
Application of Synthetic Consortia for Improvement of Soil Fertility, Pollution Remediation, and Agricultural Productivity: A Review
 ,
,  , , ,
, , ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
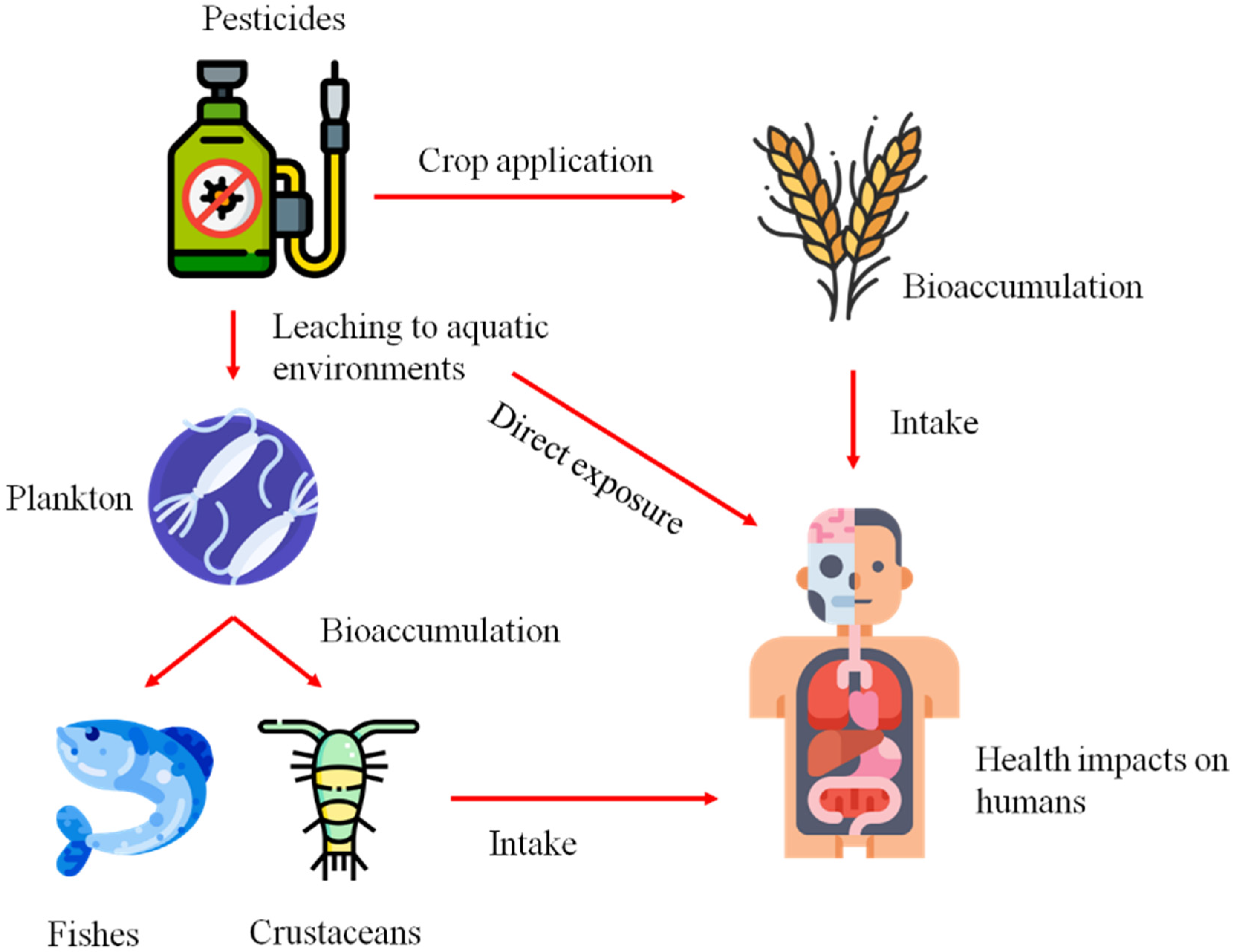
2. Synthetic Consortium as Bioremediation Agent of Pesticides
3. Synthetic Consortium as Bioremediation Agent of Heavy Metals
4. Bioengineering of Synthetic Consortia
5. Incorporation of Microbial Communities in Agricultural Nanotechnology
6. Role of Synthetic Consortium in the Improvement of Crop and Soil Properties
7. Role of Synthetic Consortium/Bio-formulation in Plant Disease Management
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte, C.E.; Purza, L.; Badea, G.E. Effects of long term application of organic and mineral fertilizers on soil enzymes. Rev. Chim. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Ali, S.S.; Kornaros, M.; Manni, A.; Sun, J.; El-Shanshoury, A.E.R.; Kenawy, E.R.; Khalil, M.A. Enhanced anaerobic digestion performance by two artificially constructed microbial consortia capable of woody biomass degradation and chlorophenols detoxification. J. Hazard. Mater. 2020, 389, 122076. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, M.; Zielinska, M.; Cydzik-Kwiatkowska, A.; Rusanowska, P.; Debowski, M. Effect of static magnetic field on microbial community during anaerobic digestion. Bioresour. Technol. 2021, 323, 124600. [Google Scholar] [CrossRef]
- Basak, B.; Patil, S.M.; Saha, S.; Kurade, M.B.; Ha, G.S.; Govindwar, S.P.; Lee, S.S.; Chang, S.W.; Chung, W.J.; Jeon, B.H. Rapid recovery of methane yield in organic overloaded-failed anaerobic digesters through bioaugmentation with acclimatized microbial consortium. Sci. Total Environ. 2021, 764, 144219. [Google Scholar] [CrossRef] [PubMed]
- McGachy, L.; Skarohlid, R.; Martinec, M.; Roskova, Z.; Smrhova, T.; Strejcek, M.; Uhlik, O.; Marek, J. Effect of chelated iron activated peroxydisulfate oxidation on perchloroethene-degrading microbial consortium. Chemosphere 2021, 266, 128928. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.; Pandey, A.; Upasani, V.N. Petroleum sludge polluted soil remediation: Integrated approach involving novel bacterial consortium and nutrient application. Sci. Total Environ. 2021, 763, 142934. [Google Scholar] [CrossRef]
- Wanapaisan, P.; Laothamteep, N.; Vejarano, F.; Chakraborty, J.; Shintani, M.; Muangchinda, C.; Morita, T.; Suzuki-Minakuchi, C.; Inoue, K.; Nojiri, H.; et al. Synergistic degradation of pyrene by five culturable bacteria in a mangrove sediment-derived bacterial consortium. J. Hazard. Mater. 2018, 342, 561–570. [Google Scholar] [CrossRef]
- Hou, D.; Zhang, P.; Wei, D.; Zhang, J.; Yan, B.; Cao, L.; Zhou, Y.; Luo, L. Simultaneous removal of iron and manganese from acid mine drainage by acclimated bacteria. J. Hazard. Mater. 2020, 396, 122631. [Google Scholar] [CrossRef]
- Wan, W.; Xing, Y.; Qin, X.; Li, X.; Liu, S.; Luo, X.; Huang, Q.; Chen, W. A manganese-oxidizing bacterial consortium and its biogenic Mn oxides for dye decolorization and heavy metal adsorption. Chemosphere 2020, 253, 126627. [Google Scholar] [CrossRef]
- Baltrenaite, E.; Baltrenas, P.; Lietuvninkas, A.; Sereviciene, V.; Zuokaite, E. Integrated evaluation of aerogenic pollution by air-transported heavy metals (Pb, Cd, Ni, Zn, Mn and Cu) in the analysis of the main deposit media. Environ. Sci. Pollut. Res. Int. 2014, 21, 299–313. [Google Scholar] [CrossRef]
- Yin, T.; Lin, H.; Dong, Y.; Li, B.; He, Y.; Liu, C.; Chen, X. A novel constructed carbonate-mineralized functional bacterial consortium for high-efficiency cadmium biomineralization. J. Hazard. Mater. 2020, 401, 123269. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Kaur, I.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Bungau, S. The dichotomy of nanotechnology as the cutting edge of agriculture: Nano-farming as an asset versus nanotoxicity. Chemosphere 2022, 288, 132533. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Sharma, A. Response of nanogypsum on the performance of plant growth promotory bacteria recovered from nanocompound infested agriculture field. Environ. Ecol. 2019, 37, 363–372. [Google Scholar]
- Chaudhary, A.; Chaudhary, P.; Upadhyay, A.; Kumar, A.; Singh, A. Effect of Gypsum on Plant Growth Promoting Rhizobacteria. Environ. Ecol. 2022, 39, 1248–1256. [Google Scholar]
- Kukreti, B.; Sharma, A.; Chaudhary, P.; Agri, U.; Maithani, D. Influence of nanosilicon dioxide along with bioinoculants on Zea mays and its rhizospheric soil. 3 Biotech 2020, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Sharma, A.; Chaudhary, P.; Khati, P. Management of plant vigor and soil health using two agriusable nanocompounds and plant growth promotory rhizobacteria in Fenugreek. 3 Biotech. 2020, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Egidi, E.; Qiu, Z.; Macdonald, C.A.; Verma, J.P.; Trivedi, P. Synthetic community improves crop performance and alters rhizosphere microbial communities. J. Sustain. Agric. Environ. 2022, 1, 118–131. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New frontiers in agriculture productivity: Optimised microbial inoculants and in situ microbiome engineering. Biotech Adv. 2019, 37, 107371. [Google Scholar] [CrossRef]
- Kaur, T.; Devi, R.; Kumar, S.; Sheikh, I.; Kour, D.; Yadav, A.N. Microbial consortium with nitrogen fixing and mineral solubilizing attributes for growth of barley (Hordeum vulgare L.). Heliyon 2022, 8, e09326. [Google Scholar] [CrossRef]
- Khati, P.; Parul; Bhatt, P.; Nisha; Kumar, R.; Sharma, A. Effect of nanozeolite and plant growth promoting rhizobacteria on maize. 3 Biotech 2018, 8, 141. [Google Scholar] [CrossRef]
- Agri, U.; Chaudhary, P.; Sharma, A. In vitro compatibility evaluation of agriusable nanochitosan on beneficial plant growth-promoting rhizobacteria and maize plant. Nat. Acad. Sci. Lett. 2021, 44, 555–559. [Google Scholar] [CrossRef]
- Agri, U.; Chaudhary, P.; Sharma, A.; Kukreti, B. Physiological response of maize plants and its rhizospheric microbiome under the influence of potential bioinoculants and nanochitosan. Plant Soil. 2022, 474, 451–468. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Tofazzal Islam, M. Co-inoculation with Enterobacter and rhizobacteria on yield and nutrient uptake by wheat (Triticum aestivum L.) in the alluvial soil under Indo-Gangetic plain of India. J. Plant Growth Regul. 2017, 36, 608–617. [Google Scholar] [CrossRef]
- Ghorchiani, M.; Etesami, H.; Alikhani, H.A. Improvement of growth and yield of maize under water stress by co-inoculating an arbuscular mycorrhizal fungus and a plant growth promoting rhizobacterium together with phosphate fertilizers. Agric. Ecosyst. Environ. 2018, 258, 59–70. [Google Scholar] [CrossRef]
- Laranjeira, S.; Fernandes-Silva, A.; Reis, S.; Torcato, C.; Raimundo, F.; Ferreira, L.; Carnide, V.; Marques, G. Inoculation of plant growth promoting bacteria and arbuscular mycorrhizal fungi improve chickpea performance under water deficit conditions. Appl. Soil Ecol. 2021, 164, 103927. [Google Scholar] [CrossRef]
- Tsolakidou, M.D.; Stringlis, I.A.; Fanega-Sleziak, N.; Papageorgiou, S.; Tsalakou, A.; Pantelides, I.S. Rhizosphere-enriched microbes as a pool to design synthetic communities for reproducible beneficial outputs. FEMS Microbiol. Ecol. 2019, 95, 138. [Google Scholar] [CrossRef] [PubMed]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; de Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.J.; Gomez-Exposito, R.; Elsayed, S.S.; et al. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Pandey, S.C.; Joshi, S.; Chaudhary, P.; Pathak, V.M.; Huang, Y.; Wu, X.; Zhou, Z.; Chen, S. Nanobioremediation: A sustainable approach for the removal of toxic pollutants from the environment. J. Hazard. Mater. 2022, 427, 128033. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, A.; Chaudhary, P.; Gangola, S. Chlorpyrifos degradation using binary fungal strains isolated from industrial waste soil. Biologia 2021, 76, 3071–3080. [Google Scholar] [CrossRef]
