
Heritability and Genetic Advance Estimates of Key Shea Fruit Traits

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Area
2.2. Data Collection
2.3. Data Analysis
| Source | D.F | Mean Square | Expected Mean Square |
| Replications | |||
| Genotype | |||
| Error |
3. Results
3.1. Morphological Variation
3.2. Multivariate Analysis
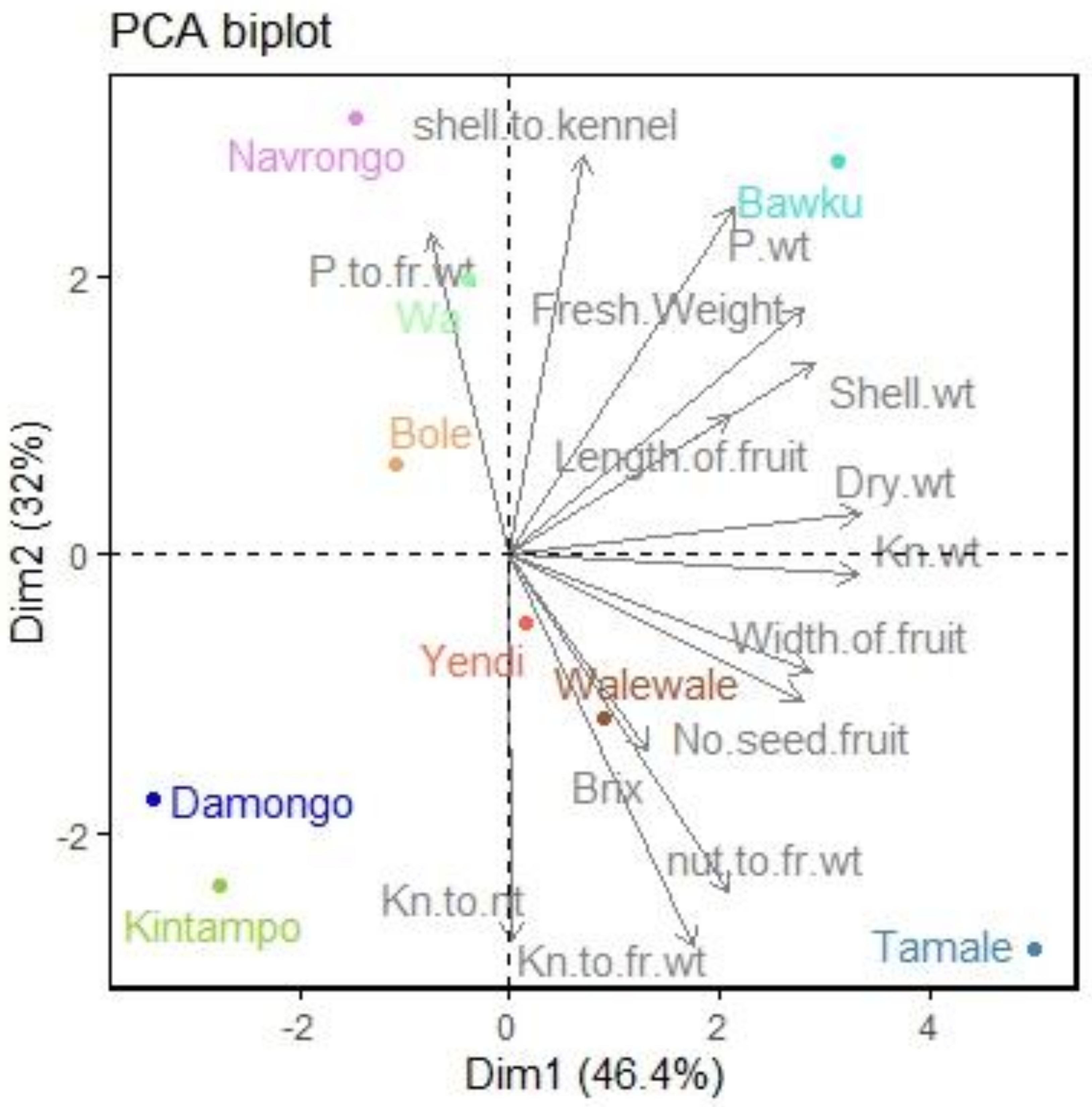
3.2.1. Principal Component Analysis
3.2.2. Morphological Clustering
3.2.3. Correlation among Traits
3.3. Qualitative Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nde, B.D.; Mohammed, M.A.; Cesar, K.; Zephirin, M. Production zones and systems, markets, benefits and constraints of shea (Vitellaria paradoxa Gaertn) butter processing. Oilseeds Fats Crops Lipids 2014, 21, D206. [Google Scholar] [CrossRef] [Green Version]
- Pulidindi, K.; Ahuja, K. Shea Butter Market Size by Product (Raw and Unrefined Shea Butter, Refined Shea Butter), By Application (Food & Beverage, Personal Care & Cosmetics and Pharmaceutical), COVID-19 impact analysis, Regional Outlook, Growth Potential, Competitive Market Share & Forecast, 2023–2032; Global Market Insight Incorporated: Sydney, Australia, 2023; Available online: https://www.gminsights.com/industry-analysis/shea-butter-market (accessed on 16 February 2023).
