
Rapid Detection of A282S Mutation in the RDL1 Gene of Rice Stem Borer via the Mutation-Specific LAMP Technique

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. suppressalis Population
2.2. Plasmid-Construction of CsRDL1 with A282S Mutation
2.3. Design of Specific Primers for LAMP Reaction
2.4. Optimization of LAMP Reaction Component
2.5. Optimization of LAMP Reaction Parameters
2.6. Verifying the Accuracy of the LAMP Technique
3. Results
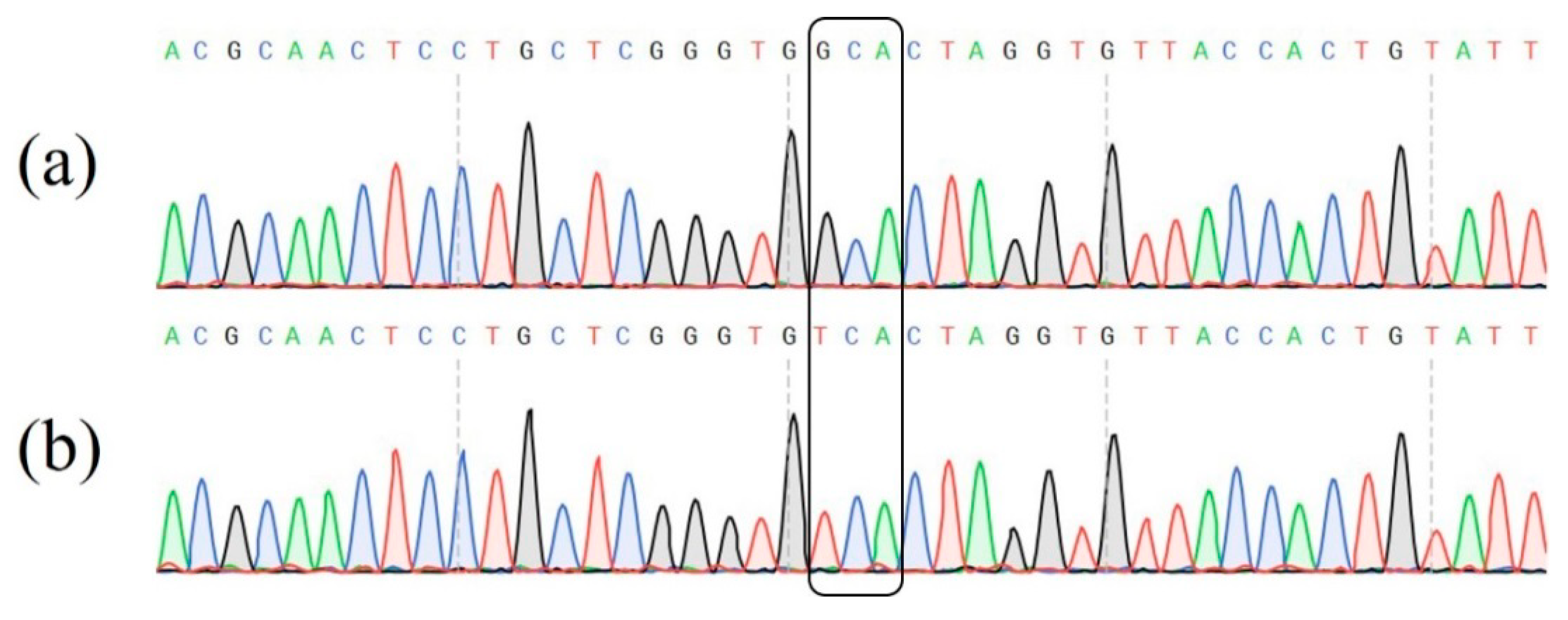
3.1. Mutation of the CsRDL1
3.2. Design Specific Primer forLAMP Reaction
3.3. Optimized Components of LAMP Reaction System
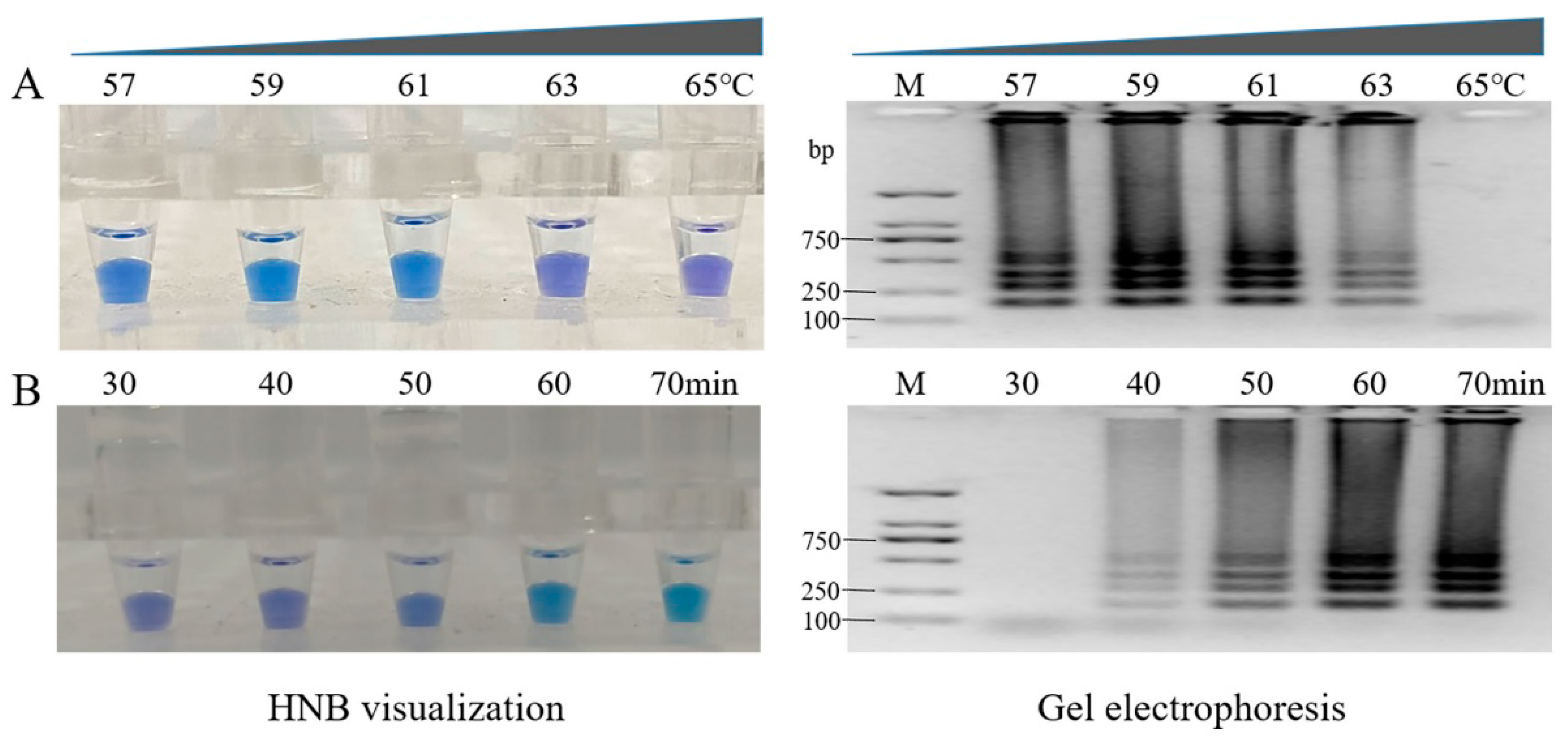
3.4. Optimization of Reaction Parameters of the LAMP Technique
3.5. Accuracy of the LAMP Technique
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sial, M.U.; Zhao, Z.Z.; Zhang, L.; Zhang, Y.N.; Mao, L.G.; Jiang, H.Y. Loop-mediated isothermal amplification for the detection of R81T mutation in nAChR with crude genomic DNA extracted from individual Myzus persicae. J. Pest Sci. 2020, 93, 531–541. [Google Scholar] [CrossRef]
- Huang, Q.C.; Deng, Y.F.; Zhan, T.S.; He, Y.A. Synergistic and antagonistic effects of piperonyl butoxide in fipronil-susceptible and resistant rice stem borrers, Chilo suppressalis. J. Insect Sci. 2010, 10, 182. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.K.; Wu, Z.L.; Jiang, C.Y.; Guan, D.J.; Zhang, N.; Jiang, H.; Shen, Q.W.; Qian, K.; Wang, J.J. Identification and characterization of glutathione S-transferases and their potential roles in detoxification of abamectin in the rice stem borer, Chilo suppressalis. Pestic. Biochem. Physiol. 2022, 182, 105050. [Google Scholar] [CrossRef]
- Wang, X.; Wu, S.; Gao, W.; Wu, Y. Dominant inheritance of field-evolved resistance to fipronil in Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2016, 109, 334–338. [Google Scholar] [CrossRef]
