
Foliar Application of Carnosine and Chitosan Improving Drought Tolerance in Bermudagrass

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1—Screening the Optimal Concentration of Carnosine and Chitosan Affecting Plant Growth
2.2. Experiment 2—Effects of Exogenous Carnosine or Chitosan on Drought Tolerance in Bermudagrass
2.3. Determination of Growth and Physiological Indexes
2.4. Measurements of Antioxidant Enzyme Activity, Malondialdehyde, and Hydrogenperoxide Content
2.5. Statistical Analysis
3. Results
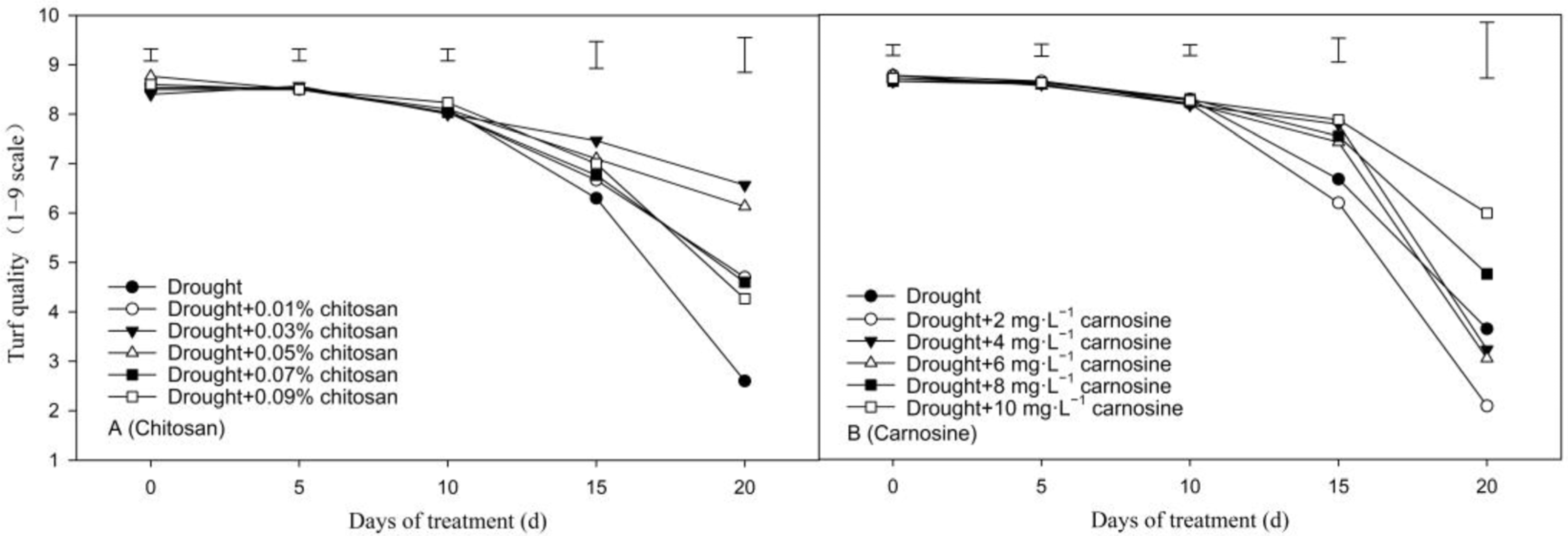
3.1. Determination of the Optimal Concentration of Carnosine and Chitosan
3.2. Effects of Carnosine and Chitosan on Soil Water Status
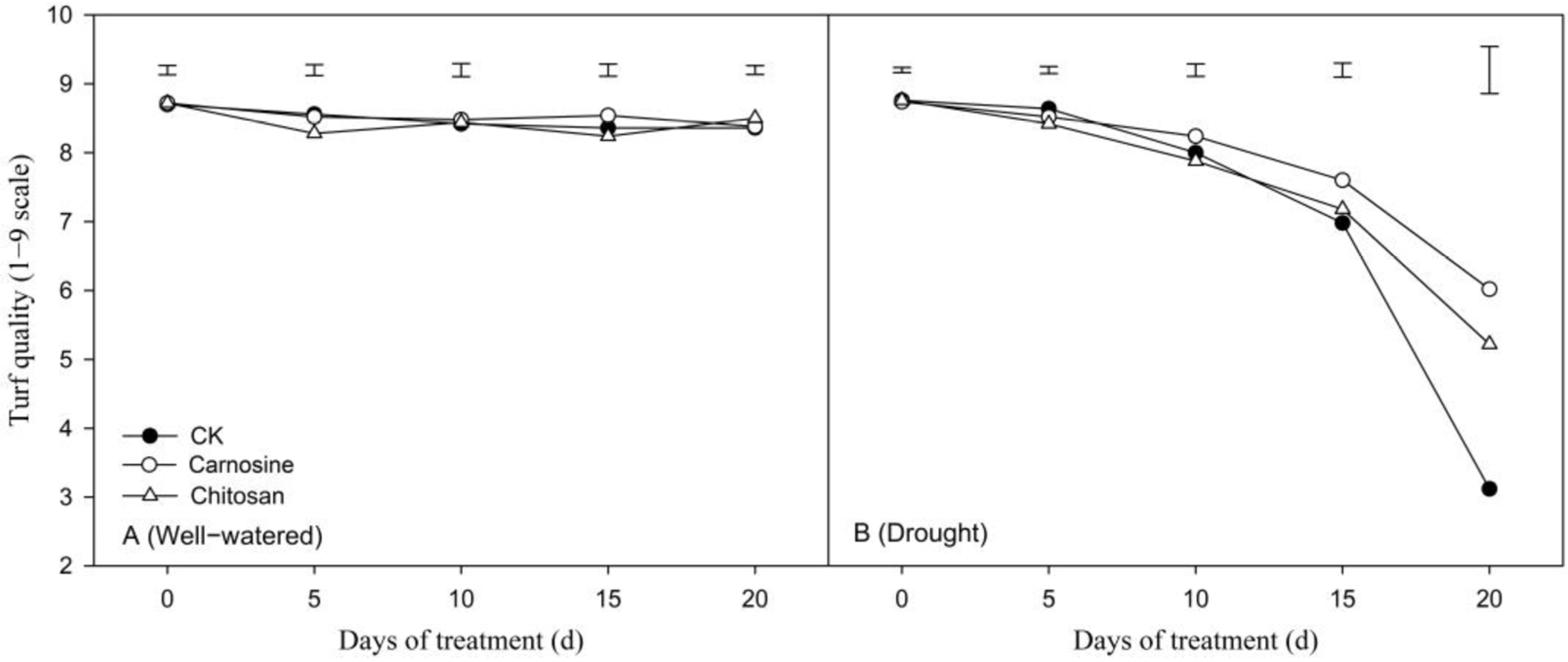
3.3. Effects of Carnosine and Chitosan on Turf Performance
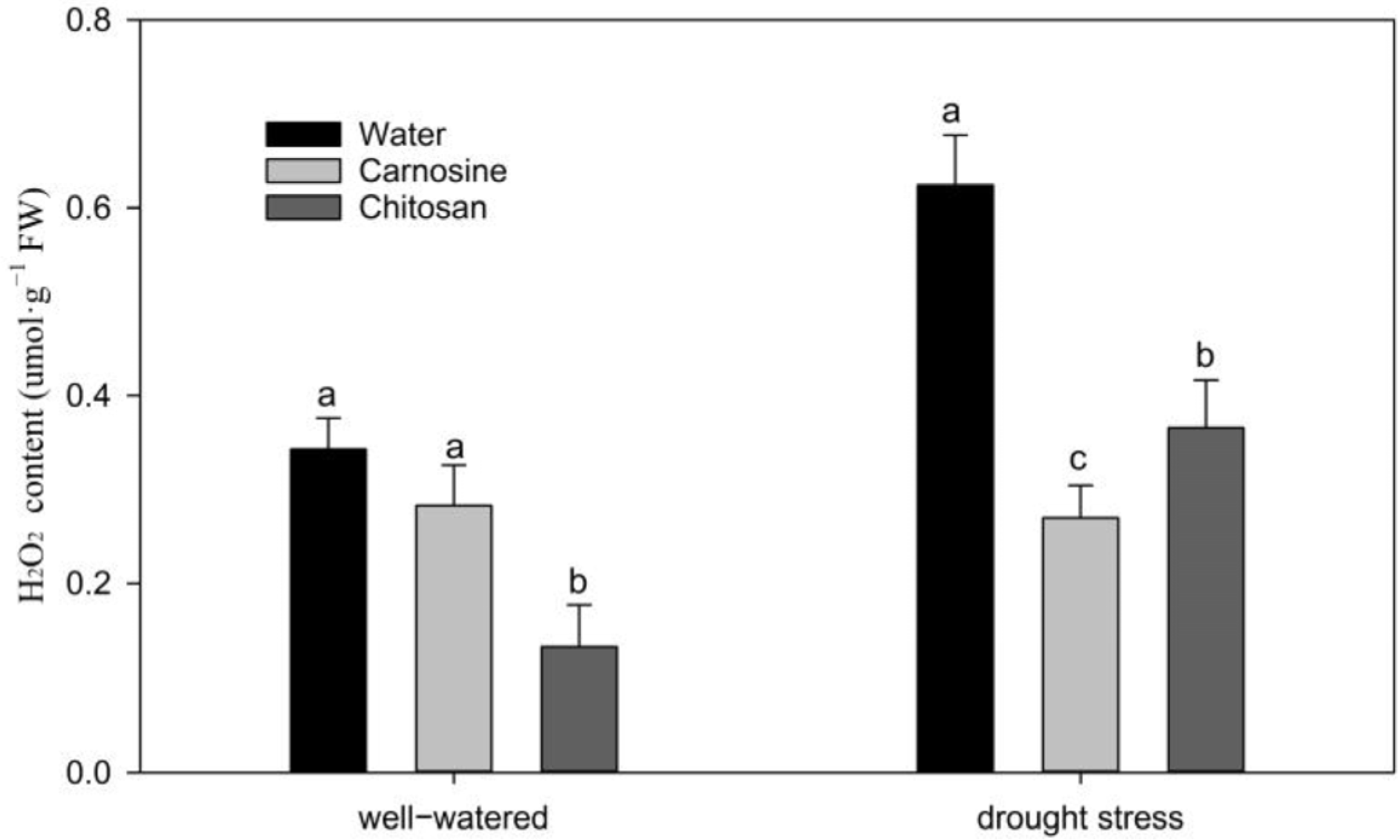
3.4. Effects of Carnosine and Chitosan on Cellular Membrane Stability
3.5. Effects of Carnosine and Chitosan on Enzyme Activities of Antioxidants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological response of Lallemantia iberica to brassinolide treatment under different watering conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Hamayun, M.; Khan, S.A.; Shinwari, Z.K.; Khan, A.L.; Ahmad, N.; Lee, I.J. Effect of polyethylene glycol induced drought stress on physio-hormonal attributes of soybean. Pak. J. Bot. 2010, 42, 977–986. [Google Scholar]
- Goswami, A.; Banerjee, R.; Raha, S. Drought resistance in rice seedlings conferred by seed priming. Protoplasma 2013, 250, 1115–1129. [Google Scholar] [CrossRef]
- Tallapragada, P.; Dikshit, R.; Seshagiri, S. Influence of rhizophagus spp. and burkholderia seminalis on the growth of tomato (Lycopersicon esculatum) and bell pepper (Capsicum annuum) under drought stress. Commun. Soil Sci. Plan. 2016, 47, 1975–1984. [Google Scholar] [CrossRef]
- Graça, J.P.D.; Rodrigues, F.A.; Farias, J.R.B.; Oliveira, M.C.N.D.; Hoffmann-Campo, C.B.; Zingaretti, S.M. Physiological parameters in sugarcane cultivars submitted to water deficit. Braz. J. Plant Physiol. 2010, 22, 189–197. [Google Scholar] [CrossRef]
