
Evaluating, Screening and Selecting Yardlong Bean [Vigna unguiculata subsp. sesquipedalis (L.) Verdc.] for Resistance to Common Cutworm (Spodoptera litura Fabricius)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Common Cutworm Rearing
2.3. Investigation on the Damage of the Common Cutworm to the Yardlong Bean in the Greenhouse
2.4. Evaluation of Yardlong Bean Varieties for Resistance to the Common Cutworm
2.5. Selection of Yardlong Bean Germplasm Resources Resistant to Common Cutworm
2.6. Determination of Crude Protein Content and Starch Content in Yardlong Bean Leaves
2.7. Resistance for Common Cutworm of Different Organs of Yardlong Bean
2.8. Data Analysis
3. Results
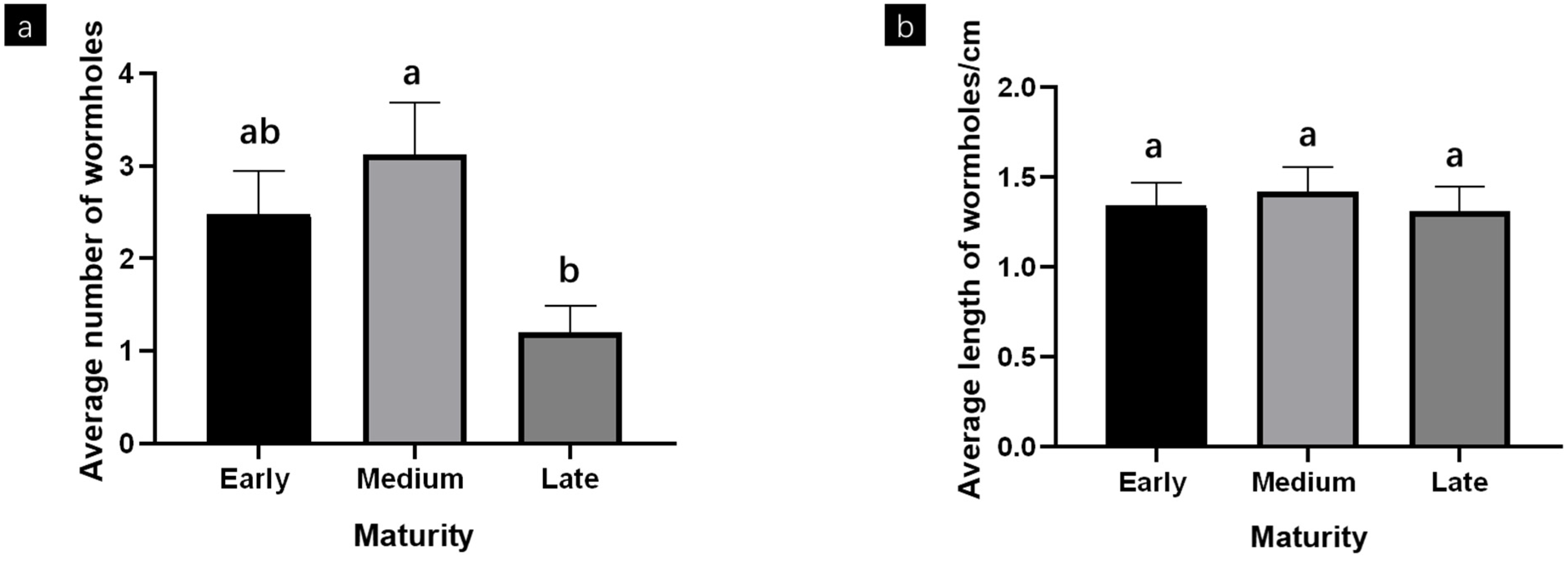
3.1. Common Cutworm Damage to the Yardlong Bean in a Greenhouse
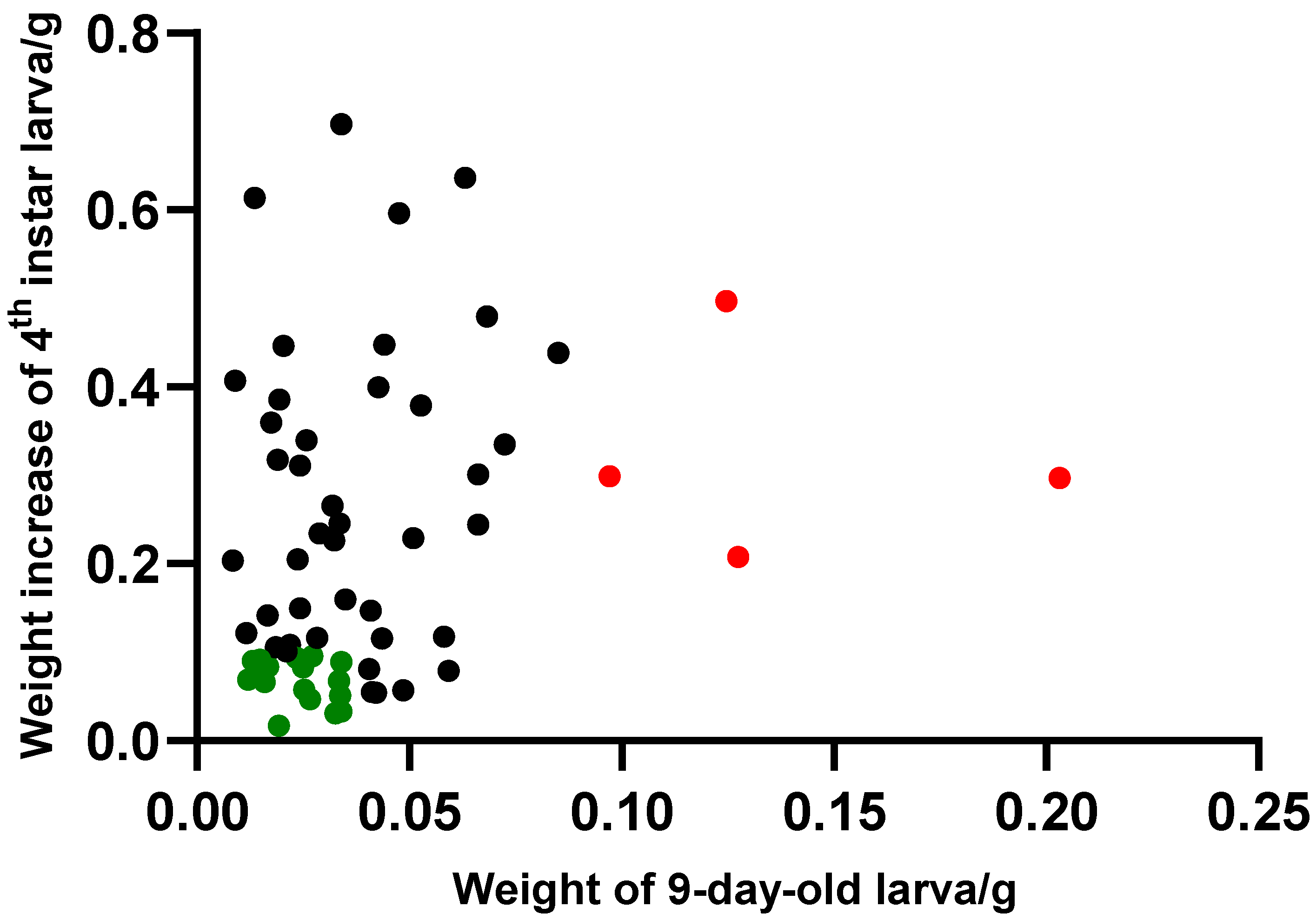
3.2. Evaluation of Yardlong Bean Varieties for Resistance to the Common Cutworm
3.3. Selection of Yardlong Bean Germplasm Resources Resistant to Common Cutworm
3.4. Determination of Crude Protein Content and Starch Content in Yardlong Bean Leaves
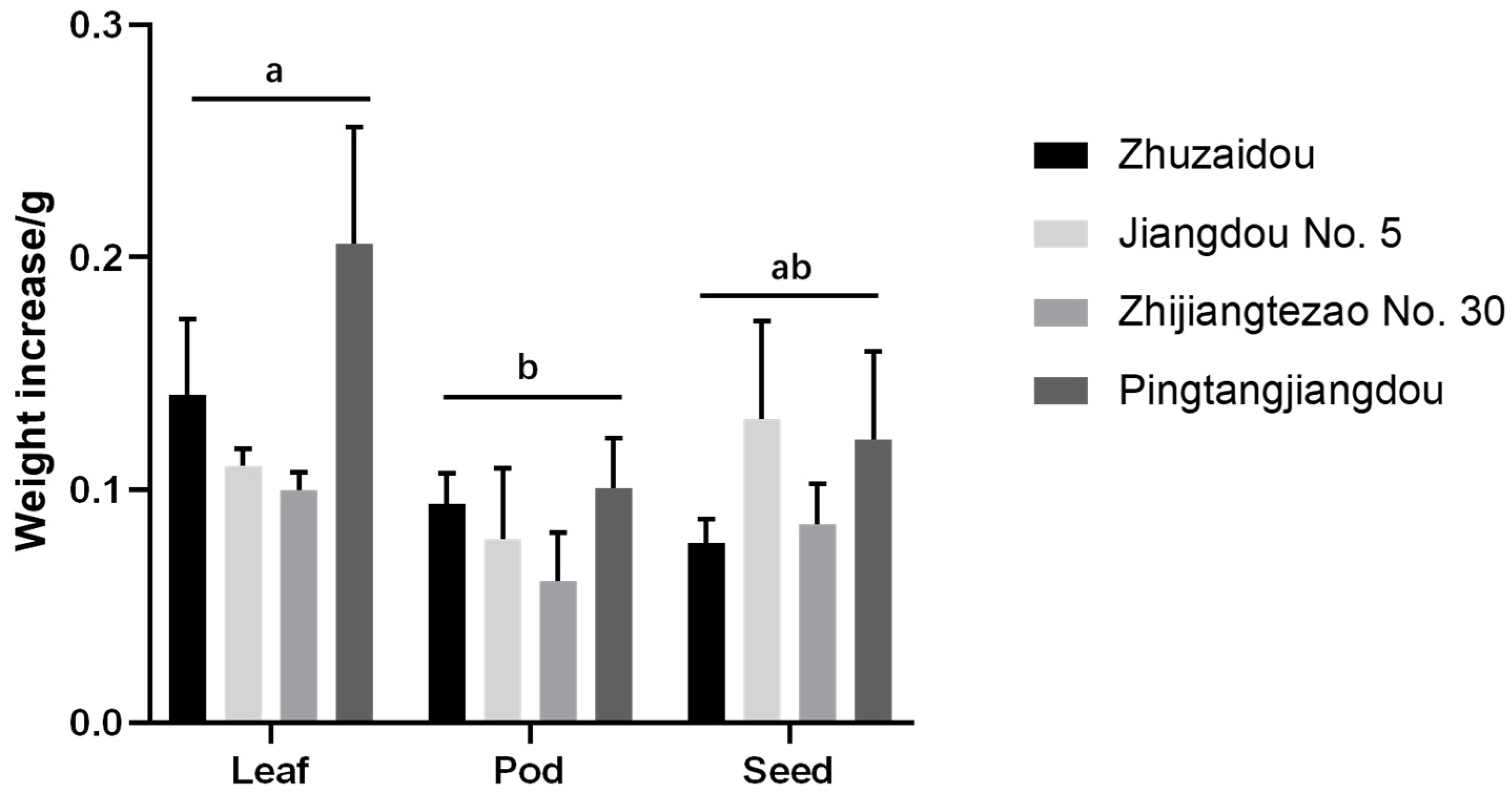
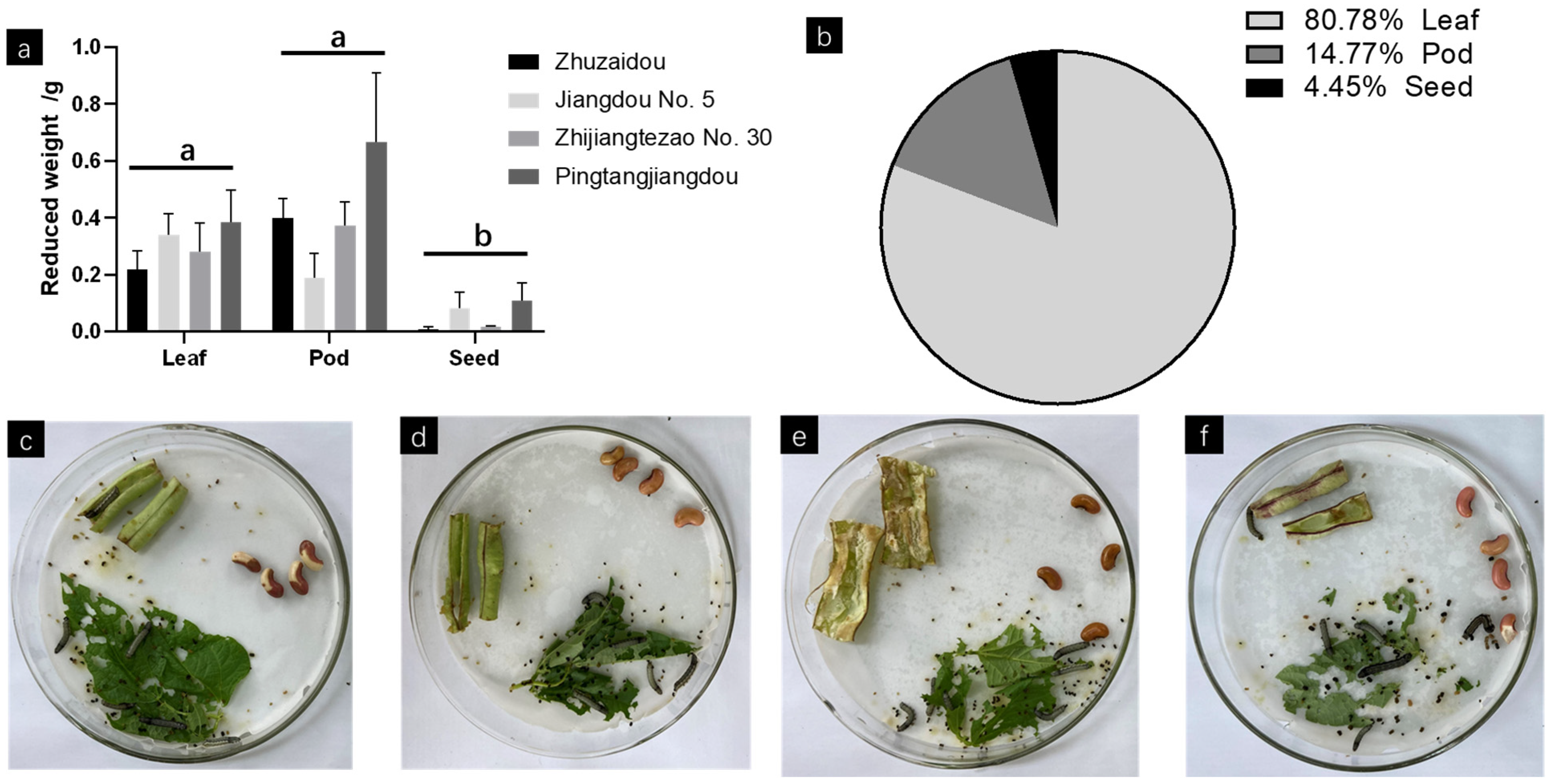
3.5. Antibiotic and Antixenotic Experiments in Different Organs of Yardlong Beans
4. Discussion
4.1. Insect Resistance Evaluation Method of Yardlong Bean
4.2. Reasons for Antibiosis
4.3. Meaning of Experiments and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, L.; Li, Y.; Yu, X.; Li, J.; Chen, C. Research progress of molecular genetics in cowpea (Vigna unguiculata). J. Changjiang Veg. 2014, 24, 1–13. [Google Scholar] [CrossRef]
- Smýkal, P.; Coyne, C.; Ambrose, M.; Maxted, N.; Schaefer, H.; Blair, M.; Berger, J.; Greene, S.; Nelson, M.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–103. [Google Scholar] [CrossRef]
- Wang, S. Origin classification and genetic resources of cowpea. China Veg. 1989, 6, 49–52. [Google Scholar]
- Wang, B.; Wu, X.; Li, S.; Chen, X.; Li, Y.; Wang, Y.; Lu, Z.; Wu, X.; Li, G. Evaluation of cowpea germplasm accessions collected from Zhejiang province. J. Plant Genet. Resour. 2021, 2, 380–389. [Google Scholar] [CrossRef]
- Coker, C.; Ely, M.; Freeman, T. Evaluation of yardlong bean as a potential new crop for growers in the Southeastern United States. HortTechnology 2007, 17, 592–594. [Google Scholar] [CrossRef]
