
Study on Root Hydraulic Lift of Drought-Tolerant and Drought-Sensitive Potato Cultivars (Solanum tuberosum L.)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design
2.3. Drought Treatments
2.4. Sampling and Measuring
2.5. Data Analysis
3. Results
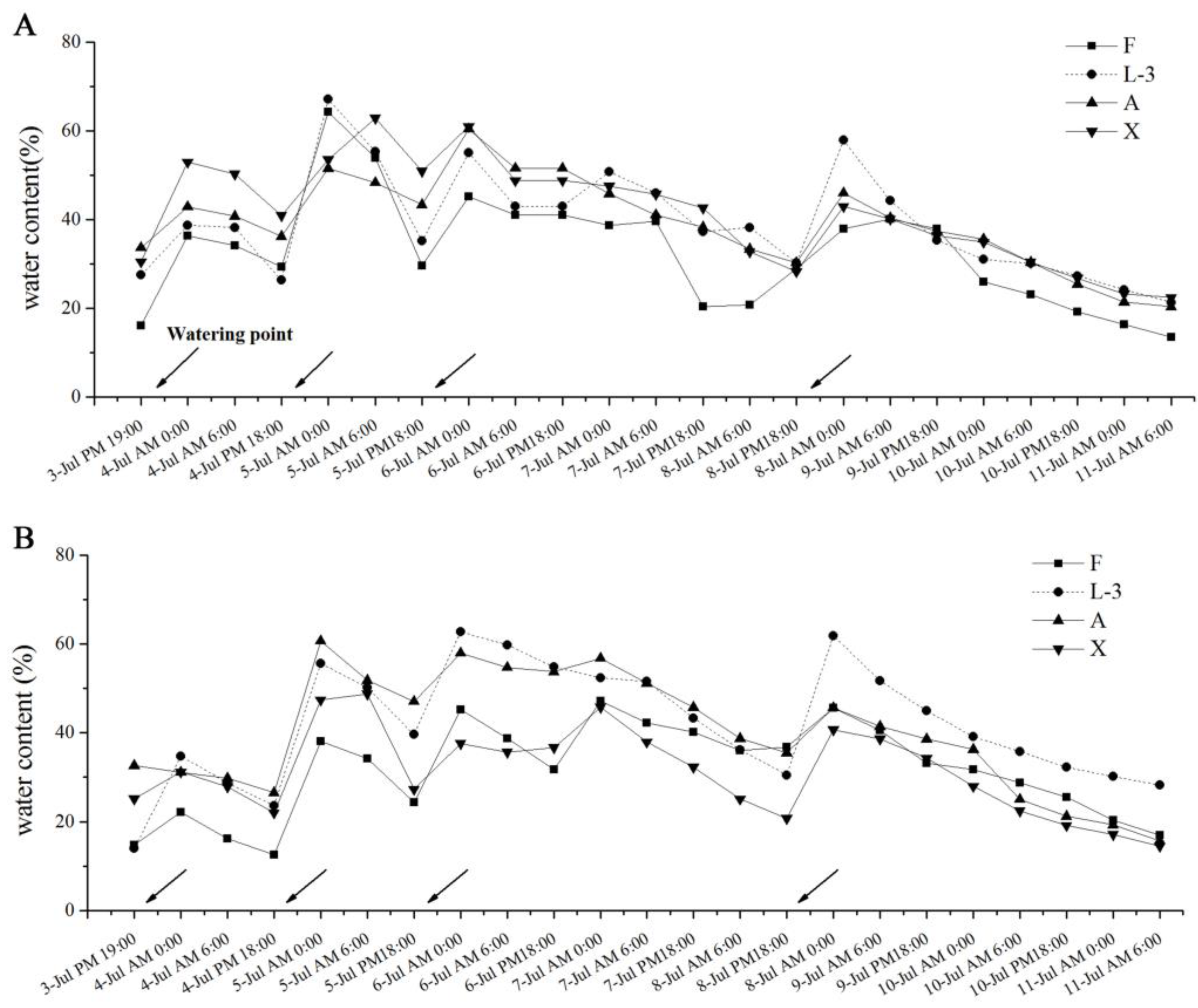