- Akram, S.; Siddiqui, M.N.; Hussain, B.M.N.; Al Bari, M.A.; Mostofa, M.G.; Hossain, M.A.; Tran, L.P. Exogenous glutathione modulates salinity tolerance of soybean [Glycine max (L.) Merrill] at reproductive stage. J. Plant Growth Regul. 2017, 36, 877–888. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Belal Chowdhury, M.; Afrin, S.; Burritt, D.J.; Murata, Y.; Hossain, M.A.; Afzal Hossain, M. Exogenous glutathione-mediated drought stress tolerance in rice (Oryza sativa L.) is associated with lower oxidative damage and favorable ionic homeostasis. Iran. J. Sci. Technol. Trans. Sci. 2020, 44, 955–971. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Hu, L.; Bi, A.; Hu, Z.; Amombo, E.; Li, H.; Fu, J. Antioxidant metabolism, photosystem II, and fatty acid composition of two tall fescue genotypes with different heat tolerance under high temperature stress. Front. Plant Sci. 2018, 9, 1242. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, T.; Amombo, E.; Wang, G.; Xie, Y.; Fu, J. The alleviation of heat damage to photosystem II and enzymatic antioxidants by exogenous spermidine in tall fescue. Front. Plant Sci. 2017, 8, 1747. [Google Scholar] [CrossRef]
- Nair, A.S.; Abraham, T.K.; Jaya, D.S. Studies on the changes in lipid peroxidation and antioxidants in drought stress induced cowpea (Vigna unguiculata L.) varieties. J. Environ. Biol. 2008, 29, 689–691. [Google Scholar] [PubMed]
- Xiong, B.; Wang, Y.; Zhang, Y.; Ma, M.; Gao, Y.; Zhou, Z.; Wang, B.; Wang, T.; Lv, X.; Wang, X.; et al. Alleviation of drought stress and the physiological mechanisms in Citrus cultivar (Huangguogan) treated with methyl jasmonate. Biosci. Biotechnol. Biochem. 2020, 84, 1958–1965. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.; Zhang, T.; Qiao, Y.; Wen, P.; Zhai, G.; Liu, E.; Al-Bakre, D.A.; Al-Harbi, M.S.; Gao, X.; Yang, X. Glycinebetaine mitigates drought stress-induced oxidative damage in pears. PLoS ONE 2021, 16, e251389. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Physiol. Plant. 2021, 172, 1212–1226. [Google Scholar] [CrossRef]
- Manavalan, L.P.U.O.; Guttikonda, S.K.; Tran, L.S.P.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Geerts, S.; Raes, D. Deficit irrigation as an on-farm strategy to maximize crop water productivity in dry areas. Agr. Water Manag. 2009, 96, 1275–1284. [Google Scholar] [CrossRef]
- Lou, R.; Li, D.; Li, Y.; Bian, Z.; Zhu, Y. Effect of pre-anthesis drought hardening on post-anthesis physiological characteristics, yield and WUE in winter wheat. Phyton 2021, 90, 245–257. [Google Scholar] [CrossRef]