- Wolff, M.A.; Wingate, V.P.M. Characterization and comparative pharmacological studies of a functional γ-aminobutyric acid (GABA) receptor cloned from the tobacco budworm, Heliothis virescens (Noctuidae: Lepidoptera). Invertebr. Neurosci. 1998, 3, 305–315. [Google Scholar] [CrossRef]
- Garrood, W.T.; Zimmer, C.T.; Gutbrod, O.; Lüke, B.; Williamson, M.S.; Bass, C.; Nauen, R.; Emyr Davies, T.G. Influence of the RDL A301S mutation in the brown planthopper Nilaparvata lugens on the activity of phenylpyrazole insecticides. Pestic. Biochem. Physiol. 2017, 142, 1–8. [Google Scholar] [CrossRef]
- Francois, P.; Tangomo, M.; Hibbs, J.; Bonetti, E.; Boehme, C.; Notomi, T.; Perkins, M.; Schrenzel, J. Robustness of a loop-mediated isothermal amplification reaction for diagnostic applications. FEMS Immunol. Med. Microbiol. 2011, 62, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Ge, C.Y.; Zhang, X.K.; Wang, J.X.; Zhou, M.G. Development and evaluation of a novel and rapid detection assay for Botrytis cinerea based on loop-mediated isothermal amplification. PLoS ONE 2014, 9, e111094. [Google Scholar] [CrossRef]
- Moradi, A.; Almasi, M.A.; Jafary, H.; Mercado-Blanco, J. A novel and rapid loop-mediated isothermal amplification assay for the specific detection of Verticillium dahliae. J. Appl. Microbiol. 2014, 116, 942–954. [Google Scholar] [CrossRef]
- Kitagawa, M.; Nagamine, K.; Oka, H.; Ouhara, K.; Ogawa, I.; Komatsuzawa, H.; Kurihara, H. Rapid detection of the Streptococcus mutans cnm gene by loop-mediated isothermal amplification. Anal. Biochem. 2020, 605, 113812. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soroka, M.; Wasowicz, B.; Rymaszewska, A. Loop-mediated isothermal amplification (LAMP): The better sibling of PCR? Cells 2021, 10, 1931. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Z.; Lowe, S.B.; Gooding, J.J. Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP). Biosens. Bioelectron. 2014, 61, 491–499. [Google Scholar] [CrossRef]
