the key regulatory step of abscisic acid biosynthesis in water-stressed bean. Proc. Natl. Acad. Sci. USA 1999, 96, 15354–15361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemhauser, J.L.; Mockler, T.C.; Chory, J. Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol. 2004, 2, E258. [Google Scholar] [CrossRef]
- Kieffer, M.; Neve, J.; Kepinski, S. Defining auxin response contexts in plant development. Curr. Opin. Plant Biol. 2010, 13, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, S.U.; Shin, D.M.; Pham, G.; Jeong, Y.S.; Kim, S.H. ATBS1-INTERACTING FACTOR 2 negatively regulates dark- and brassinosteroid-induced leaf senescence through interactions with INDUCER OF CBF EXPRESSION 1. J. Exp. Bot. 2020, 71, 1475–1490. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biotype | Dose (g a.i. ha−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| Resistant (R) | 1.875 | 3.75 | 7.5 | 15 | 30 | 60 | 120 | 240 |
| Susceptible (S) | 0.39 | 0.78 | 1.56 | 3.13 | 6.25 | 12.5 | 25 | 50 |
| Pathways | KO Name | Gene ID | Transcriptome log2FC * (FTR vs. UTR) | Transcriptome log2FC(FTS vs. UTS) | RT–qPCR log2FC(FTR vs. UTR) | RT–qPCR log2FC(FTS vs. UTS) |
|---|---|---|---|---|---|---|
| Auxin biosynthesis | ALDH | Cluster-7380.24619 | 2.20 | 3.15 | 1.33 | 3.05 |
| AAO1_2 | Cluster-7380.23124 | 1.62 | 2.46 | 1.02 | 1.93 | |
| Auxin signaling | IAA | Cluster-7380.10304 | 2.99 | 7.78 | 5.20 | 8.99 |
| IAA | Cluster-7380.17655 | 1.53 | 2.99 | 0.88 | 2.70 | |
| IAA | Cluster-7380.23356 | 1.54 | 3.45 | 3.24 | 3.53 | |
| IAA | Cluster-7380.32469 | 1.78 | 2.79 | 0.90 | 2.13 | |
| IAA | Cluster-7380.33766 | 1.68 | 1.52 | −0.11 | 1.94 | |
| IAA | Cluster-7380.38071 | 1.34 | 1.71 | 0.79 | 1.80 | |
| IAA | Cluster-7380.54703 | 1.42 | 4.53 | 1.07 | 3.46 | |
| GH3 | Cluster-7380.9345 | 8.16 | 12.43 | 6.77 | 10.19 | |
| GH3 | Cluster-7380.9346 | 6.06 | 9.03 | 4.04 | 8.13 | |
| SAUR | Cluster-7380.2810 | 0.30 | 3.47 | −2.18 | 3.37 | |
| SAUR | Cluster-7380.15012 | 4.90 | 3.10 | 3.74 | 2.76 | |
| ABA biosynthesis | NCED | Cluster-7380.40040 | 1.86 | 2.68 | 0.33 | 3.40 |
| ABA signaling | ABF | Cluster-7380.34634 | 1.59 | 1.95 | 0.50 | 1.60 |
| PP2C | Cluster-7380.45755 | 1.54 | 1.77 | 1.03 | 2.09 | |
| PP2C | Cluster-7380.827 | 3.11 | 5.79 | 1.52 | 3.22 | |
| SNRK2 | Cluster-7380.20927 | 1.60 | 2.83 | 0.10 | 3.44 | |
| ETH biosynthesis | ACS | Cluster-7380.61938 | 3.82 | 7.29 | 3.20 | 6.78 |
| ACO | Cluster-7380.43266 | 1.42 | 5.05 | 1.66 | 4.15 | |
| ACO | Cluster-7380.43267 | 2.87 | 5.96 | 1.38 | 4.08 | |
| ETH signaling | ETR, ERS | Cluster-7380.47127 | 0.49 | 1.09 | −0.34 | 1.48 |
| EBF1_2 | Cluster-7380.29178 | 0.76 | 2.43 | −0.33 | 3.34 | |
| ERF1 | Cluster-7380.8146 | 1.45 | 8.78 | −0.52 | 6.40 | |
| BR signaling | BRI1 | Cluster-7380.42410 | 0.57 | 1.28 | −0.48 | 0.95 |
| CTK signaling | AHK2_3_4 | Cluster-7380.32329 | −1.31 | −1.34 | −2.64 | −2.29 |
| AHP | Cluster-7380.62034 | −2.01 | −0.67 | −3.42 | −0.18 | |
| ARR-A | Cluster-7380.31589 | −2.10 | −1.92 | −3.54 | −2.19 | |
| ARR-A | Cluster-7380.34172 | −1.82 | −1.38 | −2.06 | −2.42 | |
| ARR-A | Cluster-7380.40603 | −1.16 | −1.16 | −2.62 | −1.84 | |
| ARR-A | Cluster-7380.40690 | −2.61 | −3.16 | −3.97 | −2.26 | |
| CTK biosynthesis | CKX | Cluster-7380.18387 | −3.68 | −2.08 | −4.54 | −1.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W.; Sun, J.; Tang, W.; Yang, Y.; Zhang, J.; Lu, Y.; Yu, X. Comparative Transcriptome Analysis of the Differential Effects of Florpyrauxifen-Benzyl Treatment on Phytohormone Transduction between Florpyrauxifen-Benzyl-Resistant and -Susceptible Barnyard Grasses (Echinochloa crus-galli (L.) P. Beauv). Agronomy 2023, 13, 702. https://doi.org/10.3390/agronomy13030702
Jin W, Sun J, Tang W, Yang Y, Zhang J, Lu Y, Yu X. Comparative Transcriptome Analysis of the Differential Effects of Florpyrauxifen-Benzyl Treatment on Phytohormone Transduction between Florpyrauxifen-Benzyl-Resistant and -Susceptible Barnyard Grasses (Echinochloa crus-galli (L.) P. Beauv). Agronomy. 2023; 13(3):702. https://doi.org/10.3390/agronomy13030702
Chicago/Turabian StyleJin, Wenyong, Jinqiu Sun, Wei Tang, Yongjie Yang, Jianping Zhang, Yongliang Lu, and Xiaoyue Yu. 2023. "Comparative Transcriptome Analysis of the Differential Effects of Florpyrauxifen-Benzyl Treatment on Phytohormone Transduction between Florpyrauxifen-Benzyl-Resistant and -Susceptible Barnyard Grasses (Echinochloa crus-galli (L.) P. Beauv)" Agronomy 13, no. 3: 702. https://doi.org/10.3390/agronomy13030702