- Mulbry, W.W.; Ahrens, E.; Karns, J.S. Use of a field-scale biofilter for the degradation of the organophosphate insecticide coumaphos in cattle dip wastes. Pest. Sci. 1998, 52, 268–274. [Google Scholar] [CrossRef]
- López, L.; Pozo, C.; Rodelas, B.; Calvo, C.; Juárez, B.; Martínez-Toledo, M.V.; González-López, J. Identification of bacteria isolated from an oligotrophic lake with pesticide removal capacities. Ecotoxicology 2005, 14, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, Y.; Yao, T.; Ma, L.; Zhang, J.; Li, C. Biodegradation Pathway and Detoxification of β-cyfluthrin by the Bacterial Consortium and Its Bacterial Community Structure. J. Agric. Food Chem. 2022, 70, 7626–7635. [Google Scholar] [CrossRef] [PubMed]
- Satsuma, K. Characterisation of new strains of atrazine-degrading Nocardioides sp. isolated from Japanese riverbed sediment using naturally derived river ecosystem. Pest Manag. Sci. 2006, 62, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Chirnside, A.E.M.; Ritter, W.F.; Radosevich, M. Isolation of a selected microbial consortium from a pesticide-contaminated mix-load site soil capable of degrading the herbicides atrazine and alachlor. Soil Biol. Biochem. 2007, 39, 3056–3065. [Google Scholar] [CrossRef]
- Hussain, S.; Siddique, T.; Arshad, M.; Saleem, M. Bioremediation and Phytoremediation of Pesticides: Rec. Adv. Critic. Rev. Environm. Sci. Technol. 2009, 39, 843–907. [Google Scholar] [CrossRef]
- Wolfaardt, G.M.; Lawrence, J.R.; Robarts, R.D.; Caldwell, S.J.; Caldwell, D.E. Multicellular organization in a degradative biofilm community. Appl. Environ. Microbiol. 1994, 60, 434–446. [Google Scholar] [CrossRef] [Green Version]
- Baghapour, M.A.; Nasseri, S.; Derakhshan, Z. Atrazine removal from aqueous solutions using submerged biological aerated filter. J. Environ. Health Sci. Eng. 2013, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Villaverde, J.; Rubio-Bellido, M.; Lara, A.; Merchan, F.; Morillo, E. Combined use of microbial consortia isolated from different agricultural soils and cyclodextrin as a bioremediation technique for herbicide contaminated soils. Chemosphere 2017, 193, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Villaverde, J.; Rubio-Bellido, M.; Merchán, F.; Morillo, E. Bioremediation of diuron contaminated soils by a novel degrading microbial consortium. J. Environ. Manag. 2017, 188, 379–386. [Google Scholar] [CrossRef]
- Góngora-Echeverría, V.R.; García-Escalante, R.; Rojas-Herrera, R.; Giácoman-Vallejos, G.; Ponce-Caballero, C. Pesticide bioremediation in liquid media using a microbial consortium and bacteria-pure strains isolated from a biomixture used in agricultural areas. Ecotoxicol. Environm. Saf. 2020, 200, 110734. [Google Scholar] [CrossRef]
- Whittington, H.D.; Singh, M.; Ta, C.; Azcárate-Peril, M.A.; Bruno-Bárcena, J.M. Accelerated Biodegradation of the Agrochemical Ametoctradin by Soil-Derived Microbial Consortia. Front. Microbiol. 2020, 11, 1898. [Google Scholar] [CrossRef] [PubMed]
- Doolotkeldieva, T.; Bobusheva, S.; Konurbaeva, M. The Improving Conditions for the Aerobic Bacteria Performing the Degradation of Obsolete Pesticides in Polluted Soils. Air Soil Water Res. 2021, 14, 117862212098259. [Google Scholar] [CrossRef]
- Khatun, M.A.; Hoque, M.A.; Zhang, Y.; Lu, T.; Cui, L.; Zhou, N.-Y.; Feng, Y. Bacterial Consortium-Based Sensing System for Detecting Organophosphorus Pesticides. Anal. Chem. 2018, 90, 10577–10584. [Google Scholar] [CrossRef] [PubMed]
- Pattanasupong, A.; Nagase, H.; Sugimoto, E.; Hori, Y.; Hirata, K.; Tani, K.; Nasu, M.; Miyamoto, K. Degradation of carbendazim and 2,4-dichlorophenoxyacetic acid by immobilized consortium on loofa sponge. J. Biosci. Bioeng. 2004, 98, 28–33. [Google Scholar] [CrossRef]
- Paliwal, R.; Uniyal, S.; Rai, J.P.N. Evaluating the potential of immobilized bacterial consortium for black liquor biodegradation. Environ. Sci. Pollut. Res. 2015, 22, 6842–6853. [Google Scholar] [CrossRef]
- Moreno-Medina, D.A.; Sánchez-Salinas, E.; Ortiz-Hernández, M.L. Removal of methyl parathion and coumaphos pesticides by a bacterial consortium immobilized in Luffa cylindrica. Rev. Int. Contam. Ambiental. 2014, 30, 51–63. [Google Scholar]
- Geed, S.R.; Kureel, M.K.; Giri, B.S.; Singh, R.S.; Rai, B.N. Performance evaluation of Malathion biodegradation in batch and continuous packed bed bioreactor (PBBR). Bioresour. Technol. 2017, 227, 56–65. [Google Scholar] [CrossRef]
- Geed, S.R.; Kureel, M.K.; Prasad, S.; Singh, R.S.; Rai, B.N. Novel study on biodegradation of malathion and investigation of mass transfer correlation using alginate beads immobilized Bacillus sp. S4 in bioreactor. J. Environ. Chem. Eng. 2018, 6, 3444–3450. [Google Scholar] [CrossRef]
- Geetha, M.; Fulekar, M.H. Bioremediation of pesticides in surface soil treatment unit using microbial consortia. Afr. J. Environm. Sci. Technol. 2008, 2, 36–45. [Google Scholar]
- Dejonghe, W.; Berteloot, E.; Goris, J.; Boon, N.; Crul, K.; Maertens, S.; Höfte, M.; De Vos, P.; Verstraete, W.; Top, E.M. Synergistic Degradation of Linuron by a Bacterial Consortium and Isolation of a Single Linuron-Degrading Variovorax Strain. Appl. Environ. Microbiol. 2003, 69, 1532–1541. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Xue, R.; Zhou, J.; Wen, X.; Shi, Z.; Chen, M.; Xin, F.; Zhang, W.; Dong, W.; Jiang, M. Characterization of Acetamiprid Biodegradation by the Microbial Consortium ACE-3 Enriched from Contaminated Soil. Front. Microbiol. 2020, 11, 1429. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Briceño, G.E.; Saez, J.M.; Benimeli, C.S.; Diez, M.C.; Amoroso, M.J. Enhanced Removal of a Pesticides Mixture by Single Cultures and Consortia of Free and Immobilized Streptomyces Strains. BioMed Res. Int. 2013, 2013, 392573. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Saini, H.S.; Raj, M. Rhamnolipid mediated enhanced degradation of chlorpyrifos by bacterial consortium in soil-water system. Ecotoxicol. Environ. Saf. 2016, 134, 156–162. [Google Scholar] [CrossRef]
- Ortiz-Hernández, M.L.; Sánchez-Salinas, E. Biodegradation of the organophosphate pesticide tetrachlorvinphos by bacteria isolated from agricultural soils in México. Rev. Int. Contam. Ambient. 2010, 26, 27–38. [Google Scholar]
- Jariyal, M.; Jindal, V.; Mandal, K.; Gupta, V.K.; Singh, B. Bioremediation of organophosphorus pesticide phorate in soil by microbial consortia. Ecotoxicol. Environ. Saf. 2018, 159, 310–316. [Google Scholar] [CrossRef]
- Zhang, L.; Hang, P.; Hu, Q.; Chen, X.L.; Zhou, X.Y.; Chen, K.; Jiang, J.D. Degradation of Phenylurea Herbicides by a Novel Bacterial Consortium Containing Synergistically Catabolic Species and Functionally Complementary Hydrolases. J. Agric. Food Chem. 2018, 66, 12479–12489. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, X.; Wu, Y.; Li, J.; An, H.; Zhang, T. Enhancement of dicarboximide fungicides degradation by two bacterial cocultures of Providencia stuartii JD and Brevundimonas naejangsanensis J3. J. Haz. Mater. 2021, 403, 123888. [Google Scholar] [CrossRef] [PubMed]
- Al-Huqail, A.A.; Kumar, V.; Kumar, R.; Eid, E.M.; Taher, M.A.; Adelodun, B.; Abou Fayssal, S.; Mioć, B.; Držaić, V.; Goala, M.; et al. Sustainable Valorization of Four Types of Fruit Peel Waste for Biogas Recovery and Use of Digestate for Radish (Raphanus sativus L. cv. Pusa Himani) Cultivation. Sustainability 2022, 14, 10224. [Google Scholar] [CrossRef]