- Naz, R.; Batool, S.; Shahid, M.; Keyani, R.; Yasmin, H.; Nosheen, A.; Hassan, M.N.; Mumtaz, S.; Siddiqui, M.H. Exogenous silicon and hydrogen sulfide alleviates the simultaneously occurring drought stress and leaf rust infection in wheat. Plant Physiol. Biochem. 2021, 166, 558–571. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Ahmad, A.; Alyemeni, M.N.; Ahmad, P. Impact of exogenously applied trehalose on leaf biochemistry, achene yield and oil composition of sunflower under drought stress. Physiol. Plant 2021, 172, 317–333. [Google Scholar] [CrossRef]
- Demirkol, G. PopW enhances drought stress tolerance of alfalfa via activating antioxidative enzymes, endogenous hormones, drought related genes and inhibiting senescence genes. Plant Physiol. Bioch. 2021, 166, 540–548. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Li, Y.; Zhang, X.; Ma, X.; Huang, L.; Yan, Y.; Peng, Y. Chitosan and spermine enhance drought resistance in white clover, associated with changes in endogenous phytohormones and polyamines, and antioxidant metabolism. Funct. Plant Biol. 2018, 45, 1205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, X.; Dai, M. Improving crop drought resistance with plant growth regulators and rhizobacteria: Mechanisms, applications, and perspectives. Plant Commun. 2022, 3, 100228. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Wang, J.; Hu, M.; Ma, Y.; Wu, S.; Qi, G.; Qiu, K.; Zhang, H. Influences of beta-alanine and l-histidine supplementation on growth performance, meat quality, carnosine content, and mRNA expression of carnosine-related enzymes in broilers. Animals 2021, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Chanadda, S.; Panpradub, S.; Kasarat, P.; Satoshi, K.; Cindy, R.; Wittawat, M.; Jirawat, Y.; Kanjana, T.; Amonrat, M. Effects of β-alanine and L-histidine supplementation on carnosine contents in and quality and secondary structure of proteins in slow-growing Korat chicken meat. Poult. Sci. 2022, 101, 101776. [Google Scholar]
- Ozel, T.U.; Bilgihan, A.; Biberoglu, G.; Mertoglu, C.O. Carnosine supplementation protects rat brain tissue against ethanol-induced oxidative stress. Mol. Cell Biochem. 2010, 339, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, L.; Cheng, X.; Yang, Y.; Luan, B.; Jia, L.; Xu, F.; Zhang, Z. Carnosine pretreatment protects against hypoxia–ischemia brain damage in the neonatal rat model. Eur. J. Pharmacol. 2011, 667, 202–207. [Google Scholar] [CrossRef]
