
Opposite Root Morphological Responses of Chinese Cabbage to Poly-γ-glutamic Acid When Applied with Urea and Ammonium Sulphate


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
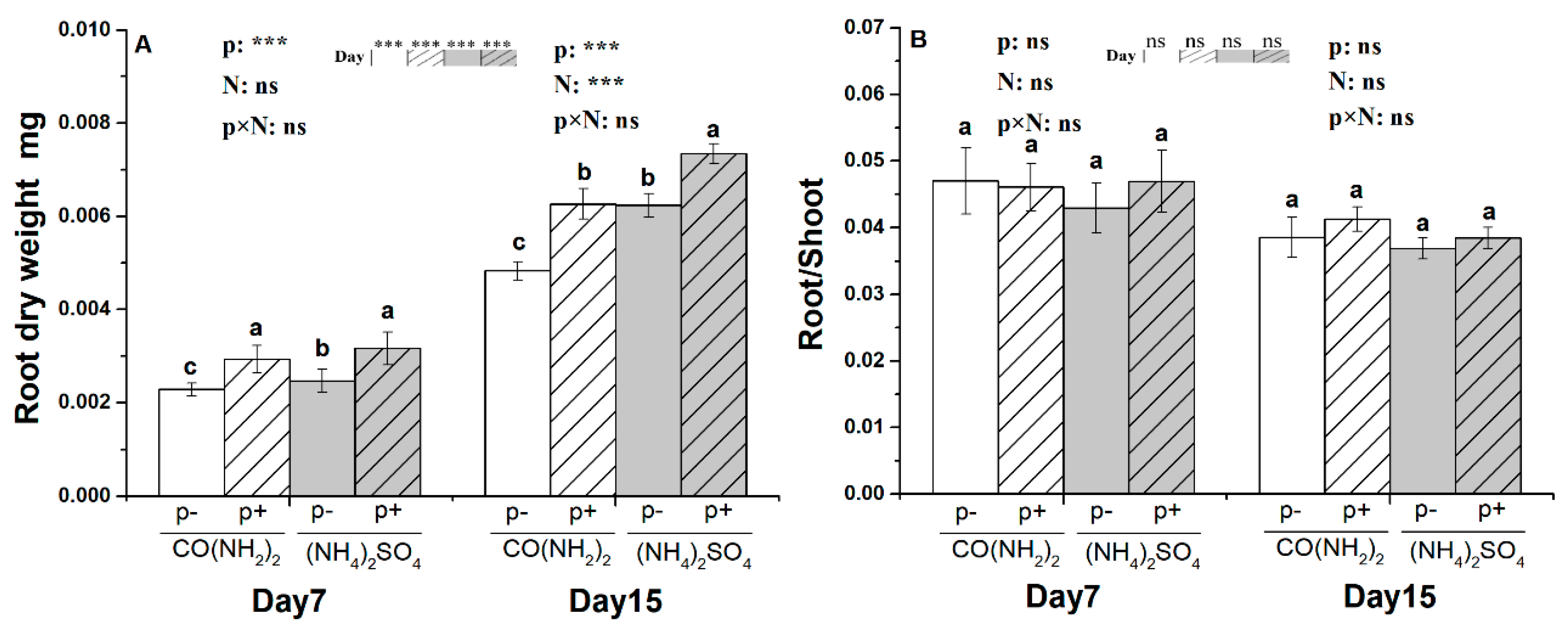
3.1. Root Dry Weight and the Ratio of Root to Shoot under the Effect of γ-PGA
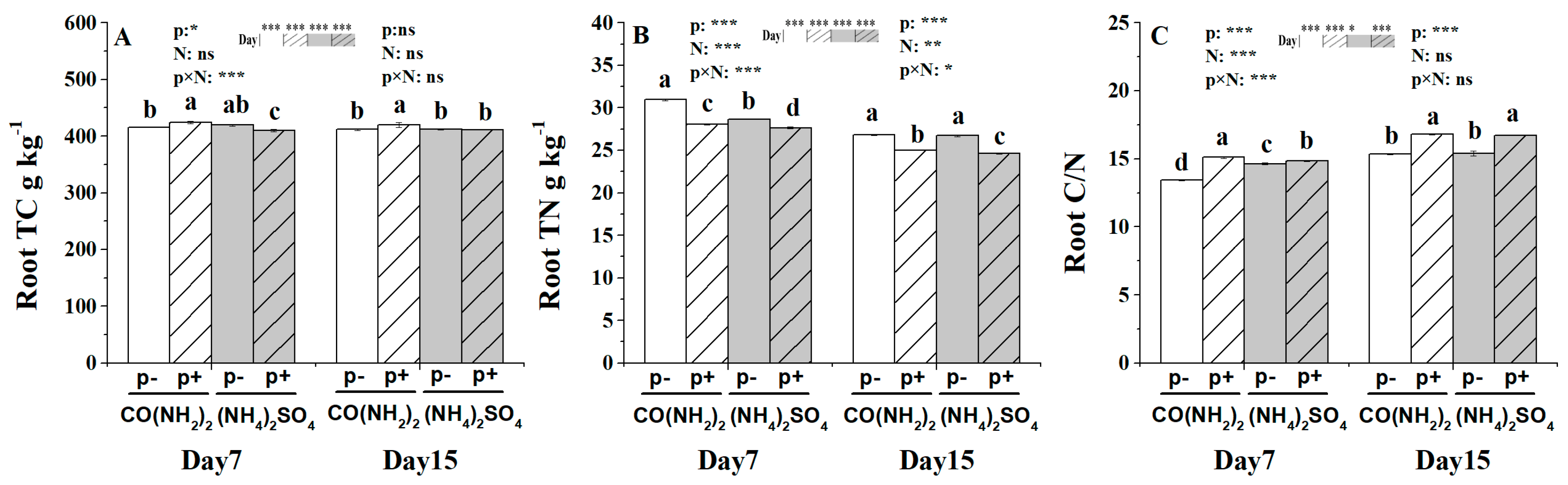
3.2. Root Total Carbon Content, Total Nitrogen Content and the Ratio of Carbon to Nitrogen under the Effect of γ-PGA
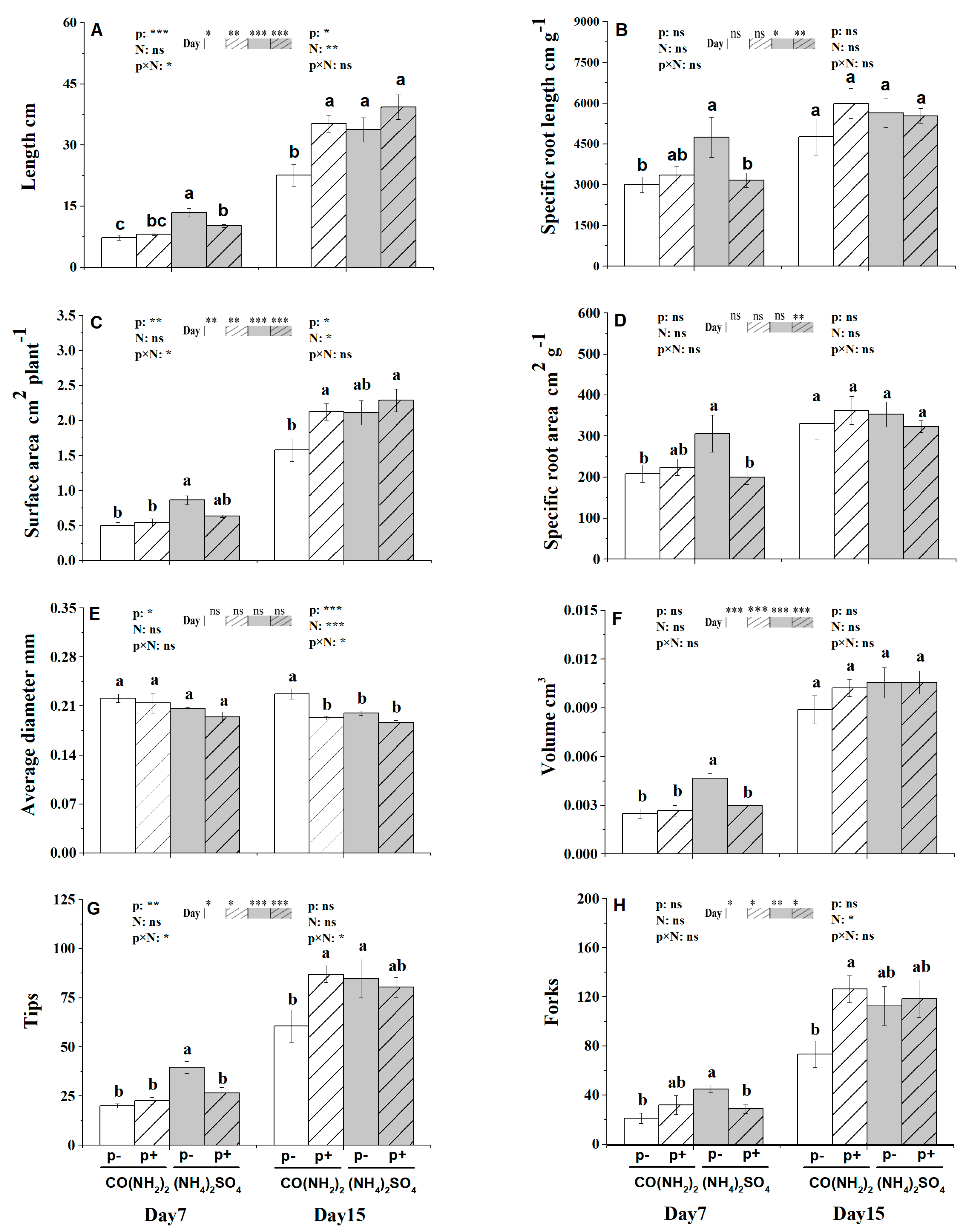
3.3. Root Morphological Traits under the Effect of γ-PGA
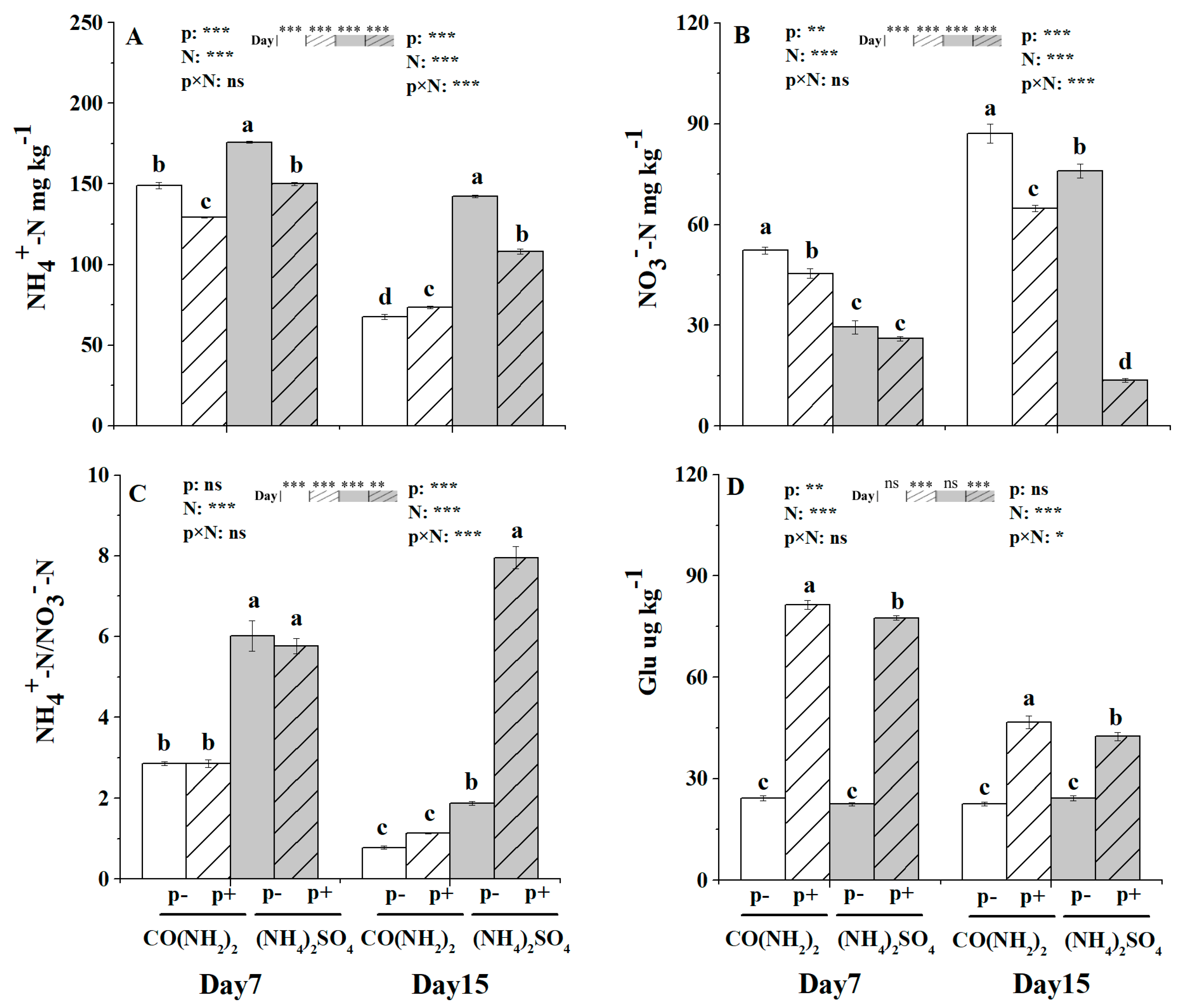
3.4. Soil Available N Content and Glutamic Acid Content under the Effect of γ-PGA
3.5. Relationships between Root Traits and Soil Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, A.J.; Cramer, M.D. Root nitrogen acquisition and assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Garnett, T.; Conn, V.; Kaiser, B.N. Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 2009, 32, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Gao, W.; Cheng, L.; Wang, X.; Duan, F.; Yuan, L.; Rengel, Z.; Zhang, F.; Li, H.; Cahill, J.F.; et