- Al-Huqail, A.A.; Kumar, P.; Eid, E.M.; Adelodun, B.; Abou Fayssal, S.; Singh, J.; Arya, A.K.; Goala, M.; Kumar, V.; Širić, I. Risk Assessment of Heavy Metals Contamination in Soil and Two Rice (Oryza sativa L.) Varieties Irrigated with Paper Mill Effluent. Agriculture 2022, 12, 1864. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, V.; Eid, E.M.; AL-Huqail, A.A.; Adelodun, B.; Abou Fayssal, S.; Goala, M.; Arya, A.K.; Bachheti, A.; Andabaka, Ž.; et al. Spatial Assessment of Potentially Toxic Elements (PTE) Concentration in Agaricus bisporus Mushroom Collected from Local Vegetable Markets of Uttarakhand State, India. J. Fungi 2022, 8, 452. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Eid, E.M.; Taher, M.A.; El-Morsy, M.H.E.; Osman, H.E.M.; Al-Bakre, D.A.; Adelodun, B.; Abou Fayssal, S.; Goala, M.; Mioč, B.; et al. Biotransforming the Spent Substrate of Shiitake Mushroom (Lentinula edodes Berk.): A Synergistic Approach to Biogas Production and Tomato (Solanum lycopersicum L.) Fertilization. Horticulturae 2022, 8, 479. [Google Scholar] [CrossRef]
- Širić, I.; Eid, E.M.; Taher, M.A.; El-Morsy, M.H.E.; Osman, H.E.M.; Kumar, P.; Adelodun, B.; Abou Fayssal, S.; Mioč, B.; Andabaka, Ž.; et al. Combined Use of Spent Mushroom Substrate Biochar and PGPR Improves Growth, Yield, and Biochemical Response of Cauliflower (Brassica oleracea var. botrytis): A Preliminary Study on Greenhouse Cultivation. Horticulturae 2022, 8, 830. [Google Scholar] [CrossRef]
- Širić, I.; Eid, E.M.; El-Morsy, M.H.E.; Osman, H.E.M.; Adelodun, B.; Abou Fayssal, S.; Mioč, B.; Goala, M.; Singh, J.; Bachheti, A.; et al. Health Risk Assessment of Hazardous Heavy Metals in Two Varieties of Mango Fruit (Mangifera indica L. var. Dasheri and Langra). Horticulturae 2022, 8, 832. [Google Scholar] [CrossRef]
- Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H.; El Sebaaly, Z.; Najjar, R.; Sassine, Y.N. Effect of olive pruning residues on substrate temperature and production of oyster mushroom (Pleurotus ostreatus). Acta Hortic. 2021, 1327, 245–252. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, V.; Adelodun, B.; Bedeković, D.; Kos, I.; Širić, I.; Alamri, S.A.M.; Alrumman, S.A.; Eid, E.M.; Abou Fayssal, S.; et al. Sustainable Use of Sewage Sludge as a Casing Material for Button Mushroom (Agaricus bisporus) Cultivation: Experimental and Prediction Modeling Studies for Uptake of Metal Elements. J. Fungi 2022, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Eid, E.M.; Al-Huqail, A.A.; Širić, I.; Adelodun, B.; Abou Fayssal, S.; Valadez-Blanco, R.; Goala, M.; Ajibade, F.O.; Choi, K.S.; et al. Kinetic Studies on Delignification and Heavy Metals Uptake by Shiitake (Lentinula edodes) Mushroom Cultivated on Agro-Industrial Wastes. Horticulturae 2022, 8, 316. [Google Scholar] [CrossRef]
- Širić, I.; Kumar, P.; Adelodun, B.; Abou Fayssal, S.; Bachheti, R.K.; Bachheti, A.; Ajibade, F.O.; Kumar, V.; Taher, M.A.; Eid, E.M. Risk Assessment of Heavy Metals Occurrence in Two Wild Edible Oyster Mushrooms (Pleurotus spp.) Collected from Rajaji National Park. J. Fungi 2022, 8, 1007. [Google Scholar] [CrossRef]
- Elbagory, M.; El-Nahrawy, S.; Omara, A.E.-D.; Eid, E.M.; Bachheti, A.; Kumar, P.; Abou Fayssal, S.; Adelodun, B.; Bachheti, R.K.; Kumar, P.; et al. Sustainable Bioconversion of Wetland Plant Biomass for Pleurotus ostreatus var. florida Cultivation: Studies on Proximate and Biochemical Characterization. Agriculture 2022, 12, 2095. [Google Scholar] [CrossRef]
- Werghemmi, W.; Abou Fayssal, S.; Mazouz, H.; Hajjaj, H.; Hajji, L. Olive and Green Tea Leaves Extract in Pleurotus ostreatus var. florida Culture Media: Effect on Mycelial Linear Growth Rate, Diameter and Growth Induction Index. IOP Conf. Ser. Earth Environ. Sci. 2022, 1090, 012020. [Google Scholar] [CrossRef]
- Kumar, P.; Eid, E.M.; Taher, M.A.; El-Morsy, M.H.E.; Osman, H.E.M.; Al-Bakre, D.A.; Adelodun, B.; Abou Fayssal, S.; Andabaka, Ž.; Goala, M.; et al. Sustainable Upcycling of Mushroom Farm Wastewater through Cultivation of Two Water Ferns (Azolla spp.) in Stagnant and Flowing Tank Reactors. Horticulturae 2022, 8, 506. [Google Scholar] [CrossRef]
- Taher, M.A.; Zouidi, F.; Kumar, P.; Abou Fayssal, S.; Adelodun, B.; Goala, M.; Kumar, V.; Andabaka, Ž.; Širić, I.; Eid, E.M. Impact of Irrigation with Contaminated Water on Heavy Metal Bioaccumulation in Water Chestnut (Trapa natans L.). Horticulturae 2023, 9, 190. [Google Scholar] [CrossRef]
- Latorre, M.; Cortés, M.P.; Travisany, D.; Genova, A.D.; Budinich, M.; Reyes-Jara, A.; Hödar, C.; González, M.; Parada, P.; Bobadilla-Fazzini, R.A.; et al. The bioleaching potential of a bacterial consortium. Bioresour. Technol. 2016, 218, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Bayer, T.S. Engineering microbial consortia to enhance biomining and bioremediation. Front. Microbiol. 2012, 3, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roane, T.M.; Josephson, K.L.; Pepper, I.L. Dual-bioaugmentation strategy to enhance remediation of co-contaminated soil. Appl. Environ. Microbiol. 2001, 67, 3208–3215. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, F.; Brunet, C.; van Zyl, L.J.; Trindade, M.; Golyshin, P.N.; Dell’Anno, A.; Ianora, A.; Sansone, C. Degradation of Hydrocarbons and Heavy Metal Reduction by Marine Bacteria in Highly Contaminated Sediments. Microorganisms 2020, 8, 1402. [Google Scholar] [CrossRef]
- Duarte, G.F.; Rosado, A.S.; Seldin, L.; de Araujo, W.; van Elsas, J.D. Analysis of Bacterial Community Structure in Sulfurous-Oil-Containing Soils and Detection of Species Carrying Dibenzothiophene Desulfurization (dsz) Genes. Appl. Env. Microbiol. 2001, 67, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.-L. Hexavalent chromium reduction and precipitation by sulphate reducing bacterial biofilms. Environ. Geochem. Health 2001, 23, 297–300. [Google Scholar] [CrossRef]
- Kang, C.-H.; Kwon, Y.-J.; So, J.-S. Bioremediation of heavy metals by using bacterial mixtures. Ecol. Eng. 2016, 89, 64–69. [Google Scholar] [CrossRef]
- Lee, Y.; Jeong, S.E.; Hur, M.; Ko, S.; Jeon, C.O. Construction and Evaluation of a Korean Native Microbial Consortium for the Bioremediation of Diesel Fuel-Contaminated Soil in Korea. Front. Microbiol. 2018, 9, 2594. [Google Scholar] [CrossRef]
- Polti, M.A.; Aparicio, J.D.; Benimeli, C.S.; Amoroso, M.J. Simultaneous bioremediation of Cr (VI) and lindane in soil by actinobacteria. Int. Biodeter. Biodegr. 2014, 88, 48–55. [Google Scholar] [CrossRef]
- Fulekar, M.H.; Sharma, J.; Tendulkar, A. Bioremediation of heavy metals using biostimulation in laboratory bioreactor. Env. Monit. Assess. 2012, 184, 7299–7307. [Google Scholar] [CrossRef] [PubMed]
- Dary, M.; Chamber-Pérez, M.A.; Palomares, A.J.; Pajuelo, E. ‘‘In situ’’ phytostabilisation of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant-growth promoting rhizobacteria. J. Hazard. Mat. 2010, 177, 323–330. [Google Scholar] [CrossRef] [PubMed]
- De, J.; Ramaiah, N.; Vardanyan, L. Detoxification of toxic heavy metals by marine bacteria highly resistant to mercury. Mar. Biotechnol. 2008, 10, 471–477. [Google Scholar] [CrossRef]
- Nwaehiri, U.L.; Akwukwaegbu, P.I.; Nwoke, B.E.B. Bacterial remediation of heavy metal polluted soil and effluent from paper mill industry. Environ. Anal. Health Toxicol. 2019, 35, e2020009. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zhao, J.; Bonet-Garcia, N.; Villagrasa, E.; Solé, A.; Liao, X.; Palet, C. Insights of microorganisms role in rice and rapeseed wastes as potential sorbents for metal removal. Int. J. Environ. Sci. Technol. 2022, 20, 801–814. [Google Scholar] [CrossRef]
- Talukdar, D.; Jasrotia, T.; Sharma, R.; Jaglan, S.; Kumar, R.; Vats, R.; Kumar, R.; Umar, A. Evaluation of novel indigenous fungal consortium for enhanced bioremediation of heavy metals from contaminated sites. Environ. Technol. Innov. 2020, 20, 101050. [Google Scholar] [CrossRef]
- Ma, L.; Xu, J.; Chen, N.; Li, M.; Feng, C. Microbial reduction fate of chromium (Cr) in aqueous solution by mixed bacterial consortium. Ecotoxicol. Environ. Saf. 2019, 170, 763–770. [Google Scholar] [CrossRef]