- Phillips, R.D.; McWatters, K.H.; Chinnan, M.S.; Hung, Y.-C.; Beuchat, L.R.; Sefa-Dedeh, S.; Sakyi-Dawson, E.; Ngoddy, P.; Nnanyelugo, D.; Enwere, J.; et al. Utilization of cowpeas for human food. Field Crop. Res. 2003, 82, 193–213. [Google Scholar] [CrossRef]
- Rabé, M.M.; Baoua, I.B.; Baributsa, D. Farmers’ referred genotype traits and socio-economic factors influencing the adoption of improved cowpea varieties in South-Central Niger. Agronomy 2022, 12, 2668. [Google Scholar] [CrossRef]
- Bell, L.W.; James, A.T.; Augustin, M.A.; Rombenso, A.; Blyth, D.; Simon, C.; Higgins, T.J.V.; Barrero, J.M. A niche for cowpea in sub-tropical Australia? Agronomy 2021, 11, 1654. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Deng, Q. Inheritance and breeding of important traits in cowpea. China Veg. 1992, 1, 50–53. [Google Scholar]
- Jackai, L.E.N. Integrated pest management of borers of cowpea and beans. Int. J. Trop. Insect Sci. 1995, 16, 237–250. [Google Scholar] [CrossRef]
- Pan, Y.F.; Luo, F.; Lei, C. The niche of important pests and natural enemies in the cowpea field ecosystem. Chin. J. Appl. Entomol. 2005, 42, 404–408. [Google Scholar]
- Nyarko, J.; Asare, A.T.; Mensah, B.A.; Adjei, F. Assessment of the response of fifteen cowpea [Vigna unguiculata L. (Walp.)] genotypes to infestation by Callosobruchus maculatus Fab. (Coloeptera: Bruchidae). Cogent Food Agric. 2022, 8, 2095713. [Google Scholar] [CrossRef]
- Huang, K.; Wen, L.; Zhang, X. Investigation on population dynamics of Prodenia litura for cowpea and its control measures. Anhui Agric. Sci. Bull. 2008, 9, 163. [Google Scholar] [CrossRef]
- Painter, R.H. Insect resistance in crop plants. Soil Sci. 1951, 72, 481. [Google Scholar] [CrossRef]
- Roy, D.; Chakraborty, G.; Biswas, A.; Sarkar, P.K. Antixenosis, tolerance and genetic analysis of some rice landraces for resistance to Nilaparvata lugens (Stål.). J. Asia Pac. Entomol. 2021, 24, 448–460. [Google Scholar] [CrossRef]
- Smith, C.M. Antibiosis: Adverse effects of resistance on arthropod biology. In Plant Resistance to Arthropods: Molecular and Conventional Approaches; Smith, C.M., Ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 65–99. [Google Scholar]
- Eduardo, W.I.; Júnior, A.L.B.; Moraes, R.F.D.O.; de Souza, B.H.S.; Louvandini, H.; Barbosa, J.C. Protocol for assessing soybean antixenosis to Heliothis virescens. Èntomol. Exp. Appl. 2020, 168, 911–927. [Google Scholar] [CrossRef]