al. Harnessing root-foraging capacity to improve nutrient-use efficiency for sustainable maize production. Field Crops Res. 2022, 279, 108462. [Google Scholar] [CrossRef]
- Zhang, H.; Rong, H.; Pilbeam, D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 2329–2338. [Google Scholar] [CrossRef] [Green Version]
- Mi, G.; Chen, F.; Wu, Q.; Lai, N.; Yuan, L.; Zhang, F. Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems. Sci. China Life Sci. 2010, 53, 1369–1373. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrogen signalling pathways shaping root system architecture: An update. Curr. Opin. Plant Biol. 2014, 21, 30–36. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; von Wiren, N. Root Nutrient Foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Love, J.; Roche, J.; Song, J.; Turnbull, M.H.; Jameson, P.E. A RootNav analysis of morphological changes in Brassica napus L. roots in response to different nitrogen forms. Plant Growth Regul. 2017, 83, 83–92. [Google Scholar] [CrossRef]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wirén, N. Plasticity of the Arabidopsis Root System under Nutrient Deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.E.; Kojima, S.; Takahashi, H.; von Wiren, N. Ammonium triggers lateral root branching in Arabidopsis in an AMMONIUM TRANSPORTER1;3-dependent manner. Plant Cell. 2010, 22, 3621–3633. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Gao, J.; Liu, Y.; Tian, Z.; Muhammad, A.; Zhang, Y.; Jiang, D.; Cao, W.; Dai, T. Higher Ammonium Transamination Capacity Can Alleviate Glutamate Inhibition on Winter Wheat (Triticum aestivum L. ) Root Growth under High Ammonium Stress. PLoS One 2016, 11, e0160997. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied. Soil Biol. Biochem. 2010, 42, 2336–2338. [Google Scholar] [CrossRef]
- Jing, J.; Zhang, F.; Rengel, Z.; Shen, J. Localized fertilization with P plus N elicits an ammonium-dependent enhancement of maize root growth and nutrient uptake. Field Crops Res. 2012, 133, 176–185. [Google Scholar] [CrossRef]
- Schwambach, J.; Ruedell, C.M.; de Almeida, M.R.; Fett-Neto, A.G. Nitrogen Sources and Adventitious Root Development in Eucalyptus globulus Microcuttings. J. Plant Nutr. 2015, 38, 1628–1638. [Google Scholar] [CrossRef]