- Sprocati, A.R.; Alisi, C.; Tasso, F.; Marconi, P.; Sciullo, A.; Pinto, V.; Chiavarini, S.; Ubaldi, C.; Cremisini, C. Effectiveness of a microbial formula, as a bioaugmentation agent, tailored for bioremediation of diesel oil and heavy metal co-contaminated soil. Process. Biochem. 2012, 47, 1649–1655. [Google Scholar] [CrossRef]
- Lampis, S.; Santi, C.; Ciurli, A.; Andreolli, M.; Vallini, G. Promotion of arsenic phytoextraction efficiency in the fern Pteris vittata by the inoculation of As-resistant bacteria: A soil bioremediation perspective. Front. Plant Sci. 2015, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Souche, Y.; Goel, R. Comparative in situ remediation potential of Pseudomonas putida 710A and Commamonas aquatica 710B using plant (Vigna radiata (L.) wilczek) assay. Ann. Microbiol. 2012, 63, 923–928. [Google Scholar] [CrossRef]
- Belimov, A.A.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Safronova, V.I.; Litvinskiy, V.A.; Nosikov, V.V.; Zavalin, A.A.; Tikhonovich, I.A. Microbial Consortium of PGPR, Rhizobia and Arbuscular Mycorrhizal Fungus Makes Pea Mutant SGECdt Comparable with Indian Mustard in Cadmium Tolerance and Accumulation. Plants 2020, 9, 975. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Hamid, Y.; Zehra, A.; Sahito, Z.A.; He, Z.; Beri, W.T.; Khan, M.B.; Yang, X. Fava bean intercropping with Sedum alfredii inoculated with endophytes enhances phytoremediation of cadmium and lead co-contaminated field. Environ. Pollut. 2020, 265, 114861. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Hamid, Y.; Zehra, A.; Shohag, M.J.I.; He, Z.; Yang, X. Endophytic inoculation coupled with soil amendment and foliar inhibitor ensure phytoremediation and argo-production in cadmium contaminated soil under oilseed rape-rice rotation system. Sci. Total Environ. 2020, 748, 142481. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, M.T.; Tahseen, R.; Ashraf, A.; Mahmood, A.; Najam-ul-haq, M.; Arslan, M.; Afzal, M. Effective plant-endophyte interplay can improve the cadmium hyperaccumulation in Brachiaria mutica. World J. Microbiol. Biotech. 2019, 35, 188. [Google Scholar] [CrossRef] [PubMed]
- Mee, M.T.; Collins, J.J.; Church, G.M.; Wang, H.H. Syntrophic exchange in synthetic microbial communities. Proc. Nat. Acad. Sci. USA 2014, 111, 2149–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, T.; Ouyang, S.; Zhou, Q. Synthesis, characterization, safety design, and application of NPs@BC for contaminated soil remediation and sustainable agriculture. Biochar 2023, 5, 5. [Google Scholar] [CrossRef]
- Miu, B.A.; Dinischiotu, A. New Green Approaches in Nanoparticles Synthesis: An Overview. Molecules 2022, 27, 6472. [Google Scholar] [CrossRef]
- Bhardwaj, B.; Singh, P.; Kumar, A.; Kumar, S.; Budhwar, V. Eco-Friendly Greener Synthesis of Nanoparticles. Adv. Pharm. Bull. 2020, 10, 566–576. [Google Scholar] [CrossRef]
- Carrapiço, A.; Martins, M.R.; Caldeira, A.T.; Mirão, J.; Dias, L. Biosynthesis of Metal and Metal Oxide Nanoparticles Using Microbial Cultures: Mechanisms, Antimicrobial Activity and Applications to Cultural Heritage. Microorganisms 2023, 11, 378. [Google Scholar] [CrossRef]
- Boroumand Moghaddam, A.; Namvar, F.; Moniri, M.; Tahir, P.; Azizi, S.; Mohamad, R. Nanoparticles Biosynthesized by Fungi and Yeast: A Review of Their Preparation, Properties, and Medical Applications. Molecules 2015, 20, 16540–16565. [Google Scholar] [CrossRef]
- Adebayo, E.A.; Azeez, M.A.; Alao, M.B.; Oke, A.M.; Aina, D.A. Fungi as veritable tool in current advances in nanobiotechnology. Heliyon 2021, 7, e08480. [Google Scholar] [CrossRef] [PubMed]
- Dameron, C.T.; Reese, R.N.; Mehra, R.K.; Kortan, A.R.; Carroll, P.J.; Steigerwald, M.L.; Brus, L.E.; Winge, D.R. Biosynthesis of cadmium sulphide quantum semiconductor crystallites. Nature 1989, 338, 596–597. [Google Scholar] [CrossRef]
- Soliman, H.; Elsayed, A.; Dyaa, A. Antimicrobial activity of silver nanoparticles biosynthesised by Rhodotorula sp. strain ATL72. Egypt. J. Bas. Appl. Sci. 2018, 5, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; He, X.; Wang, K.; Yang, X. Different Active Biomolecules Involved in Biosynthesis of Gold Nanoparticles by Three Fungus Species. J. Biomed. Nanotechnol. 2011, 7, 245–254. [Google Scholar] [CrossRef]
- Składanowski, M.; Wypij, M.; Laskowski, D.; Golińska, P.; Dahm, H.; Rai, M. Silver and gold nanoparticles synthesized from Streptomyces sp. isolated from acid forest soil with special reference to its antibacterial activity against pathogens. J. Clust. Sci. 2016, 28, 59–79. [Google Scholar] [CrossRef] [Green Version]
- Liao, Q.; He, L.; Tu, G.; Yang, Z.; Yang, W.; Tang, J.; Cao, W.; Wang, H. Simultaneous immobilization of Pb, Cd and As in soil by hybrid iron-, sulfate- and phosphate-based bio-nanocomposite: Effectiveness, long-term stability and bioavailablity/bioaccessibility evaluation. Chemosphere 2020, 266, 128960. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Mechanistically compatible mixtures of bacterial antagonists improve biological control of fire blight of pear. Phytopathology 2011, 101, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Panwar, M.; Tewari, R.; Nayyar, H. Microbial Consortium of Plant Growth-Promoting Rhizobacteria Improves the Performance of Plants Growing in Stressed Soils: An Overview. In Phosphate Solubilizing Microorganisms: Principles and Application of Microphos Technology; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2014; pp. 257–285. [Google Scholar] [CrossRef]
- Sarma, B.K.; Yadav, S.K.; Singh, S.; Singh, H.B. Microbial consortium-mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem. 2015, 87, 25–33. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzmán-Guzmán, P.; Parra-Cota, F.I.; Santos-Villalobos, S.d.l.; Orozco-Mosqueda, M.d.C.; Glick, B.R. Plant Growth Stimulation by Microbial Consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Mueller, C.W.; Carminati, A.; Kaiser, C.; Subke, J.A.; Gutjahr, C. Rhizosphere functioning and structural development as complex interplay between plants, microorganisms and soil minerals. Front. Environ. Sci. 2019, 7, 130. [Google Scholar] [CrossRef] [Green Version]
- Valentine, D.L. Opinion: Adaptations to energy stress dictate the ecology and evolution of the archaea. Nat. Rev. Microbiol. 2007, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Maiyappan, S.; Amalraj, E.L.D.; Santhosh, A.; Peter, A.J. Isolation, evaluation and formulation of selected microbial consortia for sustainable agriculture. J. Biofertil. Biopestic. 2010, 2, 109–121. [Google Scholar]
- Pandey, P.; Maheshwari, D.K. Bioformulation of Burkholderia sp. MSSP with a multispecies consortium for growth promotion of Cajanus cajan. Can. J. Microbiol. 2007, 53, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lu, X.; Wu, M.; Zhang, H.; Pan, R.; Tian, J.; Li, S.; Liao, H. Co-inoculation with rhizobia and AMF inhibited soybean red crown rot: From field study to plant defense-related gene expression analysis. PLoS ONE 2012, 7, e33977. [Google Scholar] [CrossRef] [PubMed]
- Ardakani, M.R.; Pietsch, G.; Moghaddam, A.; Raza, A.; Friedel, J.K. Response of root properties to tripartite symbiosis between lucerne (Medicago sativa L.), rhizobia and mycorrhiza under dry organic farming conditions. Am. J. Agric. Biol. Sci. 2009, 4, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Tavasolee, A.N.; Aliasgharzad, G.; Salehijouzani, M.M.; Asgharzadeh, A. Interactive effects of arbuscular mycorrhizal fungi and rhizobial strains on Chickpea growth and nutrient content in plant. Afr. J. Microbiol. 2011, 10, 7585–7591. [Google Scholar]