- Bruna, M.F.; Alessandra, R.B.; Luís, A.F. Biology and reproductive capacity of Spodoptera eridania (Cramer) (Lepidoptera, Noctuidae) in different soybean cultivars. Rev. Bras. Entomol. 2015, 59, 89–95. [Google Scholar] [CrossRef]
- Yang, Y.; Xing, G.; Gai, J. Evaluation of antibiosis to common cutworm (Spodoptera litura) and screening for resistance sources among wild soybeans (Glycine soja) in China. Soybean Sci. 2016, 35, 448–454. [Google Scholar]
- Hu, Z.; Xu, X.; Pan, L.; Li, M.; Zeng, J.; Razzaq, M.K.; Xing, G.; Gai, J. Resistance analyses of soybean organs to common cutworm (Spodoptera litura) at different reproductive stages. Soybean Sci. 2020, 39, 932–939. [Google Scholar]
- Xu, H.; Zhai, H.; Wang, F.; Piao, J.; Yang, X.; Zhu, Z. Cowpea trypsin inhibitor gene (cpti) and its application in insect resistance transgenic plants. J. Agric. Sci. Technol. 2008, 10, 18–27. [Google Scholar]
- Ma, X.; Zhu, Z.; Li, Y.; Yang, G.; Pei, Y. Expressing a modified cowpea trypsin inhibitor gene to increase insect tolerance against Pieris rapae in Chinese cabbage. Hortic. Environ. Biotechnol. 2017, 58, 195–202. [Google Scholar] [CrossRef]
- Zhou, X.; Pan, Y.; Wang, X.; Li, H.; Lei, C. Effects of food-plants on the survival, development and fecundity of Maruca vitrata (Fabricius). J. Huazhong Agric. Univ. 2008, 3, 367–369. [Google Scholar]
- Costa, E.N.; Evangelista, B.M.D.; Fernandes, M. Antibiosis levels to Spodoptera frugiperda (Lepidoptera: Noctuidae) in cowpea commercial cultivars and landrace varieties. J. Econ. Èntomol. 2019, 112, 1941–1945. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Pan, L.; Wan, H.; Guo, R.; Chen, G.; Xie, Q.; Ren, S.; Lu, S.; Yan, M.; Chen, C. Germplasm innovation of pest-resistant asparagus bean by interspecific crosses. Acta Hortic. Sin. 2020, 47, 562–570. [Google Scholar] [CrossRef]
- Lu, W.; Wei, Y.; Tan, D.; Zuo, F. The enlightenment of the poisonous cowpea incident in Hainan to plant protection in Guangxi. J. Guangxi Agric. 2010, 25, 86–87, 96. [Google Scholar]
- Enders, L.; Begcy, K. Unconventional routes to developing insect-resistant crops. Mol. Plant 2021, 14, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Eker, T.; Erler, F.; Sari, H.; Sari, D.; Berger, J.; Toker, C. Deployment of Cicer echinospermum P.H. Davis for resistance to Callosobruchus chinensis L. J. Plant Dis. Prot. 2022, 129, 843–851. [Google Scholar] [CrossRef]