- Kohen, R.; Yamamoto, Y.; Cundy, K.C.; Ames, B.N. Antioxidant activity of carnosine, homocarnosine, and anserine present in muscle and brain. Proc. Natl. Acad. Sci. USA 1988, 85, 3175–3179. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, E.J.; Kim, Y.N.; Choi, C.; Lee, B.H. Effects of alpha-lipoic acid and L-carnosine supplementation on antioxidant activities and lipid profiles in rats. Nutr. Res. Pract. 2011, 5, 421–428. [Google Scholar] [CrossRef]
- Tokatlı, K.; Demirdöven, A. Effects of chitosan edible film coatings on the physicochemical and microbiological qualities of sweet cherry (Prunus avium L.). Sci. Hortic. 2020, 259, 108656. [Google Scholar] [CrossRef]
- Dang, Q.F.; Yan, J.Q.; Li, Y.; Cheng, X.; Liu, C.S.; Chen, X.G. Chitosan acetate as an active coating material and its effects on the storing of Prunus avium L. J. Food Sci. 2010, 2, S125–S131. [Google Scholar] [CrossRef] [PubMed]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 29. [Google Scholar] [CrossRef] [Green Version]
- Chien, P.; Sheu, F.; Yang, F. Effects of edible chitosan coating on quality and shelf life of sliced mango fruit. J. Food Eng. 2007, 78, 225–229. [Google Scholar] [CrossRef]
- Kumar, P.; Sethi, S.; Sharma, R.R.; Srivastav, M.; Varghese, E. Effect of chitosan coating on postharvest life and quality of plum during storage at low temperature. Sci. Hortic. 2017, 226, 104–109. [Google Scholar] [CrossRef]
- Boonlertnirun, S.; Sarobol, E.; Sooksathan, I. Effects of molecular weight of chitosan on yield potential of rice cultivar suphan buri 1. Kasetsart J. 2006, 40, 854–861. [Google Scholar]
- Uthairatanakij, A.; Silva, J.A.T.D.; Obsuwan, K. Chitosan for improving orchid production and quality. Orchid Sci. Biotechnol. 2007, 1, 1–5. [Google Scholar]
- Tourian, N.; Sinaki, J.M.; Hasani, N.; Madani, H. Change in photosynthetic pigment concentration of wheat grass (Agropyron repens) cultivars response to drought stress and foliar application with Chitosan. Intl. J. Agron. Plant Prod. 2013, 4, 1084–1091. [Google Scholar]