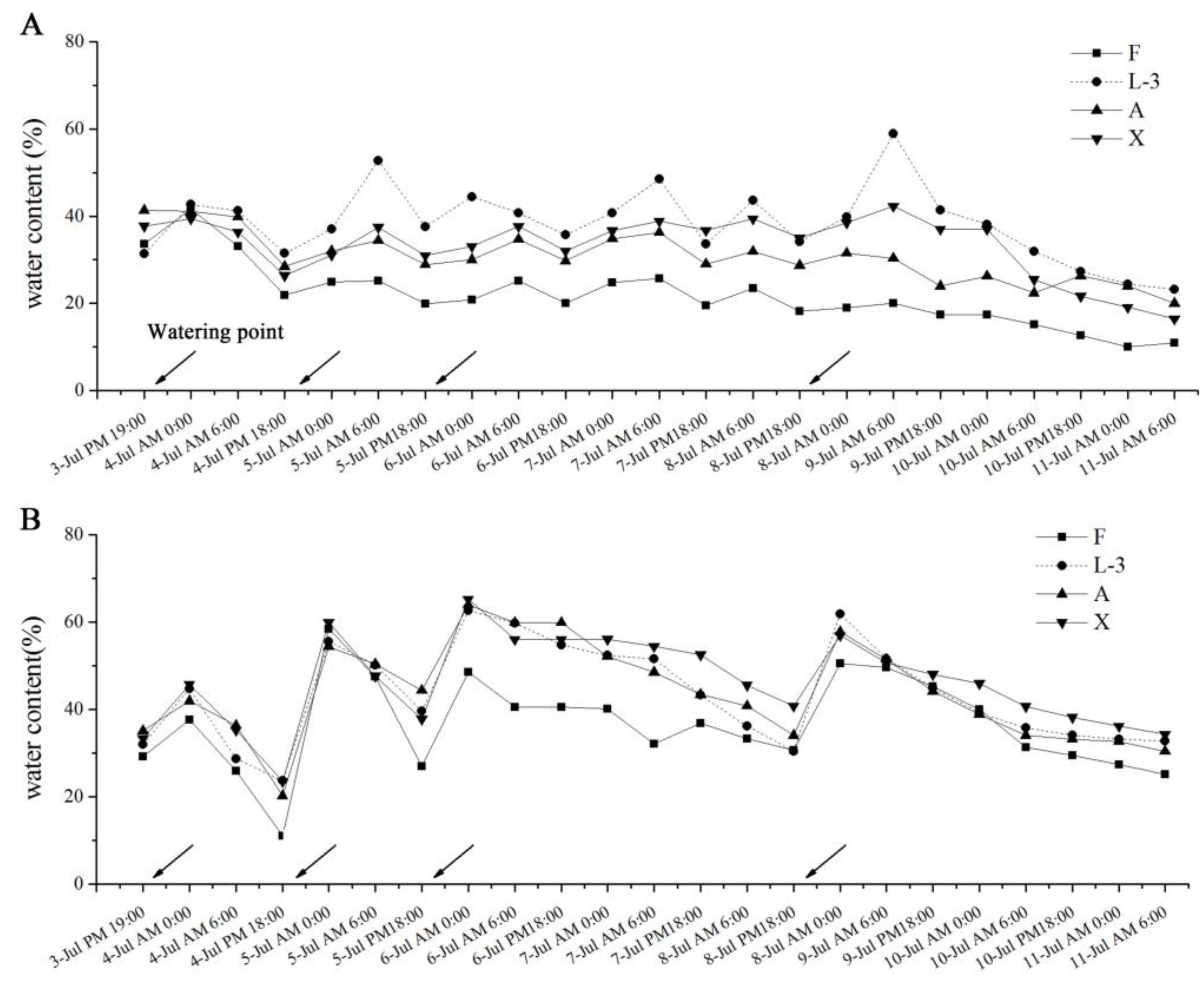
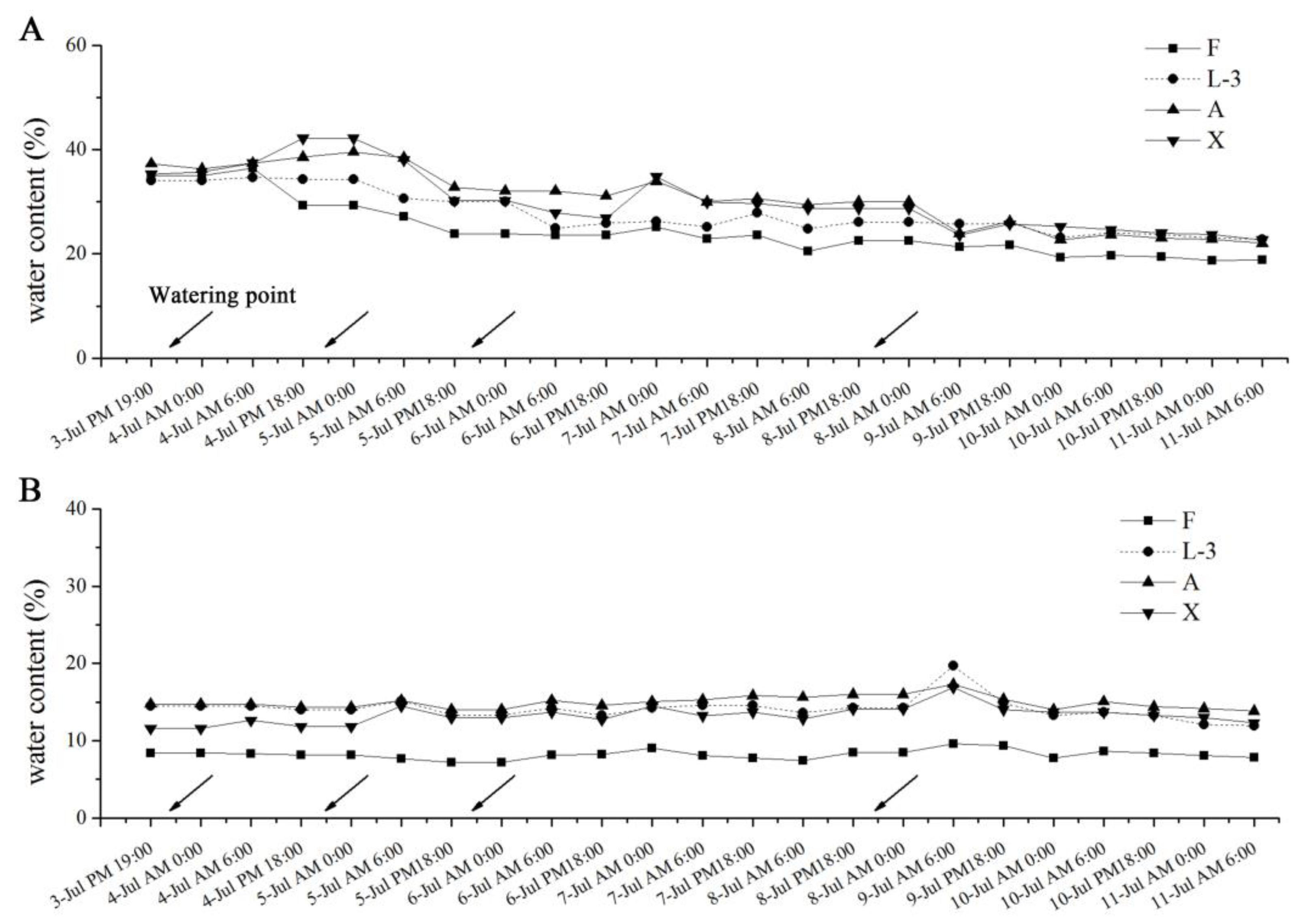
3.1. Changes in Soil Moisture