- Tu, Y.; Zeng, J. A method for artificial rearing of common cutworm (Spodoptera litura). Acta Agric. Jiangxi 2010, 22, 87–88. [Google Scholar]
- Laurentin, A.; Edwards, C.A. A microtiter modification of the anthrone-sulfuric acid colorimetric assay for glucose-based carbohydrates. Anal. Biochem. 2003, 315, 143–145. [Google Scholar] [CrossRef]
- Zhao, F.; Qian, J.; Liu, H.; Wang, C.; Wang, X.; Wu, W.; Wang, D.; Cai, C.; Lin, Y. Quantification, identification and comparison of oligopeptides on five tea categories with different fermentation degree by Kjeldahl method and ultra-high performance liquid chromatography coupled with quadrupole-orbitrap ultra-high resolution mass spectrometry. Food Chem. 2022, 378, 132130. [Google Scholar] [CrossRef]
- Xing, G.; Zhou, B.; Wang, Y.; Zhao, T.; Yu, D.; Chen, S.; Gai, J. Genetic components and major QTL confer resistance to bean pyralid (Lamprosema indicata Fabricius) under multiple environments in four RIL populations of soybean. Theor. Appl. Genet. 2012, 125, 859–875. [Google Scholar] [CrossRef]
- Cong, S.; Xu, D.; Yang, N.; Wang, L.; Wang, J.; Liu, W.; Wan, P. Effects of stacked genetically modified cotton on the feeding behavior and nutrient metabolism of Spodoptera litura larvae. Plant Prot. 2022, 48, 104–110. [Google Scholar] [CrossRef]
- Yao, W. Biology characteristies of Prodenia litura. J. Biosaf. 2005, 2, 122–127. [Google Scholar]
- Xing, G.; Liu, K.; Gai, J. A high-throughput phenotyping procedure for evaluation of antixenosis against common cutworm at early seedling stage in soybean. Plant Methods 2017, 13, 66. [Google Scholar] [CrossRef]
- Yang, J.; Xiao, G. The insect-resistance physiology of plants: A review. Chin. Agric. Sci. Bull. 2021, 37, 130–136. [Google Scholar] [CrossRef]
- Zhao, J.; Segar, S.T.; McKey, D.; Chen, J. Macroevolution of defense syndromes in Ficus (Moraceae). Ecol. Monogr. 2021, 91, e01428. [Google Scholar] [CrossRef]
- Zeng, W.; Cai, Z.; Zhang, Z.; Chen, H.; Yang, S.; Tang, X.; Lai, Z.; Sun, Z. Physiological and biochemical characteristics of Lamprosema indicate (Fabricius)-resistant soybean. J. S. Agric. 2015, 46, 2112–2116. [Google Scholar] [CrossRef]
- Gaquerel, E.; Stitz, M. Insect resistance: An emerging molecular framework linking plant age and JA ignaling. Mol. Plant 2017, 10, 537–539. [Google Scholar] [CrossRef]