- Stout, J.C.; Nombre, I.; de Bruijn, B.; Delaney, A.; Doke, D.A.; Gyimah, T.; Kamano, F.; Kelly, R.; Lovett, P.; Marshall, E. Insect pollination improves yield of shea (Vitellaria paradoxa subsp. paradoxa) in the agroforestry parklands of west Africa. J. Pollinat. Ecol. 2018, 22, 11–20. [Google Scholar] [CrossRef]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestry Database: A Tree Reference and Selection Guide Version 4.0; World Agroforestry Centre: Nairobi, Kenya, 2009; Available online: http://worldagroforestry.org/output/agroforestree-database (accessed on 20 December 2022).
- NRC. Lost Crops of Africa. Volume II: Vegetables; The National Academies Press: Washington, DC, USA, 2006; p. 379. [Google Scholar]
- Gwali, S.; Nakabongea, G.; Okulloa, B.J.L.; Eilua, G.; Forestier-Chironc, N.; Piombod, G.; Davrieuxc, F. Fat content and fatty acid profiles of shea tree (Vitellaria paradoxa subspecies nilotica) ethno-varieties in Uganda. For. Trees Livelihoods 2012, 21, 267–278. [Google Scholar] [CrossRef]
- Chalfin, B. Shea Butter Republic; Routledge: New York, NY, USA, 2004. [Google Scholar]
- Maranz, S.; Kpikpi, W.; Wiesman, Z.; De Saint Sauveur, A.; Chapagain, B. Nutritional values and indigenous preferences for shea fruits (Vitellaria paradoxa C.V. Gaertn. F.) in African agroforestry parklands. Econ. Bot. 2004, 58, 588–600. [Google Scholar] [CrossRef]
- Konate, L. ‘Creating Competitive Market Models Burkina Faso: The Case of Nununa Women’s Shea Butter Federation’. Case Study. The Hague: SNV. Netherlands Development Organization: Case Studies Burkina-Shea. (2012). 11–13th April 2012 Published by SNV Netherlands Development Organisation. Available online: https://www.fao.org/sustainable-food-value-chains/library/details/ru/c/262506/ (accessed on 16 January 2023).