- Emami, B.Z.; Siadat, S.A.; Bakhshandeh, A.; Ghasemi, P.A.; Hashemi, M. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis Celak. Crop. J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Huang, S.; Jiang, S.; Liang, J.; Chen, M.; Shi, Y. Current knowledge of bermudagrass responses to abiotic stresses. Breed. Sci. 2019, 69, 215–226. [Google Scholar] [CrossRef]
- Hu, L.; Wang, Z.; Du, H.; Huang, B. Differential accumulation of dehydrins in response to water stress for hybrid and common bermudagrass genotypes differing in drought tolerance. J. Plant Physiol. 2010, 167, 103–109. [Google Scholar] [CrossRef]
- Turgeon, R. Phloem loading and plasmodesmata. Trends Plant Sci. 1996, 1, 418–423. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity techniques for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop. Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar]
- Xu, L.; Han, L.; Huang, B. Antioxidant enzyme activities and gene expression patterns in leaves of kentucky bluegrass in response to drought and post-drought recovery. J. Am. Soc. Hortic. Sci. 2011, 136, 247–255. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Fidilio, A.; Donnell, F.O.; Musso, N.; Lazzarino, G.; Grasso, M.; Amorini, A.M.; Tascedda, F.; Bucolo, C.; et al. Carnosine decreases PMA-induced oxidative stress and inflammation in murine macrophages. Antioxidants 2019, 8, 281. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agr. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef]
- Huang, C.; Tian, Y.; Zhang, B.; Hassan, M.J.; Li, Z.; Zhu, Y. Chitosan (CTS) alleviates heat-induced leaf senescence in creeping bentgrass by regulating chlorophyll metabolism, antioxidant defense, and the heat shock pathway. Molecules 2021, 26, 5337. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. Balancing the generation and elimination of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2005, 102, 3175–3176. [Google Scholar] [CrossRef] [PubMed]
- Vanisri, S.; Sreedhar, M.; Jeevan, L.; Pavani, A.; Chaturvedi, A.; Aparna, M.; Pavan Kumar, D.; Sunitha, T.; Aruna, K.; Tripura Venkata, V.G.N.; et al. Evaluation of rice genotypes for chlorophyll content and scavenging enzyme activity under the influence of mannitol stress towards drought tolerance. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2907–2917. [Google Scholar] [CrossRef]