- Bhattacharjee, S.G.; Sharma, D. Effect of dual inoculation of Arbuscular Mycorrhiza and Rhizobium on the chlorophyll, nitrogen and phosphorus contents of pigeon pea (Cajanus cajan L.). Adv. Microbiol. 2012, 2, 561–564. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Gray, V.M.; Straker, C.J. The influence of Rhizobium and arbuscular mycorrhizal fungi on nitrogen and phosphorus accumulation by Vicia faba. Ann. Bot. 2004, 94, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Messele, B.; Pant, L.M. Effects of inoculation of Sinorhizobium cicero and phosphate solubilizing bacteria on nodulation, yield and nitrogen and phosphorus uptake of chickpea (Cicer arietinum L.) in Shoa Robit Area. J. Biofertil. Biopestic. 2012, 3, 1000129. [Google Scholar] [CrossRef]
- Zayadan, B.K.; Matorin, D.N.; Baimakhanova, G.B.; Bolathan, K.; Oraz, G.D.; Sadanov, A.K. Promising microbial consortia for producing biofertilizers for rice fields. Microbiology 2014, 83, 391–397. [Google Scholar] [CrossRef]
- Hashmi, I.; Paul, C.; Al-Dourobi, A.; Sandoz, F.; Deschamps, P.; Junier, T.; Junier, P.; Bindschedler, S. Comparison of the plant growth-promotion performance of a consortium of Bacilli inoculated as endospores or as vegetative cells. FEMS Microbiol. Ecol. 2019, 95, 147. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional soil microbiome: Belowground solutions to an aboveground problem. Plant Physiol. 2014, 166, 689–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, O.M.; Castrillo, G.; Paredes, S.H.; González, I.S.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Cur. Op. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Lyu, D.; Msimbira, L.A.; Nazari, M.; Antar, M.; Pagé, A.; Shah, A.; Monjezi, N.; Zajonc, J.; Tanney, C.A.; Backer, R.; et al. The coevolution of plants and microbes underpins sustainable agriculture. Microorganisms 2021, 9, 1036. [Google Scholar] [CrossRef]
- Nehal, N.; Rathore, U.S.; Sharma, N. Microbes and soil health for sustainable crop production. In Microbial Metatranscriptomics Belowground; Nath, M., Bhatt, D., Bhargava, P., Choudhari, D.K., Eds.; Springer: Singapore, 2021; pp. 581–613. [Google Scholar] [CrossRef]
- Lynch, J.M.; Benedetti, A.; Insam, H.; Nuti, M.P.; Smalla, K.; Torsvik, V.; Nannipieri, P. Microbial diversity in soil: Ecological theories, the contribution of molecular techniques and the impact of transgenic plants and transgenic microorganisms. Biol. Fertil. Soils 2004, 40, 363–385. [Google Scholar] [CrossRef]
- Piazza, G.; Pellegrino, E.; Moscatelli, M.C.; Ercoli, L. Long-term conservation tillage and nitrogen fertilization effects on soil aggregate distribution, nutrient stocks and enzymatic activities in bulk soil and occluded microaggregates. Soil Tillage Res. 2020, 196, 104482. [Google Scholar] [CrossRef]
- Cardoso, E.J.B.N.; Vasconcellos, R.L.F.; Bini, D.; Miyauchi, M.Y.H.; Santos, C.A.D.; Alves, P.R.L.; Paula, A.M.D.; Nakatani, A.S.; Pereira, J.D.M.; Nogueira, M.A. Soil health: Looking for suitable indicators. What should be considered to assess the effects of use and management on soil health? Sci. Agric. 2013, 70, 274–289. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.L.E.; Glick, B.R. Applications of Plant Growth-Promoting Bacteria for Plant and Soil Systems. In Applications of Microbial Engineering; Gupta, V., Schmoll, M., Maki, M., Tuohy, M., Mazutti, M.A., Eds.; Taylor and Francis: Enfield, CT, USA, 2013; pp. 181–229. [Google Scholar]
- Villalobos, S.d.l.S.; Cota, F.I.P.; Sepúlveda, A.H.; Aragón, B.V.; Mora, J.C.E. Colmena: Colección de microorganismos edáficos y endófitos nativos, para contribuir a la seguridad alimentaria nacional. Rev. Mex. Cienc. Agríc. 2018, 9, 191–202. (In Spanish) [Google Scholar]
- Ju, W.; Liu, L.; Fang, L.; Cui, Y.; Duan, C.; Wu, H. Impact of co-inoculation with plant-growth-promoting rhizobacteria and rhizobium on the biochemical responses of alfalfa-soil system in copper contaminated soil. Ecotoxicol. Environ. Saf. 2019, 167, 218–226. [Google Scholar] [CrossRef]
- Rana, A.; Saharan, B.; Nain, L.; Prasanna, R.; Shivay, Y.S. Enhancing micronutrient uptake and yield of wheat through bacterial PGPR consortia. Soil Sci. Plant Nutr. 2012, 58, 573–582. [Google Scholar] [CrossRef]
- Gosal, S.K.; Kaur, J. Microbial Inoculants: A Novel Approach for Better Plant Microbiome Interactions. In Probiotics in Agroecosystem; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 269–289. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere microbiome manipulation for sustainable crop production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.E.; Teo, W.F.A.; Teoh, E.Y.; Tan, B.C. Microbiome engineering and plant biostimulants for sustainable crop improvement and mitigation of biotic and abiotic stresses. Discover. Food 2022, 2, 1–23. [Google Scholar] [CrossRef]
- Baetz, U.; Martinoia, E. Root exudates: The hidden part of plant defense. Trends Plant Sci. 2014, 19, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Khan, M.S.A.; Aqil, F.; Singh, M. Microbial applications in agriculture and the environment: A broad perspective. In Microbes and Microbial Technology: Agricultural and Environmental Applications; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 1–27. [Google Scholar]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B. Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Springer: Cham, Switzerland, 2015; p. 448. [Google Scholar]
- Schoebitz, M.; López, M.D.; Serrí, H.; Martínez, O.; Zagal, E. Combined application of microbial consortium and humic substances to improve the growth performance of blueberry seedlings. J. Soil Sci. Plant Nutr. 2016, 16, 1010–1023. [Google Scholar] [CrossRef]
- Jain, D.; Meena, R.H.; Choudhary, J.; Sharma, S.K.; Chauhan, S.; Bhojiya, A.A.; Khandelwal, S.K.; Mohanty, S.R. Effect of microbial consortia on growth and yield of wheat under typic haplustepts. Plant Physiol. Rep. 2021, 26, 570–580. [Google Scholar] [CrossRef]
- Hu, J.; Yang, T.; Friman, V.P.; Kowalchuk, G.A.; Hautier, Y.; Li, M.; Wei, Z.; Xu, Y.; Shen, Q.; Jousset, A. Introduction of probiotic bacterial consortia promotes plant growth via impacts on the resident rhizosphere microbiome. Proc. R. Soc. 2021, 288, 20211396. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Bai, Z.; Wu, S.; Li, X.; Wang, N.; Du, X.; Fan, H.; Zhuang, G.; Bohu, T.; et al. Unraveling mechanisms and impact of microbial recruitment on oilseed rape (Brassica napus L.) and the rhizosphere mediated by plant growth-promoting rhizobacteria. Microorganisms 2021, 9, 161. [Google Scholar] [CrossRef]
- Shen, H.; He, X.; Liu, Y.; Chen, Y.; Tang, J.; Guo, T. A complex inoculant of N2-fixing, P- and K-solubilizing bacteria from a purple soil improves the growth of kiwifruit (Actinidia chinensis) plantlets. Front. Microbiol. 2016, 7, 841. [Google Scholar] [CrossRef] [PubMed]
- Mengual, C.; Schoebitz, M.; Azćon, R.; Roldán, A. Microbial inoculants and organic amendment improves plant establishment and soil rehabilitation under semiarid conditions. J. Environ. Manag. 2014, 134, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Swarnalakshmi, K.; Prasanna, R.; Kumar, S.; Pattnaik, S.; Chakravarty, K.; Shivay, Y.S.; Singh, R.; Saxena, A.K. Evaluating the influence of novel cyanobacterial biofilmed biofertilizers on soil fertility and plant nutrition in wheat. Eur. J. Soil Biol. 2013, 55, 107–116. [Google Scholar] [CrossRef]
- Prasanna, R.; Triveni, S.; Bidyarani, N.; Babu, S.; Yadav, K.; Adak, A.; Khetarpal, S.; Pal, M.; Shivay, Y.S.; Saxena, A.K. Evaluating the efficacy of cyanobacterial formulations and biofilmed inoculants for leguminous crops. Arch. Agron. Soil Sci. 2014, 60, 349–366. [Google Scholar] [CrossRef]
- Zhang, J.; Ahmed, W.; Dai, Z.; Zhou, X.; He, Z.; Wei, L.; Ji, G. Microbial consortia: An engineering tool to suppress clubroot of Chinese cabbage by changing the rhizosphere bacterial community composition. Biology 2022, 11, 918. [Google Scholar] [CrossRef] [PubMed]