- Diezel, C.; Allmann, S.; Baldwin, I.T. Mechanisms of optimal defense patterns in Nicotiana attenuata: Flowering attenuates herbivory-elicited ethylene and jasmonate signaling. J. Integr. Plant Biol. 2011, 53, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Malook, S.U.; Qi, J.; Hettenhausen, C.; Xu, Y.; Zhang, C.; Zhang, J.; Lu, C.; Li, J.; Wang, L.; Wu, J. The oriental armyworm (Mythimna separata) feeding induces systemic defence responses within and between maize leaves. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180307. [Google Scholar] [CrossRef] [Green Version]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Luo, S.; Yi, Q.; Li, C.; Luo, Q.; Hua, J.; Liu, Y.; Li, S. Secondary metabolites from Glycine soja and their growth inhibitory effect against Spodoptera litura. J. Agric. Food Chem. 2011, 59, 6004–6010. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Ullah, F.; Hafeez, M.; Han, X.; Dara, M.Z.N.; Gul, H.; Zhao, C.R. Biological control of fall armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2704. [Google Scholar] [CrossRef]
- Koch, K.G.; Chapman, K.; Louis, J.; Heng-Moss, T.; Sarath, G. Plant tolerance: Aunique approach to control hemipteran pests. Front. Plant Sci. 2016, 7, 1363. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.R.; Roy, M.C.; Rahman, M.M.; Miah, M.G.; Kwon, Y.J.; Suh, S.J. Foraging and growth responses of cotton armyworm Spodoptera litura to the biophysical characteristics of five cotton varieties. Entomol. Res. 2015, 45, 286–293. [Google Scholar] [CrossRef]
- Gomes, A.M.F.; Draper, D.; Nhantumbo, N.; Massinga, R.; Ramalho, J.C.; Marques, I.; Ribeiro-Barros, A.I. Diversity of cowpea [Vigna unguiculata (L.) Walp] landraces in mozambique: New opportunities for crop improvement and future breeding programs. Agronomy 2021, 11, 991. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, W.; Chen, H.; Chen, J.; Liu, X.; Chen, X.; Yang, S. Transcriptomic analysis of salt tolerance-associated genes and diversity analysis using indel markers in yardlong bean (Vigna unguiculata ssp. sesquipedialis). BMC Genom. Data 2021, 22, 34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties Code | Resistance Type | Variety | Origin | 4thLW/g | LW9/g |
|---|---|---|---|---|---|
| 1 | sensitive | Changtingbaopihongjiangdou | Changting, Fujian | 0.497 | 0.125 |
| 2 | sensitive | Zhuzaidou | Guangzhou, Guangdong | 0.297 | 0.203 |
| 3 | sensitive | Suijiao101 | Guangzhou, Guangdong | 0.301 | 0.066 |
| 4 | sensitive | 156fanjiang | Wuhan, Hubei | 0.299 | 0.097 |