- Aguzue, O.C.; Akanji, F.T.; Tafida, M.A.; Kamal, M.J. Nutritional and some elemental composition of shea (Vitellaria paradoxa) fruit pulp. Scholars Research Library. Arch. Appl. Sci. Res. 2013, 5, 63–65. [Google Scholar]
- Adomako, D. Shea Nut Research Programme: Internal Memo; Physiology/Biochemistry Division, Cocoa Research Institute: New Tafo, Ghana, 1982. [Google Scholar]
- Gyedu-Akoto, E.; Amon-Armah, F.; Yabani, D. Utilization of shea fruit to enhance food security and reduce poverty in Ghana. Afr. J. Sci. Technol. Innov. Dev. 2017, 9, 697–705. [Google Scholar] [CrossRef]
- Boffa, J.M.; Yameogo, G.; Nikiema, P.; Taonda, J.B. What future for the Shea tree? Agrofor. Today 1996, 8, 5–9. [Google Scholar]
- Nyarko, G.; Mahunu, G.K.; Chimsah, F.A.; Yidana, J.A.; Abubakari, A.H.; Abagale, F.K.; Quainoo, A.; Poudyal, M. Leaf and fruit characteristics of Shea (Vitellaria paradoxa) in Northern Ghana. Res. Plant Biol. 2012, 2, 38–45. [Google Scholar]
- Hall, J.B.; Aebischer, D.P.; Tomlinson, H.F.; Osei-Amaning, E.; Hindle, J. Vitellaria Paradoxa: A Monograph; University of Wales: Bangor, UK, 1996; p. 105. [Google Scholar]
- Tom-Dery, D.; Eller, F.; Reisdorff, C. Shea (Vitellaria paradoxa C. F. Gaertn.) at the crossroads: Current knowledge and research gaps. Agrofor. Syst. 2018, 92, 1353–1371. [Google Scholar] [CrossRef]
- Sanou, H.; Kambou, S.; Teklehaimanot, Z.; Dembele, M.; Yossi, H.; Sina, S.; Djingdia, L. Vegetative propagation of Vitellaria paradoxa by grafting. Agrofor. Syst. 2004, 60, 93–99. [Google Scholar] [CrossRef]
- Yeboah, J.; Banful, B.K.; Boateng, P.Y.; Amoah, F.M.; Maalekuu, B.K.; Lowor, S.T. Rooting Response of Air-Layered Shea (Vitellaria paradoxa) Trees to Media and Hormonal Application under Two Different Climatic Conditions. Am. J. Plant Sci. 2014, 5, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- IPGRI; INIA. Descriptors for Shea Tree (Vitellaria paradoxa); International Plant Genetic Resources Institute: Rome, Italy; Instituto Nacional de Investigacion y Tecnologia Agrariay Alimentaria: Madrid, Spain, 2006. [Google Scholar]
- Popat, R.; Patel, R.; Parmer, D. Variability Package of R Statistical Software, 2020. R Package Version 0.1.0. Available online: http://CRAN.R-project.org/package (accessed on 1 September 2022).
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of genetic and environmental variability in soybean. J. Agron. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Burton, G.M.; Devane, E.M. Estimating heritability in tall Fescue from replication clonal material. Agron. J. 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Sivasubramanian, S.; Menon, M. Heterosis and inbreeding depression in rice. Madras Agric. J. 1973, 60, 1139. [Google Scholar]
- Allard, R.W. Principles of Plant Breeding; John Wiley and Sons Inc.: New York, NY, USA, 1960; p. 485. Available online: https://bit.ly/2NENkBv (accessed on 20 November 2022).