- Bradáčová, K.; Kandeler, E.; Berger, N.; Ludewig, U.; Neumann, G. Microbial consortia inoculants stimulate early growth of maize depending on nitrogen and phosphorus supply. Plant Soil Env. 2020, 66, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Sharma, S.; Kunal; Gosal, S.K.; Choudhary, R.; Singh, R.; Adholeya, A.; Singh, B. Synergistic use of plant growth-promoting Rhizobacteria, arbuscular mycorrhizal fungi, and spectral properties for improving nutrient use efficiencies in wheat (Triticum aestivum L.). Commun. Soil Sci. Plant Anal. 2020, 51, 14–27. [Google Scholar] [CrossRef]
- Ficke, A.; Cowger, C.; Bergstrom, G.; Brodal, G. Understanding Yield Loss and Pathogen Biology to Improve Disease Management: Septoria Nodorum Blotch—A Case Study in Wheat. Plant Dis. 2018, 102, 696–707. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Srivastava, M.; Kumar, A.; Singh, A.K.; Pandey, K.D. Endophytic bacteria in plant disease management. In Microbial Endophytes; Woodhead Publishing: Sawston, Cambridge, UK, 2020; pp. 61–89. [Google Scholar] [CrossRef]
- Kannojia, P.; Choudhary, K.K.; Srivastava, A.K.; Singh, A.K. PGPR bioelicitors: Induced systemic resistance (ISR) and proteomic perspective on biocontrol. In PGPR Amelioration in Sustainable Agriculture; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 67–84. [Google Scholar] [CrossRef]
- Bejarano, A.; Puopolo, G. Bioformulation of Microbial Biocontrol Agents for a Sustainable Agriculture. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases. Progress in Biological Control; De Cal, A., Melgarejo, P., Magan, N., Eds.; Springer: Cham, Switzerland, 2020; Volume 21. [Google Scholar] [CrossRef]
- Rahman, S.F.S.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Rändler-Kleine, M.; Wolfgang, A.; Dietel, K.; Junge, H.; Cernava, T.; Berg, G. How microbiome approaches can assist industrial development of biological control products. In Integrative Biological Control; Gao, Y., Hokkanen, H., Menzler-Hokkanen, I., Eds.; Springer: Berlin, Germany, 2020; Volume 20, pp. 201–215. [Google Scholar] [CrossRef]
- Ram, R.M.; Debnath, A.; Negi, S.; Singh, H.B. Use of microbial consortia for broad spectrum protection of plant pathogens: Regulatory hurdles, present status and future prospects. Biopesticides 2022, 2, 319–335. [Google Scholar]
- Minchev, Z.; Kostenko, O.; Soler, R.; Pozo, M.J. Microbial Consortia for Effective Biocontrol of Root and Foliar Diseases in Tomato. Front. Plant Sci. 2021, 12, 756368. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.V.; Cota, F.P.; Longoria, E.C.; Cervantes, J.L.; de los Santos-Villalobos, S. Bacillus subtilis TE3: A promising biological control agent against Bipolaris sorokiniana, the causal agent of spot blotch in wheat (Triticum turgidum L. subsp. durum). Biol. Control 2019, 132, 135–143. [Google Scholar] [CrossRef]
- Aamir, M.; Rai, K.K.; Zehra, A.; Dubey, M.K.; Kumar, S.; Shukla, V.; Upadhyay, R.S. Microbial bioformulation-based plant biostimulants: A plausible approach toward next generation of sustainable agriculture. In Microbial Endophytes; Woodhead Publishing: Sawston, Cambridge, UK, 2020; pp. 195–225. [Google Scholar]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial Interactions Within Multiple-Strain Biological Control Agents Impact Soil-Borne Plant Disease. Front. Microbiol. 2020, 11, 585404. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Muller, D.B. Establishing causality: Opportunities of synthetic communities for plant microbiome research. Cell Host Microbe 2017, 22, 142–155. [Google Scholar] [CrossRef]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef]
- McKellar, M.E.; Nelson, E.B. Compost-induced suppression of Pythium damping-off is mediated by fatty-acid-metabolizing seed-colonizing microbial communities. Appl. Environ. Microbiol. 2003, 69, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wei, Z.; Friman, V.P.; Gu, S.H.; Wang, X.F.; Eisenhauer, N.; Yang, T.J.; Ma, J.; Shen, Q.R.; Xu, Y.C.; et al. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. MBio 2016, 7, e01790-16. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, A.; Saraf, M. Development of microbial consortia as a biocontrol agent for effective management of fungal diseases in Glycine max L. Arch. Phytopathol. Plant Prot. 2014, 48, 459–474. [Google Scholar] [CrossRef]
- Santhanam, R.; Menezes, R.C.; Grabe, V.; Li, D.; Baldwin, I.T.; Groten, K. A suite of complementary biocontrol traits allows a native consortium of root-associated bacteria to protect their host plant from a fungal sudden-wilt disease. Mol. Ecol. 2019, 28, 1154–1169. [Google Scholar] [CrossRef] [PubMed]
- Solanki, M.K.; Yandigeri, M.S.; Kumar, S.; Singh, R.K.; Srivastava, A.K. Co-inoculation of different antagonists can enhance the biocontrol activity against Rhizoctonia solani in tomato. Antonie Van Leeuwenhoek 2019, 112, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ni, M.; Dou, K.; Tang, J.; Ren, J.; Yu, C.; Chen, J. Co-culture of Bacillus amyloliquefaciens ACCC11060 and Trichoderma asperellum GDFS1009 enhanced pathogen-inhibition and amino acid yield. Microb. Cell Factor. 2018, 17, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno- Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A compatible interaction controlling Fusarium wilt of cape gooseberry. Sci. Rep. 2020, 10, 6857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, V.; Kanoujia, N. Biological management of vascular wilt of tomato caused by Fusarium oxysporum f.sp. lycospersici by plant growthpromoting rhizobacterial mixture. Biol. Control 2011, 57, 85–93. [Google Scholar] [CrossRef]
- Kannan, V.; Sureendar, R. Synergistic effect of beneficial rhizosphere microflora in biocontrol and plant growth promotion. J. Basic Microbiol. 2009, 49, 158–164. [Google Scholar] [CrossRef]
- Li, Z.; Bai, X.; Jiao, S.; Li, Y.; Li, P.; Yang, Y.; Zhang, H.; Wei, G. A simplified synthetic community rescues Astragalus mongholicus from root rot disease by activating plant-induced systemic resistance. Microbiome 2021, 9, 217. [Google Scholar] [CrossRef]
- Wong, C.K.F.; Zulperi, D.; Saidi, N.B.; Vadamalai, G. A consortium of Pseudomonas aeruginosa and Trichoderma harzianum for improving growth and induced biochemical changes in Fusarium wilt infected bananas. Tropic. Life Sci. Res. 2021, 32, 23–45. [Google Scholar] [CrossRef]
- Haritha, T.R.; Gopal, K.S. Microbial inoculants and Trichoderma viride consortia for growth promotion and disease management in ginger. J. Spic. Arom. Crops 2021, 30, 58–68. [Google Scholar] [CrossRef]
- Manjula, K.; Podile, A.R. Chitin supplemented formulations improve biocontrol and plant growth-promoting efficiency of Bacillus subtilis AF1. Can. J. Microbiol. 2001, 47, 618–625. [Google Scholar] [CrossRef]
- Vassilev, N.; Reyes, A.; Garcia, M.; Vassileva, M. Production of chitinase by free and immobilized cells of Penicillium janthinellum. A comparison between two biotechnological schemes for inoculant formulation. In Proceedings of the XVIth International Conference on Bioencapsulation, Dublin, Ireland, 4–6 September 2008; pp. 1–4. [Google Scholar]
- Maciag, T.; Krzyzanowska, D.M.; Jafra, S.; Siwinska, J.; Czajkowski, R. The great five-an artificial bacterial consortium with antagonistic activity towards Pectobacterium spp. And Dickeya spp.: Formulation, shelf life, and the ability to prevent soft rot of potato in storage. Appl. Microbiol. Biotechnol. 2020, 104, 4547–4561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.K.F.; Saidi, N.B.; Vadamalai, G.; Teh, C.Y.; Zulperi, D. Effect of bioformulations on the biocontrol efficacy, microbial viability and storage stability of a consortium of biocontrol agents against Fusarium wilt of banana. J. Appl. Microbiol. 2019, 127, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Singh, A.; Singh, S.; Singh, H.B. Biological management of Sclerotinia sclerotiorum in pea using plant growth promoting microbial consortium. J. Bas. Microbiol. 2015, 55, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, Y.; Susilowati, D.N.; Priyatno, T.P.; Samudra, I.M.; Kadir, T.S.; Mubarik, N.R. Prospect of using bacterial bio-formulation to suppress bacterial leaf blight of rice: A case study in Cianjur, West Java. Penelit. Pertan. Tanam. Pangan 2013, 32, 83–90. [Google Scholar]