under Different Treatments
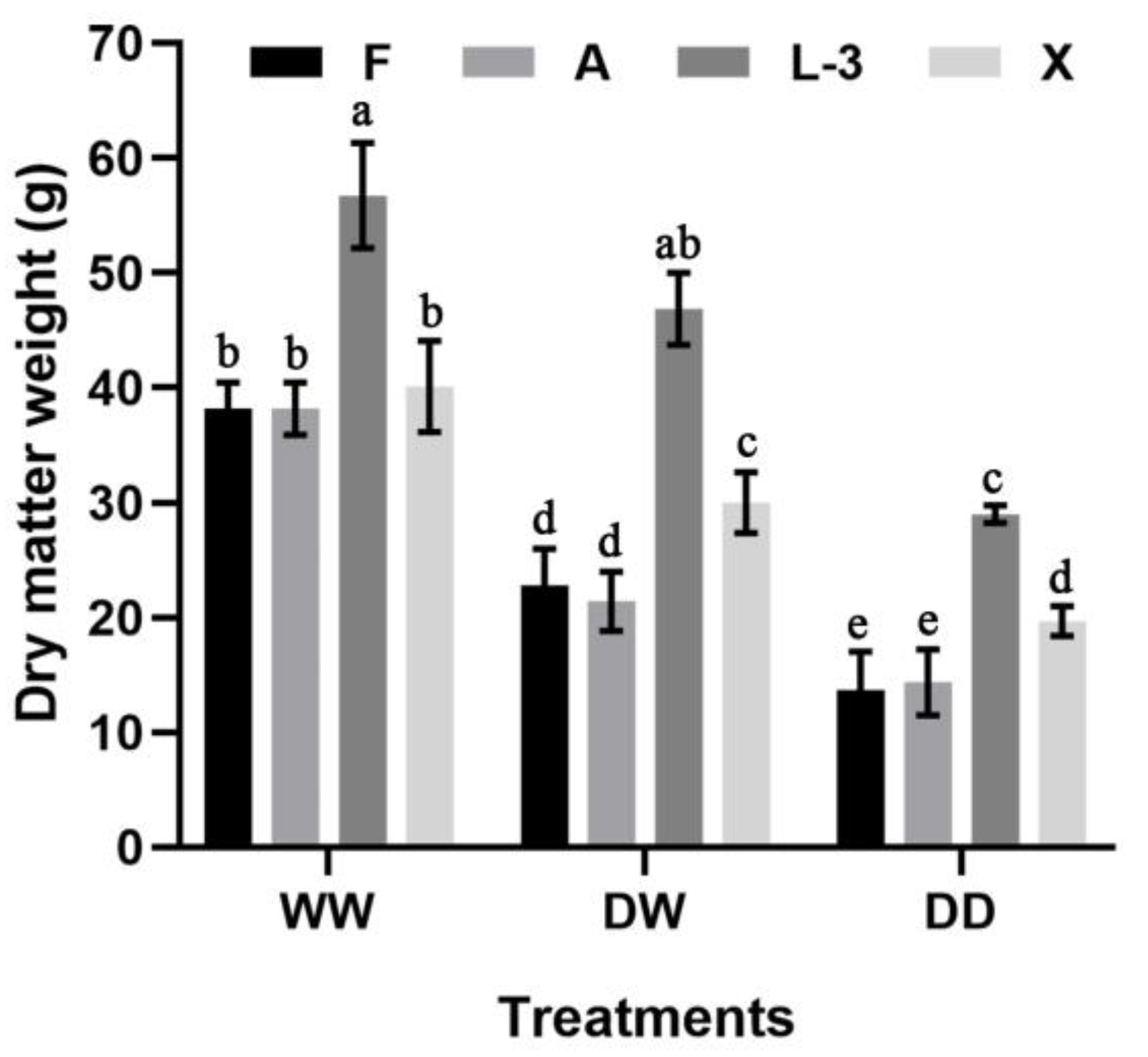
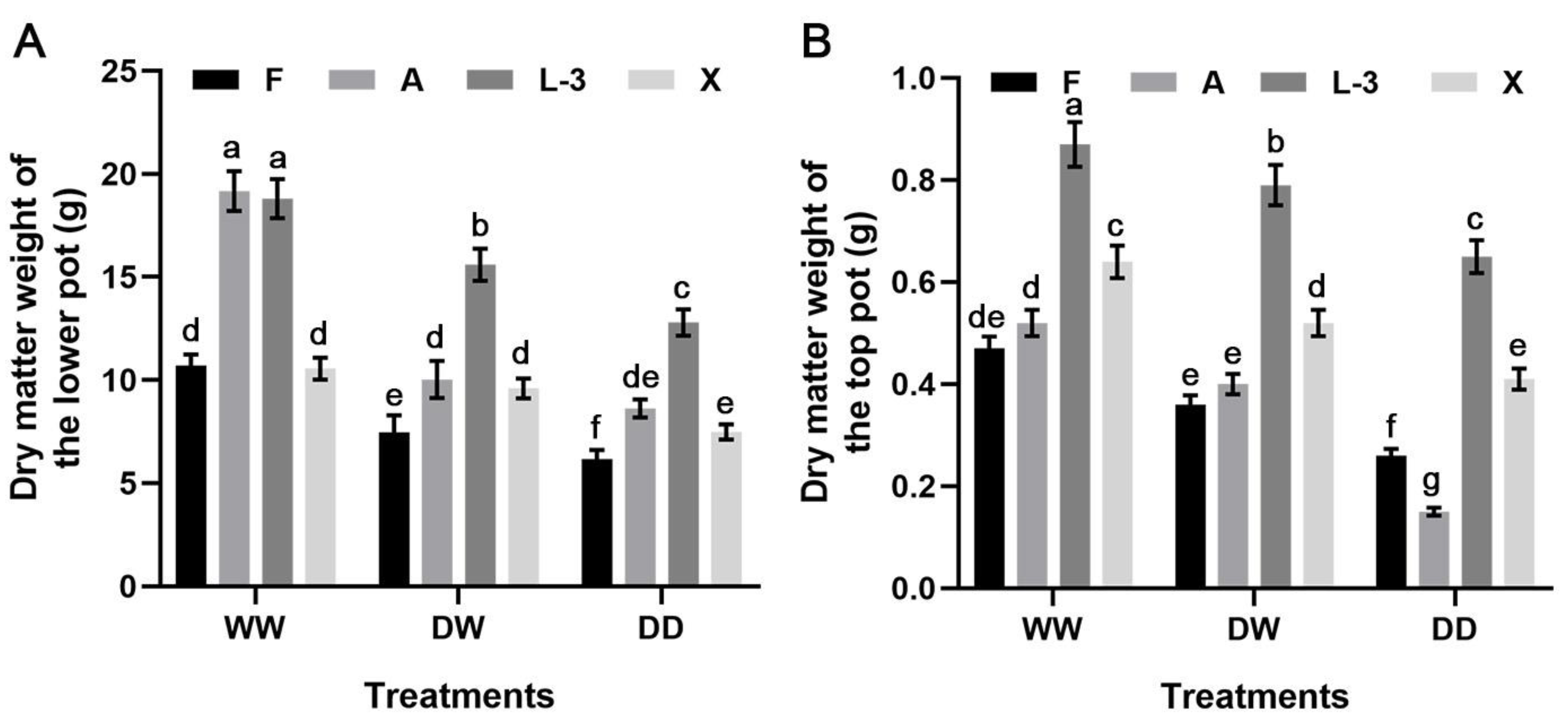