- Robinson, H.F.; Comstock, R.E.; Harvey, P.H. Genotypic and phenotypic correlation in corn and their implications in selection. Agron. J. 1949, 43, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Djekota, C.; Diouf, D.; Sane, S.; Mbaye, M.S.; Noba, K. Morphological characterization of shea tree (Vitellaria paradoxa subsp. paradoxa) populations in the region of Mandoul in Chad. Int. J. Biodivers. Conserv. 2014, 6, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Quainoo, A.K.; Jibrim, A.; Chimsah, F. Effect Of Shea Fruit Sugar Content On Shea (Vitellaria paradoxa L.) Butter Quality, UDS. Int. J. Dev. 2020, 7, 256–261. [Google Scholar]
- Agbahungba, G.; Depommier, D. Aspects du parc a karites-neres (Vitellaria paradoxa Gaertn. f. Parkia biglobosa Jacq. Benth) dans le Sud du Borgou (Benin). Bois For. Trop. 1989, 222, 41–54. [Google Scholar]
- Animasaun, D.A.; Oyedeji, S.; Olorunmaiye, K.S.; Azeez, M.A.; Tijani, I.T.; Morakinyo, J.A. Morpho-chemical divergence and fatty acid profile of shea tree seeds (Vitellaria paradoxa) collected from different locations in Kwara State, Nigeria. Acta Bot. Croatica 2019, 78, 17–24. [Google Scholar] [CrossRef]
- Diarrassouba, N.; Bup, N.D.; Kapseu, C.; Kouame, C.; Sangare, A. Phenotypic Diversity of Shea (Vitellaria paradoxa C. F. Gaertn.) Populations across Four Agro-Ecological Zones of Cameroon. J. Crop Sci. Biotechnol. 2007, 10, 211–218. [Google Scholar]
- Xia, Y.; Yu, K.; Navarre, D.; Seebold, K.; Kachroo, A.; Kachroo, P. The glabra1 mutation affects cuticle formation and plant responses to microbes. Plant Physiol. 2010, 154, 833–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stavrianakou, S.; Liakopoulos, G.; Miltiadou, D.; Markoglou, A.N.; Ziogas, B.N.; Karabourniotis, G. Antifungal and antibacterial capacity of extracted material from non-glandular and glandular leaf hairs applied at physiological concentrations. Plant Stress 2010, 4, 25–30. [Google Scholar]
- Karabourniotis, G.; Kotsabassidis, D.; Manetas, Y. Trichome density and its protective potential against ultraviolet-B radiation damage during leaf development. Can. J. Bot. 1995, 73, 376–383. [Google Scholar] [CrossRef]
- Nwangburuka, C.C.; Denton, O.A. Heritability, character association and genetic advance in six agronomic and yield related characters in leaf Corchorus olitorius. Int. J. Agric. Res. 2012, 7, 367–375. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Location | ID | Location | ID | Location |
|---|---|---|---|---|---|
| CRIG 189 | Bawku | CRIG 104 | Bole | CRIG KA 87 | Bole |
| G6 | Bawku | CRIG 105 | Bole | CRIG KA 24 | Bole |
| G2 | Bawku | CRIG R1NBT3 | Bole | CRIG KA 21 | Bole |
| CRIG PHBA 43 | Bole | CRIG R2CT1 | Bole | CRIG 91 | Damongo |
| CRIG PHBA 54 | Bole | CRIG R2CT2 | Bole | CRIG 90 | Damongo |
| CRIG PHBA 28 | Bole | CRIG R1ENT4 | Bole | CRIG 293 | Damongo |
| CRIG PHBA 50 | Bole | CRIG R2EBT1 | Bole | CRIG 84 | Damongo |
| CRIG PHBA 37 | Bole | CRIG R2NBT1 | Bole | CRIG 86 | Damongo |