- Badolo, A.; Bando, H.; Traore, A.; Ko-Ketsu, M.; Guelbeogo, W.M.; Kanuka, H.; Ranson, H.; Sagnon, N.; Fukumoto, S. Detection of G119S ace-1R mutation in field-collected Anopheles gambiae mosquitoes using allele-specific loop-mediated isothermal amplification (AS-LAMP) method. Malar. J. 2015, 14, 477. [Google Scholar] [CrossRef] [Green Version]
- Uchibori-Asano, M.; Uchiyama, T.; Jouraku, A.; Shinoda, T. Development of allele-specific loop-mediated isothermal amplification (AS-LAMP) to detect the tebufenozide-resistant allele in the smaller tea tortrix, Adoxophyes honmai (Lepidoptera: Tortricidae). Appl. Entomol. Zool. 2022, 57, 93–99. [Google Scholar] [CrossRef]
- Li, B.; Han, L.Z.; Peng, Y.F. Development of a standardized artificial diet and rearing technique for the striped stem borer, Chilo suppressalis Walker (Lepidoptera: Crambidae). Chin. J. Entomol. 2015, 52, 498–503. [Google Scholar] [CrossRef]
- Tomita, N.; Mori, Y.; Kanda, H.; Notomi, T. Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nat. Protoc. 2008, 3, 877–882. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Rocheleau, T.A.; Steichen, J.C.; Chalmers, A.E. A point mutation in a Drosophila GABA receptor confers insecticide resistance. Nature 1993, 363, 449–451. [Google Scholar] [CrossRef]
- Nakao, T.; Naoi, A.; Hama, M.; Kawahara, N.; Hirase, K. Concentration-dependent effects of GABA on insensitivity to fipronil in the A2′S mutant RDL GABA receptor from fipronil-resistant Oulema oryzae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2012, 105, 1781–1788. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes. 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K.-I. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. BioTechniques 2009, 46, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Coates, B.S.; Chen, H.; Sappington, T.W.; Guillemaud, T.; Siegfried, B.D. Role of a gamma-aminobutryic acid (GABA) receptor mutation in the evolution and spread of Diabrotica virgifera virgifera resistance to cyclodiene insecticides. Insect Mol. Biol. 2013, 22, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Anthony, N.M.; Brown, J.K.; Markham, P.G.; Ffrench-Constant, R.H. Molecular analysis of cyclodiene resistance-associated mutations among populations of the sweetpotato whitefly Bemisia tabaci. Pestic. Biochem. Physiol. 1995, 51, 220–228. [Google Scholar] [CrossRef]