- Walch-Liu, P.; Liu, L.H.; Remans, T.; Tester, M.; Forde, B.G. Evidence that L-glutamate can act as an exogenous signal to modulate root growth and branching in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Walch-Liu, P.; Forde, B.G. Nitrate signalling mediated by the NRT1.1 nitrate transporter antagonises l-glutamate-induced changes in root architecture. Plant J. 2008, 54, 820–828. [Google Scholar] [CrossRef]
- Parati, M.; Khalil, I.; Tchuenbou-Magaia, F.; Adamus, G.; Mendrek, B.; Hill, R.; Radecka, I. Building a circular economy around poly(D/L-gamma-glutamic acid)- a smart microbial biopolymer. Biotechnol. Adv. 2022, 61, 108049. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Yu, Z. Degradability of poly-γ-glutamic acid in environment. Environ. Sci. Technol. 2008, 31, 35–37, 88. [Google Scholar]
- Zhang, L.; Wei, Z.; Wang, L.; Sun, Y.; Pei, J.; Wang, J.; Gao, J.; Zhang, L.; Shi, Y. Fate of Urea and Ammonium Sulfate in the Plant and Soil System as Affected by Poly-gamma-glutamic Acid. J. Soil Sci. Plant Nut. 2022, 22, 2457–2468. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Gao, D.; Wang, L.; Li, J.; Wei, Z.; Shi, Y. Effects of poly-γ-glutamic acid (γ-PGA) on plant growth and its distribution in a controlled plant-soil system. Sci. Rep. 2017, 7, 6090. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Lei, P.; Feng, X.; Xu, X.; Xu, H.; Yang, H.; Tang, W. Effect of poly(γ-glutamic acid) on microbial community and nitrogen pools of soil. Acta Agr. Scand. 2013, 63, 657–668. [Google Scholar] [CrossRef]
- Xu, Z.; Wan, C.; Xu, X.; Feng, X.; Xu, H. Effect of poly (γ-glutamic acid) on wheat productivity, nitrogen use efficiency and soil microbes. J. Soil Sci. Plant Nutr. 2013, 13, 744–755. [Google Scholar] [CrossRef]
- Yin, A.; Jia, Y.; Qiu, T.; Gao, M.; Cheng, S.; Wang, X.; Sun, Y. Poly-γ-glutamic acid improves the drought resistance of maize seedlings by adjusting the soil moisture and microbial community structure. Appl. Soil Ecol. 2018, 129, 128–135. [Google Scholar] [CrossRef]
- IUSS Working group WRB. World Reference Base for soil resources 2014, update 2015. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports (2015) No. 106; FAO: Rome, Italy.