| S-MEAN | 0.349 | 0.123 | |||
| 5 | resistant | Baijiangdou | Danzhai, Guizhou | 0.067 | 0.016 |
| 6 | resistant | Changcaidou | Zanhuang, Hebei | 0.067 | 0.033 |
| 7 | resistant | Jiangdou No.5 | Wuhan, Hubei | 0.031 | 0.033 |
| 8 | resistant | Zhijiangtezao No.30 | Wuhan, Hubei | 0.017 | 0.019 |
| 9 | resistant | SS-97 | Nanjing, Jiangsu | 0.095 | 0.027 |
| 10 | resistant | Suzi41 | Nanjing, Jiangsu | 0.051 | 0.034 |
| 11 | resistant | Sujiang1419 | Nanjing, Jiangsu | 0.033 | 0.034 |
| 12 | resistant | j-5 | Nanjing, Jiangsu | 0.057 | 0.025 |
| 13 | resistant | Sujiang12 | Nanjing, Jiangsu | 0.083 | 0.017 |
| 14 | resistant | Dabaitiaojiangdou | Shenyang, Liaoning | 0.077 | 0.015 |
| 15 | resistant | Paojiangdou-2 | Yaan, Sichuan | 0.069 | 0.012 |
| 16 | resistant | Gaochan No.4 | Shantou, Guangdong | 0.090 | 0.013 |
| 17 | resistant | Pingtangjiangdou | Pingtang, Guizhou | 0.047 | 0.027 |
| 18 | resistant | j-1 | Jiangsu Academy of Agricultural Sciences | 0.091 | 0.015 |
| 19 | resistant | Feicui | Guangdong Academy of Agricultural Sciences | 0.089 | 0.034 |
| 20 | resistant | Sujiang No.1 | Jiangsu Academy of Agricultural Sciences | 0.094 | 0.023 |
| 21 | resistant | Jiangdou2045 | Wuhan, Hubei | 0.081 | 0.025 |
| R-MEAN | 0.067 | 0.024 |
| Source of Variation | LW6 | LW9 | LW12 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DF | F | p (>F) | DF | F | p (>F) | DF | F | p (>F) | |
| Variety | 20 | 5.64 | <0.0001 | 20 | 1.46 | 0.0992 | 20 | 1.92 | 0.014 |
| Batch | 2 | 315.40 | <0.0001 | 2 | 22.97 | <0.0001 | 2 | 32.55 | <0.0001 |
| Variety × Batch | 40 | 5.74 | <0.0001 | 40 | 0.47 | 0.9947 | 40 | 0.85 | 0.7264 |
| Block | 2 | 1.89 | 0.1550 | 2 | 1.55 | 0.2163 | 2 | 2.92 | 0.0581 |
| Error | 119 | 110 | 108 | ||||||
| Indicator | Variety | Block | Error | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DF | MS | F | p (>F) | DF | MS | F | p (>F) | MS | |
| LW6-1 | 20 | 0.00003 | 3.87 | 0.0001 | 2 | 0.00007 | 10.32 | 0.0003 | 0.00001 |
| LW6-2 | 20 | 0.00017 | 9.51 | <0.0001 | 2 | 0.00002 | 1.04 | 0.3629 | 0.00002 |
| LW6-3 | 20 | 0.00005 | 1.24 | 0.2723 | 2 | 0.00037 | 9.85 | 0.0003 | 0.00004 |
| LW9-1 | 20 | 0.00985 | 11.11 | <0.0001 | 2 | 0.00224 | 2.53 | 0.0930 | 0.00089 |
| LW9-2 | 20 | 0.03681 | 19.23 | <0.0001 | 2 | 0.00760 | 3.97 | 0.0276 | 0.00191 |
| LW9-3 | 20 | 0.00305 | 0.92 | 0.5680 | 2 | 0.00690 | 2.08 | 0.1379 | 0.00332 |