- Yuan, G.; Jia, C.; Li, Z.; Sun, B.; Zhang, L.; Liu, N.; Wang, Q. Effect of brassinosteroids on drought resistance and abscisic acid concentration in tomato under water stress. Sci. Hortic. 2010, 126, 103–108. [Google Scholar] [CrossRef]
- Mishra, A.; Choudhuri, M.A. Effects of salicylic acid on heavy metal-induced. Biol. Plant. 1999, 42, 409–415. [Google Scholar] [CrossRef]
- Yu, Y.F.; Yang, Y.H.; Pan, S.J.; Xu, Z.M.; Cen, Y.; Wang, Y. Effects of L-carnosine on seed germination and physiological characteristics of rice under high temperature stress. Mol. Plant Breed. 2022, 11, 1587. [Google Scholar]
- Hong, K.; Xie, J.; Zhang, L.; Sun, D.; Gong, D. Effects of chitosan coating on postharvest life and quality of guava (Psidium guajava L.) fruit during cold storage. Sci. Hortic. 2012, 144, 172–178. [Google Scholar] [CrossRef]
- Xu, Q.J.; Nian, Y.G.; Jin, X.C.; Yan, C.Z.; Liu, J.; Jiang, G.M. Effects of chitosan on growth of an aquatic plant (Hydrilla verticillata) in polluted waters with different chemical oxygen demands. J. Environ. Sci. 2007, 19, 217–221. [Google Scholar] [CrossRef]
- Yang, F.; Hu, J.; Li, J.; Wu, X.; Qian, Y. Chitosan enhances leaf membrane stability and antioxidant enzyme activities in apple seedlings under drought stress. Plant Growth Regul. 2009, 58, 131–136. [Google Scholar] [CrossRef]
- Gerami, M.; Majidian, P.; Ghorbanpour, A.; Alipour, Z. Stevia rebaudiana Bertoni responses to salt stress and chitosan elicitor. Physiol. Mol. Biol. Plants 2020, 26, 965–974. [Google Scholar] [CrossRef]
- Xie, W.; Xu, P.; Liu, Q. Antioxidant activity of water-soluble chitosan derivatives. Bioorg. Med. Chem. Lett. 2001, 11, 1699–1701. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, T.; Yang, Z.; Liang, J.; Yu, J.; Liu, J. Foliar Application of Carnosine and Chitosan Improving Drought Tolerance in Bermudagrass. Agronomy 2023, 13, 442. https://doi.org/10.3390/agronomy13020442
Hao T, Yang Z, Liang J, Yu J, Liu J. Foliar Application of Carnosine and Chitosan Improving Drought Tolerance in Bermudagrass. Agronomy. 2023; 13(2):442. https://doi.org/10.3390/agronomy13020442
Chicago/Turabian StyleHao, Tian, Zhimin Yang, Jianfeng Liang, Jingjin Yu, and Jun Liu. 2023. "Foliar Application of Carnosine and Chitosan Improving Drought Tolerance in Bermudagrass" Agronomy 13, no. 2: 442. https://doi.org/10.3390/agronomy13020442