- Cai, H.; Chen, F.; Mi, G.; Zhang, F.; Maurer, H.P.; Liu, W.; Reif, J.C.; Yuan, L. Mapping QTLs for root system architecture of maize (Zea mays L.) in the field at different developmental stages. Theor. Appl. Genet. 2012, 125, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Costs and benefits of constructing roots of small diameter. J. Plant Nutr. 1992, 15, 763–782. [Google Scholar] [CrossRef]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root Development and Nutrient Uptake. Crit. Rev. Plant Sci. 2007, 25, 279–301. [Google Scholar] [CrossRef]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B.; Bardgett, R.D.; De Deyn, G.B.; Johnson, D.; Klimesova, J.; et al. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytol. 2021, 232, 1123–1158. [Google Scholar] [CrossRef]
- Tang, L.; Sun, H.; Sun, R.; Niu, Y.; Song, J.; Li, S.; Shen, Y. Optimized Nitrogen Application Increases Soil Water Extraction by Changing in-Season Maize Root Morphology and Distribution in Rainfed Farmland. Agronomy 2020, 10, 1606. [Google Scholar] [CrossRef]
- Jia, Z.; von Wiren, N. Signaling pathways underlying nitrogen-dependent changes in root system architecture: From model to crop species. J. Exp. Bot. 2020, 71, 4393–4404. [Google Scholar] [CrossRef]
- Ke, J.; Pu, W.-X.; Wang, H.; Liu, L.-H.; Sheng, S. Phenotypical evidence of effective amelioration of ammonium-inhibited plant (root) growth by exogenous low urea. J. Plant Physiol. 2020, 255, 153306. [Google Scholar] [CrossRef]
- Liu, L.; Bi, X.Y.; Sheng, S.; Gong, Y.Y.; Pu, W.-X.; Ke, J.; Huang, P.J.; Liang, Y.L.; Liu, L.H. Evidence that exogenous urea acts as a potent cue to alleviate ammonium-inhibition of root system growth of cotton plant (Gossypium hirsutum). Physiol. Plantarum 2021, 171, 137–150. [Google Scholar] [CrossRef]
- Peng, Y.; Yu, P.; Zhang, Y.; Sun, G.; Ning, P.; Li, X.; Li, C. Temporal and spatial dynamics in root length density of field-grown maize and NPK in the soil profile. Field Crops Res. 2012, 131, 9–16. [Google Scholar] [CrossRef]
- Liu, Y.X.; Zhang, W.P.; Sun, J.H.; Li, X.F.; Christie, P.; Li, L. High morphological and physiological plasticity of wheat roots is conducive to higher competitive ability of wheat than maize in intercropping systems. Plant. Soil. 2015, 397, 387–399. [Google Scholar] [CrossRef]
- Ning, P.; Li, S.; White, P.J.; Li, C. Maize Varieties Released in Different Eras Have Similar Root Length Density Distributions in the Soil, Which Are Negatively Correlated with Local Concentrations of Soil Mineral Nitrogen. PLoS ONE 2015, 10, e0121892. [Google Scholar] [CrossRef]
- Mehrabi, F.; Sepaskhah, A.R.; Ahmadi, S.H. Winter wheat root distribution with irrigation, planting methods, and nitrogen application. Nutr. Cycl. Agroecosystems 2021, 119, 231–245. [Google Scholar] [CrossRef]
- Chen, W.; Koide, R.T.; Eissenstat, D.M. Nutrient foraging by mycorrhizas: From species functional traits to ecosystem processes. Funct. Ecol. 2018, 32, 858–869. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Li, H.; Shen, Q.; Tang, X.; Xiong, C.; Li, H.; Pang, J.; Ryan, M.H.; Lambers, H.; Shen, J. Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species. New Phytol. 2019, 223, 882–895. [Google Scholar] [CrossRef]