| LW12-1 | 20 | 0.05880 | 10.49 | <0.0001 | 2 | 0.01310 | 2.34 | 0.1104 | 0.00560 |
| LW12-2 | 20 | 0.13736 | 10.18 | <0.0001 | 2 | 0.00749 | 0.56 | 0.5789 | 0.01350 |
| LW12-3 | 20 | 0.06898 | 2.30 | 0.0122 | 2 | 0.09932 | 3.32 | 0.0465 | 1.19794 |
| Correlation | LW6-1 | LW6-2 | LW6-3 | LW9-1 | LW9-2 | LW9-3 | LW12-1 | LW12-2 | LW12-3 | Starch |
|---|---|---|---|---|---|---|---|---|---|---|
| LW6-2 | 0.41 | |||||||||
| LW 6-3 | 0.02 | −0.19 | ||||||||
| LW 9-1 | 0.67 ** | 0.65 ** | −0.14 | |||||||
| LW 9-2 | 0.34 | 0.81 ** | −0.11 | 0.55 * | ||||||
| LW 9-3 | 0.24 | 0.18 | 0.65 ** | 0.33 | 0.27 | |||||
| LW 12-1 | 0.60 ** | 0.61 ** | −0.11 | 0.95 ** | 0.59 ** | 0.37 | ||||
| LW 12-2 | 0.30 | 0.76 ** | −0.21 | 0.54 * | 0.95 ** | 0.18 | 0.59 ** | |||
| LW 12-3 | 0.40 | 0.19 | 0.18 | 0.58 ** | 0.29 | 0.69 ** | 0.64 ** | 0.23 | ||
| Starch | 0.09 | 0.14 | 0.14 | −0.27 | −0.04 | −0.02 | −0.28 | −0.03 | −0.47 * | |
| Crude protein | −0.15 | 0.10 | −0.24 | 0.16 | 0.29 | 0.12 | 0.24 | 0.32 | 0.20 | −0.26 |
| Varieties | LW6-1 /g | LW6-2 /g | LW6-3 /g | LW9-1 /g | LW9-2 /g | LW9-3 /g | LW12-1 /g | LW12-2 /g | LW12-3 /g | Starch mg/g | Crude Protein g/kg |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Changtingbaopihongjiangdou | 0.006 | 0.014 | 0.031 | 0.107 | 0.337 | 0.238 | 0.260 | 0.690 | 0.730 | 55.6 | 325 |
| Zhuzaidou | 0.007 | 0.015 | 0.020 | 0.168 | 0.274 | 0.219 | 0.430 | 0.740 | 0.600 | 52.3 | 308 |
| Suijiao101 | 0.013 | 0.006 | 0.029 | 0.152 | 0.145 | 0.188 | 0.340 | 0.450 | 0.480 | 72.1 | 304 |
| 156fanjiang | 0.010 | 0.016 | 0.019 | 0.112 | 0.240 | 0.214 | 0.240 | 0.510 | 0.710 | 66.8 | 316 |
| S-MEAN | 0.009 | 0.013 | 0.025 | 0.135 | 0.249 | 0.215 | 0.318 | 0.598 | 0.630 | 61.7 | 313 |
| Baijiangdou | 0.006 | 0.021 | 0.021 | 0.167 | 0.282 | 0.220 | 0.390 | 0.640 | 0.580 | 55.0 | 385 |
| Changcaidou | 0.003 | 0.002 | 0.013 | 0.036 | 0.098 | 0.109 | 0.060 | 0.280 | 0.180 | 54.8 | 317 |
| Jiangdou No.5 | 0.004 | 0.001 | 0.020 | 0.021 | 0.046 | 0.147 | 0.050 | 0.190 | 0.260 | 58.3 | 321 |
| Zhijiangtezao No.30 | 0.005 | 0.002 | 0.028 | 0.018 | 0.049 | 0.207 | 0.020 | 0.220 | 0.380 | 81.4 | 326 |
| SS-97 | 0.005 | 0.003 | 0.022 | 0.039 | 0.082 | 0.198 | 0.110 | 0.290 | 0.450 | 93.3 | 288 |
| Suzi41 | 0.011 | 0.024 | 0.028 | 0.096 | 0.259 | 0.184 | 0.230 | 0.520 | 0.270 | 99.3 | 266 |
| Sujiang1419 | 0.010 | 0.015 | 0.020 | 0.159 | 0.350 | 0.213 | 0.390 | 0.680 | 0.600 | 60.9 | 342 |