| CRIG PHBA 51 | Bole | CRIG KRAAL | Bole | CRIG 85 | Damongo |
| CRIG PHBA 25 | Bole | CRIG J48 | Bole | CRIG 95 | Kintampo |
| CRIG P1R1T5 | Bole | CRIG J47 | Bole | CRIG 94 | Kintampo |
| CRIG EA 1 | Bole | CRIG GMSA | Bole | CRIG 93 | Kintampo |
| CRIG EA 4 | Bole | CRIG MB 13 | Bole | CRIG 15 | Kintampo |
| CRIG RH 1 | Bole | CRIG MB 16 | Bole | CRIG 17 | Kintampo |
| CRIG LAB C | Bole | CRIG MB 5 | Bole | CRIG 64 | Kintampo |
| CRIG SG130 | Bole | CRIG MB 14 | Bole | CRIG 169 | Navrongo |
| CRIG SG128 | Bole | CRIG KA 30 | Bole | CRIG 172 | Navrongo |
| CRIG SG170 | Bole | CRIG KA 11 | Bole | CRIG 39 | Tamale |
| CRIG SG129 | Bole | CRIG KA 105 | Bole | CRIG 510 | Wa |
| CRIG SG171 | Bole | CRIG KA 16 | Bole | CRIG 130 | Wa |
| CRIG SG302 | Bole | CRIG KA 09 | Bole | CRIG 125 | Wa |
| CRIG SG118 | Bole | CRIG KA 93 | Bole | CRIG 123 | Wa |
| CRIG SG254 | Bole | CRIG KA 1 | Bole | CRIG 136 | Wa |
| CRIG SG97 | Bole | CRIG KA 27 | Bole | CRIG 138 | Wa |
| CRIG SG100 | Bole | CRIG KA 110 | Bole | CRIG 126 | Wa |
| CRIG SG116 | Bole | CRIG KA 29 | Bole | CRIG 141 | Wa |
| CRIG SG282 | Bole | CRIG KA 3 | Bole | CRIG 137 | Wa |
| CRIG SG114 | Bole | CRIG KA 10 | Bole | CRIG 59 | Walewale |
| CRIG SG142 | Bole | CRIG KA 2 | Bole | CRIG 330 | Walewale |
| CRIG SG284 | Bole | CRIG KA 5 | Bole | CRIG 4 | Yendi |
| CRIG SG113 | Bole | CRIG KA 33 | Bole | CRIG 6 | Yendi |
| CRIG 107 | Bole | CRIG KA 40 | Bole |
| Trait | Mean | Range | Mean Square |
|---|---|---|---|
| Fruit weight (g) | 23.05 | 10.25–48.87 | 158.6 *** |
| Fruit length (mm) | 37.89 | 26.44–58.46 | 93.29 *** |
| Fruit width (mm) | 32.92 | 21.80–69.38 | 80.57 *** |
| Brix | 23.11 | 11.93–46.00 | 36.36 *** |
| Pulp weight (g) | 13.94 | 4.67–35.05 | 92.98 *** |
| Nut weight (g) | 5.82 | 2.77–12.10 | 8.35 *** |
| Shell weight (g) | 1.63 | 0.63–4.76 | 1.04 *** |
| Kernel weight (g) | 4.17 | 1.86–8.16 | 4.49 *** |
| %Pulp | 59.38 | 31.00–97.92 | 226.13 *** |
| Kernel to Nut ratio | 71.56 | 51.02–91.75 | 69.85 *** |
| Trait | GV | PV | GCV | PCV | H | GA% |
|---|---|---|---|---|---|---|
| Fruit weight | 48.36 | 61.88 | 30.17 | 34.13 | 0.78 | 54.94 |
| Fruit Length | 27.59 | 35.12 | 13.86 | 16.3 | 0.72 | 24.3 |
| Fruit width | 24.39 | 31.79 | 15 | 17.13 | 0.77 | 27.07 |
| Brix | 9.67 | 17.01 | 13.46 | 17.85 | 0.57 | 20.91 |
| Pulp Weight | 28.32 | 36.33 | 38.19 | 43.25 | 0.78 | 69.46 |
| Nut Weight | 2.56 | 3.23 | 27.51 | 30.9 | 0.79 | 50.45 |
| Shell Weight | 0.32 | 0.39 | 34.77 | 38.82 | 0.80 | 64.14 |
| Kernel Weight | 1.37 | 1.76 | 28.04 | 31.83 | 0.78 | 50.89 |
| %Pulp | 69.29 | 87.55 | 14.02 | 15.76 | 0.79 | 25.69 |
| Trait | Correlation | Cos2 | Contribution | |||
|---|---|---|---|---|---|---|
| Dim 1 | Dim 2 | Dim 1 | Dim 2 | Dim 1 | Dim 2 | |
| Fresh weight | 0.835 | 0.524 | 0.696 | 0.275 | 10.716 | 6.132 |
| Fruit length | 0.622 | 0.295 | 0.387 | 0.087 | 5.962 | 1.941 |
| Fruit width | 0.854 | −0.247 | 0.730 | 0.061 | 11.234 | 1.360 |