- Wen, Z.; White, P.J.; Shen, J.; Lambers, H. Linking root exudation to belowground economic traits for resource acquisition. New Phytol. 2022, 233, 1620–1635. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RDW | RS | TN | TC | CN | NH4+-N | NO3−-N | NH4+-N/NO3−-N | Glu | RL | SRL | RA | SRA | Diameter | Volume | Tip | Fork | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RDW | 1 | ||||||||||||||||
| RS | −0.38 | 1 | |||||||||||||||
| TN | −0.440 * | 0.14 | 1 | ||||||||||||||
| TC | −0.889 ** | 0.502 * | 0.342 | 1 | |||||||||||||
| C/N | 0.807 ** | −0.491 * | −0.019 | −0.919 ** | 1 | ||||||||||||
| NH4+-N | −0.650 ** | 0.234 | 0.086 | 0.695 ** | −0.807 ** | 1 | |||||||||||
| NO3−-N | 0.095 | −0.155 | 0.038 | −0.044 | 0.182 | −0.435 * | 1 | ||||||||||
| NH4+-N/NO3−-N | −0.095 | 0.131 | −0.126 | 0.122 | −0.295 | 0.549 * | −0.961 ** | 1 | |||||||||
| Glu | 0.056 | 0.202 | 0.178 | −0.163 | 0.206 | −0.097 | −0.38 | 0.217 | 1 | ||||||||
| RL | 0.911 ** | −0.560 * | −0.311 | −0.897 ** | 0.876 ** | −0.694 ** | 0.17 | −0.197 | 0.122 | 1 | |||||||
| SRL | 0.581 * | −0.489 * | −0.07 | −0.651 ** | 0.705 ** | −0.550 * | 0.23 | −0.287 | 0.079 | 0.767 ** | 1 | ||||||
| RA | 0.889 ** | −0.507 * | −0.307 | −0.878 ** | 0.854 ** | −0.724 ** | 0.222 | −0.246 | 0.067 | 0.970 ** | 0.815 ** | 1 | |||||
| SRA | 0.561 * | −0.379 | 0.006 | −0.609 * | 0.693 ** | −0.613 ** | 0.328 | −0.376 | 0.058 | 0.737 ** | 0.957 ** | 0.811 ** | 1 | ||||
| Diameter | −0.412 * | 0.363 | 0.214 | 0.459 * | −0.368 | −0.05 | 0.322 | −0.286 | −0.281 | −0.465 * | −0.289 | −0.311 | −0.08 | 1 | |||
| Volume | 0.735 ** | −0.497 * | −0.073 | −0.781 ** | 0.855 ** | −0.794 ** | 0.415 * | −0.464 * | 0.037 | 0.855 ** | 0.794 ** | 0.855 ** | 0.830 ** | −0.192 | 1 | ||
| Tips | 0.818 ** | −0.439 * | −0.147 | −0.830 ** | 0.883 ** | −0.776 ** | 0.419 * | −0.466 * | 0.046 | 0.877 ** | 0.730 ** | 0.903 ** | 0.755 ** | −0.222 | 0.855 ** | 1 | |
| Forks | 0.858 ** | −0.581 * | −0.144 | −0.836 ** | 0.864 ** | −0.654 ** | 0.269 | −0.272 | 0.031 | 0.938 ** | 0.768 ** | 0.948 ** | 0.776 ** | −0.304 | 0.849 ** | 0.913 ** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Wang, L.; Sun, Y.; Dong, X.; Wei, Z.; Zhang, L.; Shi, Y. Opposite Root Morphological Responses of Chinese Cabbage to Poly-γ-glutamic Acid When Applied with Urea and Ammonium Sulphate. Agronomy 2023, 13, 608. https://doi.org/10.3390/agronomy13020608
Zhang L, Wang L, Sun Y, Dong X, Wei Z, Zhang L, Shi Y. Opposite Root Morphological Responses of Chinese Cabbage to Poly-γ-glutamic Acid When Applied with Urea and Ammonium Sulphate. Agronomy. 2023; 13(2):608. https://doi.org/10.3390/agronomy13020608
Chicago/Turabian StyleZhang, Lei, Lingli Wang, Yu Sun, Xiaoyan Dong, Zhanbo Wei, Lili Zhang, and Yuanliang Shi. 2023. "Opposite Root Morphological Responses of Chinese Cabbage to Poly-γ-glutamic Acid When Applied with Urea and Ammonium Sulphate" Agronomy 13, no. 2: 608. https://doi.org/10.3390/agronomy13020608