- Suryadi, Y.; Susilowati, D.; Riana, E.; Mubarik, N.R. Management of rice blast disease (Pyricularia oryzae) using formulated bacterial consortium. Emir. J. Food Agric. 2013, 25, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.N.; Wang, D.C.; Hu, Q.; Dai, X.Q.; Xie, Y.S.; Li, Q.; Liu, H.M.; Guo, J.H. Consortium of plant growth-promoting rhizobacteria strains suppresses sweet pepper disease by altering the rhizosphere microbiota. Front. Microbiol. 2019, 10, 1668. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zheng, L.; Liu, H.X.; Wang, K.B.; Yu, Y.Y.; Luo, Y.M.; Guo, J.H. Evaluation of the effectiveness of a consortium of three plant-growth promoting rhizobacteria for biocontrol of cotton Verticillium wilt. Biocontrol. Sci. Technol. 2014, 24, 489–502. [Google Scholar] [CrossRef]
- Munhoz, L.D.; Fonteque, J.P.; Santos, I.M.O.; Navarro, M.O.P.; Simionato, A.S.; Goya, E.T.; Rezende, M.I.; Balbi-Peña, M.L.; de Oliveira, A.G.; Andrade, G. Control of bacterial stem rot on tomato by extracellular bioactive compounds produced by Pseudomonas aeruginosa LV strain. Cogent Food Agric. 2017, 3, 1282592. [Google Scholar] [CrossRef]
- Liu, K.; Garrett, C.; Fadamiro, H.; Kloepper, J.W. Induction of systemic resistance in Chinese cabbage against black rot by plant growth-promoting rhizobacteria. Biol. Contr. 2016, 99, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Jorjani, M.; Heydari, A.; Zamanizadeh, H.R.; Rezaee, S.; Naraghi, L.; Zamzami, P. Controlling sugar beet mortality disease by application of new bioformulations. J. Plant Protec. Res. 2012, 52, 303–307. [Google Scholar] [CrossRef]
- Lucas, J.A.; Solano, B.R.; Montes, F.; Ojeda, J.; Megias, M.; Manero, F.G. Use of two PGPR strains in the integrated management of blast disease in rice (Oryza sativa) in Southern Spain. Field Crops Res. 2009, 114, 404–410. [Google Scholar] [CrossRef]
- Singh, A.; Sarma, B.K.; Upadhyay, R.S.; Singh, H.B. Compatible rhizosphere Microbes mediated alleviation of biotic stress in chickpea through enhanced antioxidant and phenylpropanoid activities. Microbiol. Res. 2013, 168, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Sudharani, M.; Shivaprakash, M.K.; Prabhavathi, M.K. Role of consortia of biocontrol agents and PGPRs in the production of cabbage under nursery condition. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 1055–1064. [Google Scholar]
- Srinivasan, K.; Mathivanan, N. Plant growth promoting microbial consortia mediated classical biocontrol of sunflower necrosis virus disease. J. Biopest. 2011, 4, 65–72. [Google Scholar]
- Bharathi, R.; Vivekananthan, R.; Harish, S.; Ramanathan, A.; Samiyappan, R. Rhizobacteria-based bio-formulations for the management of fruit rot infection in chillies. Crop Protec. 2004, 23, 835–843. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J.; Bubici, G. Designing a synthetic microbial community devoted to biological control: The case study of Fusarium wilt of banana. Front. Microbiol. 2022, 13, 967885. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Synthetic Consortium/ Microorganisms | Test Plant | Impact on Plant Growth/Yield | Effects on Soil Fertility | Soil Microbiological Activity/ Soil Microbiome | Reference |
|---|---|---|---|---|---|
| Bacillus licheniformis, B. subtilis, B. polymyxa, B. megaterium, B. macerans, P. putida, P. fluorescens, S. cerevisiae, N. orallina, T. viride with Biosolve (humic acid substance) | Blueberry | It increased the shoot and dry weights of plants | It improved the nitrogen and potassium uptake of plants as well as nitrate content in the soil | It changed the composition of the rhizobacterial community in the soil | [143] |
| B. thuringiensis-1312 (BT1) B. thuringiensis-1310 (BT2), B. licheniformis (BL) as consortium of vegetative cells and endospores | Oat | It had positive effects on seed germination and enhanced the total dry biomass of plants | - | It colonized plants’ rhizosphere without modifying the overall structure of microbial communities | [123] |
| SynCom candidates, Arthhrobacter sp., Enterobacter sp., Brevibacterium sp., Plantibacter sp. | Cotton | It increased the germination, plant height, shoot biomass as well as the number of flowers and yield | It enhanced nitrate content and soil nutrient availability by increasing soil fertility | It triggered the enrichment of bacterial members of the phyla Firmicutes, Actinobacteria, and Cyanobacteria in the rhizosphere. However, a shift in fungal communities was observed with the increase in the relative abundance of Basidiomycota and Chytridiomycota | [17] |
| Azotobacter, potassium mobilizing bacteria, zinc solubilizing bacteria, phosphorus solubilizing bacteria, inorganic fertilizers | Wheat | It improved plant growth and increased chlorophyll content was noticed in biofertilizer plus RDF treatments. However, the yield was higher in biofertilizer consortia 2 with RDF | Physical and chemical properties were above critical limits | - | [144] |
| P. fluorescens mvp1–4, P. fluorescens 1m1–96, P. fluorescens Q2–87; P. fluorescens Phl1c2, P. protegens Pf-5, P. protegens CHA0, P. kilonensis F113, P. brassicacearum Q8r1-96 | Tomato | It enhanced plant growth | - | It produced changes in the resident community diversity and composition and an increase in the relative abundance of initially rare taxa. However, the beneficial role of microbial consortium can be indirect on diversity and composition. | [145] |
| S. rhizophila, R. sphaeroides, B. amyloliquefaciens | Oilseed rape | The use of microbes significantly increased the total N content in plants | The application of microbes-maintained soil fertility | It selectively enhanced the growth of Pseudomonadacea and Flavobacteriaceae as well as the recruitment of diazotrophic rhizobacteria such as members of Cyanobacteria and Actinobacteria in the rhizosphere | [146] |
| B. amyloliquefaciens, B. pumilus, B. circulans | Golden kiwi | The application of microbes improved kiwifruit growth | The complex bacterial inoculant was able to increase the availability of N, P, and K contents in soil | - | [147] |
| Enterobacter sp., B. megaterium, B. thuringiensis, Bacillus sp. | French lavender | The combined use with sugar beet residue was the most effective in increasing shoot and root dry biomass | It improved the total N content in the soil | It increased the microbiological and biochemical properties | [148] |
| Anabaena torulosa used as a matrix for agriculturally useful bacteria (Rhizobium, Azotobacter, Pseudomonas, Serratia) | Wheat | It enhanced plant growth with an increase and the nutrient uptake of wheat | Soil fertility was improved | - | [149] |
| T. viride–Bradyrhizobium, T. viride–Azotobacter, T. viride–Bradyrhizobium, Anabaena–T. viride | Mungbean and soybean | The treatment enhanced the fresh and dry weights of plants | It exhibited a high dehydrogenase activity in the soil and nitrogen fixation | The use of microbial treatments enhanced microbial activity in the rhizosphere | [150] |
| B. cereus BT23, Lysobacter capsici ZST1-2, L. antibioticus 13-6 | Chinese cabbage | The use of microbes improved plant yield | It decreased soil acidity | The presence of Bacteroidetes and Proteobacteria was relatively more abundant in rhizosphere and Firmicutes as unique phyla | [151] |
| Microbial consortia product (MCP) (EuroChem Agro GmbH, Mannheim, Germany) | Maize | The plant growth was improved and MCP inoculation stimulated root length development | C, N, and P-turnover in the rhizosphere were slightly affected by MCP inoculation, as deduced from extracellular soil enzymes activities | It increased the abundance of bacteria in the rhizosphere and the auxin production capacity of rhizosphere bacteria | [152] |