| Brix | 0.391 | −0.413 | 0.153 | 0.171 | 2.348 | 3.814 |
| Pulp weight | 0.634 | 0.739 | 0.403 | 0.546 | 6.194 | 12.183 |
| Dry nut weight | 0.990 | 0.086 | 0.981 | 0.007 | 15.092 | 0.164 |
| Shell weight | 0.860 | 0.410 | 0.739 | 0.168 | 11.371 | 3.750 |
| Kernel weight | 0.988 | −0.042 | 0.975 | 0.002 | 15.009 | 0.040 |
| Pulp to fresh weight | −0.224 | 0.682 | 0.05 | 0.465 | 0.773 | 10.394 |
| Kernel to fresh weight | 0.520 | −0.834 | 0.271 | 0.695 | 4.168 | 15.523 |
| Shell to kernel | 0.210 | 0.848 | 0.044 | 0.718 | 0.676 | 16.042 |
| Nut to fresh weight | 0.621 | −0.715 | 0.386 | 0.512 | 5.941 | 11.432 |
| Kernel to nut ratio | 0.004 | −0.822 | 0.000 | 0.676 | 0.000 | 15.104 |
| Number of seed/fruit | 0.827 | −0.308 | 0.683 | 0.095 | 10.515 | 2.122 |
| Variables | FWt | FL | FWd | Brix | PWt | DWt | SWt | KWt | %PWt | %KWt | DW to FW |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NF.L | 0.74 | ||||||||||
| F.Wd | 0.67 | 0.52 | |||||||||
| Brix | −0.03 | 0.003 | −0.06 | ||||||||
| Pulp Wt | 0.92 | 0.65 | 0.65 | −0.04 | |||||||
| Dry wt | 0.70 | 0.48 | 0.52 | −0.01 | 0.52 | ||||||
| Shell.Wt | 0.62 | 0.40 | 0.39 | −0.12 | 0.47 | 0.88 | |||||
| K. wt | 0.68 | 0.48 | 0.51 | 0.02 | 0.50 | 0.97 | 0.77 | ||||
| %pulp to wt | 0.37 | 0.22 | 0.32 | −0.08 | 0.68 | −0.04 | −0.02 | −0.05 | |||
| %K to F.wt | −0.42 | −0.34 | −0.23 | 0.08 | −0.54 | 0.27 | 0.11 | 0.35 | −0.56 | ||
| Dry to F.wt | −0.45 | −0.38 | −0.26 | 0.05 | −0.58 | 0.29 | 0.22 | 0.29 | −0.61 | 0.95 | |
| %K to Nut | 0.01 | 0.04 | 0.07 | 0.11 | −0.01 | 0.06 | −0.30 | 0.27 | −0.01 | 0.39 | 0.09 |
| Variable | Categories | Shape | Counts | Frequencies | % |
|---|---|---|---|---|---|
| Shape of fruit | 1 | Oblate | 1 | 1 | 1.053 |
| 2 | Spheroid | 8 | 8 | 8.421 | |
| 3 | Ellipsoid | 66 | 66 | 69.474 | |
| 4 | Oblong | 1 | 1 | 1.053 | |
| 6 | Inverted ovoid | 19 | 19 | 20.000 | |
| Fruit surface | 1 | Smooth | 50 | 50 | 52.632 |
| 2 | Rough | 45 | 45 | 47.368 | |
| Number of seed(s) per fruit | 1 | 92 | 92 | 96.842 | |
| 2 | 3 | 3 | 3.158 | ||
| Seed shape | 3 | Ellipsoid | 82 | 82 | 86.316 |
| 6 | Inverted ovoid | 13 | 13 | 13.684 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anyomi, W.E.; Barnor, M.T.; Danquah, A.; Ofori, K.; Padi, F.K.; Avicor, S.W.; Hale, I.; Danquah, E.Y. Heritability and Genetic Advance Estimates of Key Shea Fruit Traits. Agronomy 2023, 13, 640. https://doi.org/10.3390/agronomy13030640
Anyomi WE, Barnor MT, Danquah A, Ofori K, Padi FK, Avicor SW, Hale I, Danquah EY. Heritability and Genetic Advance Estimates of Key Shea Fruit Traits. Agronomy. 2023; 13(3):640. https://doi.org/10.3390/agronomy13030640
Chicago/Turabian StyleAnyomi, Wisdom Edem, Michael Teye Barnor, Agyemang Danquah, Kwadwo Ofori, Francis Kwame Padi, Silas Wintuma Avicor, Iago Hale, and Eric Yirenkyi Danquah. 2023. "Heritability and Genetic Advance Estimates of Key Shea Fruit Traits" Agronomy 13, no. 3: 640. https://doi.org/10.3390/agronomy13030640