| j-5 | 0.003 | 0.009 | 0.021 | 0.010 | 0.170 | 0.159 | 0.010 | 0.410 | 0.330 | 75.2 | 331 |
| Sujiang12 | 0.003 | 0.004 | 0.031 | 0.062 | 0.155 | 0.242 | 0.290 | 0.420 | 0.560 | 74.0 | 340 |
| Dabaitiaojiangdou | 0.004 | 0.011 | 0.023 | 0.051 | 0.253 | 0.185 | 0.110 | 0.760 | 0.380 | . | . |
| Paojiangdou-2 | 0.005 | 0.012 | 0.026 | 0.069 | 0.157 | 0.236 | 0.170 | 0.370 | 0.510 | 82.7 | 327 |
| Gaochan No.4 | 0.007 | 0.002 | 0.037 | 0.091 | 0.101 | 0.281 | 0.180 | 0.190 | 0.620 | 57.1 | 310 |
| Pingtangjiangdou | 0.007 | 0.022 | 0.023 | 0.162 | 0.414 | 0.239 | 0.430 | 0.830 | 0.720 | 52.0 | 313 |
| j-1 | 0.002 | 0.001 | 0.034 | 0.010 | 0.041 | 0.202 | 0.020 | 0.110 | 0.460 | 47.9 | 278 |
| Feicui | 0.009 | 0.004 | 0.026 | 0.098 | 0.064 | 0.261 | 0.210 | 0.210 | 0.740 | 59.7 | 304 |
| Sujiang No.1 | 0.008 | 0.001 | 0.020 | 0.104 | 0.042 | 0.175 | 0.310 | 0.200 | 0.810 | 38.0 | 321 |
| Jiangdou2045 | 0.006 | 0.017 | 0.020 | 0.183 | 0.080 | 0.180 | 0.360 | 0.290 | 0.530 | 63.2 | 295 |
| R-MEAN | 0.006 | 0.009 | 0.024 | 0.081 | 0.155 | 0.202 | 0.196 | 0.389 | 0.493 | 65.8 | 317 |
| Source of Variation | DF | SS | MS | F | p (>F) |
|---|---|---|---|---|---|
| Variety | 3 | 0.0171 | 0.0057 | 2.46 | 0.0868 |
| Organ | 2 | 0.0192 | 0.0096 | 4.16 | 0.0281 |
| Variety × Organ | 6 | 0.0125 | 0.0021 | 0.90 | 0.5107 |
| Error | 24 | 0.0554 | 0.0023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, T.; Xu, Y.; Jiang, H.; Chen, X.; Liu, X.; Chen, H.; Zhang, H.; Xing, G. Evaluating, Screening and Selecting Yardlong Bean [Vigna unguiculata subsp. sesquipedalis (L.) Verdc.] for Resistance to Common Cutworm (Spodoptera litura Fabricius). Agronomy 2023, 13, 502. https://doi.org/10.3390/agronomy13020502
Yao T, Xu Y, Jiang H, Chen X, Liu X, Chen H, Zhang H, Xing G. Evaluating, Screening and Selecting Yardlong Bean [Vigna unguiculata subsp. sesquipedalis (L.) Verdc.] for Resistance to Common Cutworm (Spodoptera litura Fabricius). Agronomy. 2023; 13(2):502. https://doi.org/10.3390/agronomy13020502
Chicago/Turabian StyleYao, Tianchi, Yufei Xu, Huiyan Jiang, Xinxin Chen, Xiaofeng Liu, Huatao Chen, Hongmei Zhang, and Guangnan Xing. 2023. "Evaluating, Screening and Selecting Yardlong Bean [Vigna unguiculata subsp. sesquipedalis (L.) Verdc.] for Resistance to Common Cutworm (Spodoptera litura Fabricius)" Agronomy 13, no. 2: 502. https://doi.org/10.3390/agronomy13020502