- Hansen, K.K.; Kristensen, M.; Jensen, K.M.V. Correlation of a resistance-associated Rdl mutation in the German cockroach, Blattella germanica (L), with persistent dieldrin resistance in two Danish field populations. Pest Manag. Sci. 2005, 61, 749–753. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′-3′) | Function |
|---|---|---|
| CsRDL1-F | CACACGCTAACACGACAT | amplification of CsRDL1 gene |
| CsRDL1-R | CACGATGCGAGAATACTTG | |
| CsRDL1A282S-F | ACTCCTGCTCGGGTGTCACTAGGTGT | mutagenesis of A282S site |
| CsRDL1A282S-R | ACACCCGAGCAGGAGTTGCGTTTCGG | |
| CsRDL1-F3 | CGAAACAGGTGCCCAAAT | outer primer for LAMP |
| CsRDL1-B3 | CATCCAAATTTATATTCCATCTGG | |
| CsRDL1-FIP | AGACCCGAGCAGGAGTTGCTGATTGTAATCATATCATGGGTATC | inner primer for LAMP |
| CsRDL1-BIP | GTTACCACTGTATTGACTATGACTACGTCGATAGATTTCACGTAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Z.; Zhang, S.; Man, W.; Li, Y.; Luo, G.; Yu, J.; Fu, Y.; Yao, J.; Chen, Y.; Zhao, C. Rapid Detection of A282S Mutation in the RDL1 Gene of Rice Stem Borer via the Mutation-Specific LAMP Technique. Agronomy 2023, 13, 601. https://doi.org/10.3390/agronomy13020601
Song Z, Zhang S, Man W, Li Y, Luo G, Yu J, Fu Y, Yao J, Chen Y, Zhao C. Rapid Detection of A282S Mutation in the RDL1 Gene of Rice Stem Borer via the Mutation-Specific LAMP Technique. Agronomy. 2023; 13(2):601. https://doi.org/10.3390/agronomy13020601
Chicago/Turabian StyleSong, Zijiao, Sugui Zhang, Wentao Man, Yao Li, Guanghua Luo, Jinyao Yu, Yilan Fu, Junxi Yao, Yiqu Chen, and Chunqing Zhao. 2023. "Rapid Detection of A282S Mutation in the RDL1 Gene of Rice Stem Borer via the Mutation-Specific LAMP Technique" Agronomy 13, no. 2: 601. https://doi.org/10.3390/agronomy13020601