| R. irregularis, P. jessenii, P. synxantha | Wheat | This application improved grain yield in wheat plants | It enhanced the dehydrogenase and alkaline phosphatase activities in the soil | Improvement in the PGPR colonization and soil microbiological properties were noted | [153] |
| Microbial Consortium/Bio-Formulations | Disease | Pathogen | Mode or Mechanism of Action | Reference |
|---|---|---|---|---|
| Bacillus megaterium-KAU-PSB, B. sporothermodurans-KAU-KSB, A. lipoferum-KAU-AZO and Bioagents T. viride-KAU-TV and P. fluorescens-KAU-PF | Rhizome rot and leaf blight | R. solani | It reduced the rot and blight incidence in ginger | [180] |
| Enterobacter amnigenus-A167, Serratia plymuthica-A294, S. rubidaea-H440, S. rubidaea-H469, Rahnella aquatilis-H145 | Soft rot disease | Dickeya spp. and Pectobacterium spp. | Potato disease suppression occurred by induction of biosurfactants, siderophores, and antibiotic compounds | [183] |
| T. harzianum-CBF2, P. aeruginosa-DRB1 | Wilt disease | F. oxysporum f. sp. cubense | It induced the production of chitinase and 2,4-diacetylphloroglucinol in banana | [184] |
| T. harzianum-TNHU27, B. subtilis-BHHU100, P. aeruginosa-PJHU15 | White rot | Sclerotinia sclerotiorum | It enhanced oxygen species with induction of systemic resistance in disease management of pea | [185] |
| Formulations of T. harzianum-CBF2 and P. aeruginosa-DRB1 | Wilt disease | F. oxysporum F. sp. cubense (Foc-TR4) | Applied formulations increased the defense response in the host through phenolic and proline contents improvements which in turn reduced root damage in banana | [179] |
| Bacterial community Rhizobium sp., Stenotrophomonas sp., Advenella sp. and Ochrobactrum sp. | Root rot | F. oxysporum | Plants were protected through the synergistic response of highly abundant bacteria with the inhibition of fungal growth. Less abundant bacteria-induced systemic resistance in Astragalus mongholicus | [178] |
| Mixture of B. cereus, B. firmus, P. aeruginosa | Bacterial leaf blight | Xanthomonas oryzae pv. oryzae | The mixture showed a good ability to reduce bacterial blight infection in rice | [186] |
| SynCom1 (P. azotoformans-F30A, T. harzianum-T22, B. amyloliquefaciens-CECT 8238), SynCom2 (P. azotoformans-F30A, B. amyloliquefaciens CECT 8238 and CECT 8237, T. harzianum-22 and ESALQ1306, Pseudomonas chlororaphis-MA 342) | Root and foliar pathogen | F. oxysporum and B. cinerea | Both consortia controlled the pathogens effectively under any of the application schemes through induced systemic resistance and direct antagonism in tomato | [161] |
| B. firmus-E65 C32b, B. cereus II.14, P. aeruginosa, S. marcescens-E31 | Rice blast, sheath, and bacterial leaf blight | Pyricularia oryzae, R. solani, and X. oryzae pv oryzae | Formulations were effective towards leaf and sheath blight while less effect was observed on rice neck blast disease | [187] |
| B. subtilis-SM21, B. cereus-AR156, Serratia sp.-XY21 | Phytophthora blight | Phytophthora capsici | Alternations were observed in the bacterial community of soil in sweet pepper plants | [188] |
| B. cereus-MBAA2, B. amyloliquefaciens-MBAA3, P. aeruginosa-MBAA1 | Charcoal and stem rot | Macrophomina phaseolina and S. sclerotiorum | It induced the production of siderophore, ammonia and beta-1,3 glucanase, cellulose, and chitinase enzymes in soybean | [171] |
| B. subtilis-SM21, B. cereus-AR156, Serratia sp. -XY21 | Wilt disease | Verticillium dahliae | It induced a systematic resistance and secreted antifungal metabolites in cotton plants | [189] |
| P. aeruginosa -LV strain compounds from cell-free supernatant of a bacterial culture. The fraction (F4A) consisted of two main compounds (antibiotic and phenazine-PCN) | Stem rot | Pectobacterium carotovorum subsp. Carotovorum | Elicit systemic acquired resistance (SAR) in tomato | [190] |
| B. velezensis AP136, B. mojavensis AP209, L. macrolides AP282, B. velezensis AP305 | Black rot | X. campestris pv. Campestris | PGPR strain mixtures had the potential to elicit induced systemic resistance challenged with black rot pathogen in cabbage plants | [191] |
| Pseudomonas sp. (PF5, CHA0, Q8R1-96, Q2-87, MVP1-4, 1M1-96, Phl1C227, F113) | Bacterial wilt | Ralstonia solanacearum | It resulted in a competition of resources among bacteria and caused interference with wilt pathogen in tomato | [170] |
| Bioformulations of P. fluorescens and B. coagulans | Seedling damping-off disease | R. solani | A reduction in sugar beet mortality disease was observed | [192] |
| B. cereus-BT-23, Lysobacter antibioticus-13-6, L. capsici-ZST1-2 | Clubroot disease | Plasmodiophora brassicae | Microbial consortia suppressed the disease incidence by recovering the imbalance in the indigenous microbial community composition | [151] |
| P. fluorescens Aur6, Chryseobacterium balustinum Aur9 | Rice blast | Pyricularia oryzae | The disease incidence was reduced by the induction of systemic resistance in rice | [193] |
| P. aeruginosa (PHU094), T. harzianum (THU0816), Rhizobium sp. (RL091) | Collar rot | Sclerotium rolfsii | A disease suppression through antioxidant mechanisms was followed by the activation of phenylpropanoid pathway (PPP) and deposition of lignin in chickpea | [194] |
| Mixture of Azotobacter chroococcum, B. megaterium, P. fluorescens, B. subtilis, T. harzianum | Wilt | Pythium sp. and Fusarium sp. | It showed growth-promoting and disease-suppressing abilities in cabbage | [195] |
| Pantoea vagans-C9-1, P. fluorescens-A506 | Fire blight | E. amylovora | Compatible strain mixtures had greater biological activity suppressing the blight disease in the pear | [107] |
| Bacillus sp., B. licheniformis, S. fradiae, P. aeruginosa | Sunflower Necrosis Virus Disease (SNVD) | Sunflower Necrosis Virus (SNV) | The reductions in virus disease symptoms were associated with a concomitant increase in plant growth and ISR enzymes in sunflower | [196] |
| Pseudomonas community | Bacterial wilt | R. solanacearum | The density of pathogens in the rhizosphere was reduced along with a reduction in the disease incidence because of resource competition and interference with the pathogen in tomato plants | [170] |
| Neem extracts and chitin with P. fluorescens (Pf1) and B. subtilis | Dieback and fruit rot | Colletotrichum capsica | It reduced the fruit rot incidence by the induction of chitinase, peroxidase (POX), β-1,3 glucanase, phenylalanine ammonia-lyase (PAL), polyphenol oxidase (PPO), and accumulation of phenols in chili pepper | [197] |
| Isolates of Bacillus spp., Pseudomonas spp., Streptomyces spp., Trichoderma spp. | Wilt disease | F. oxysporum F. sp. cubense (TR4) | Mixtures of antagonists (synthetic microbial community, SynCom) might provide effective biocontrol against fusarium wilt of banana | [198] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhary, P.; Xu, M.; Ahamad, L.; Chaudhary, A.; Kumar, G.; Adeleke, B.S.; Verma, K.K.; Hu, D.-M.; Širić, I.; Kumar, P.; et al. Application of Synthetic Consortia for Improvement of Soil Fertility, Pollution Remediation, and Agricultural Productivity: A Review. Agronomy 2023, 13, 643. https://doi.org/10.3390/agronomy13030643
Chaudhary P, Xu M, Ahamad L, Chaudhary A, Kumar G, Adeleke BS, Verma KK, Hu D-M, Širić I, Kumar P, et al. Application of Synthetic Consortia for Improvement of Soil Fertility, Pollution Remediation, and Agricultural Productivity: A Review. Agronomy. 2023; 13(3):643. https://doi.org/10.3390/agronomy13030643
Chicago/Turabian StyleChaudhary, Parul, Miao Xu, Lukman Ahamad, Anuj Chaudhary, Govind Kumar, Bartholomew Saanu Adeleke, Krishan K. Verma, Dian-Ming Hu, Ivan Širić, Pankaj Kumar, and et al. 2023. "Application of Synthetic Consortia for Improvement of Soil Fertility, Pollution Remediation, and Agricultural Productivity: A Review" Agronomy 13, no. 3: 643. https://doi.org/10.3390/agronomy13030643