3.2. Changes in Biomass under Different Treatments
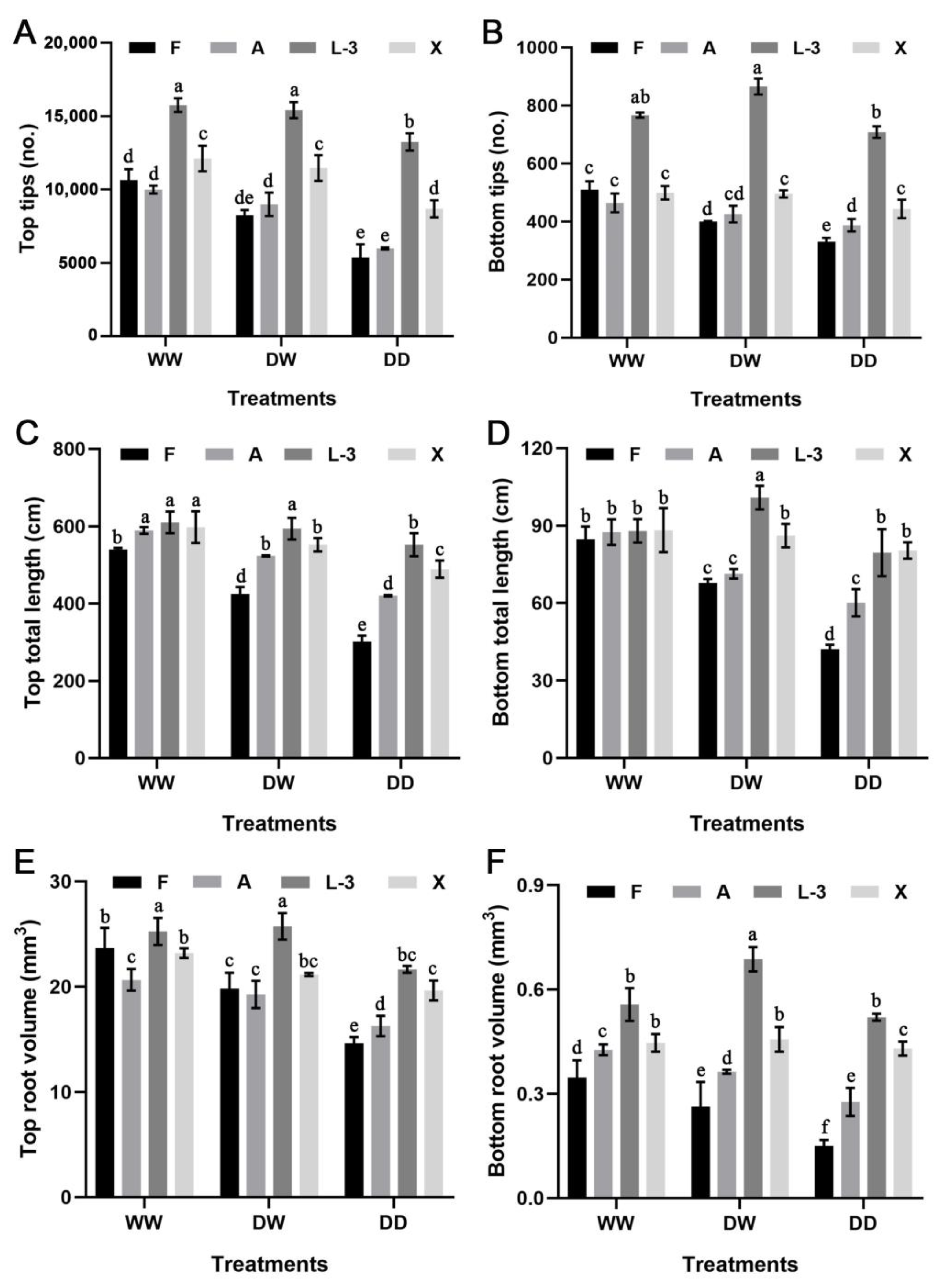
3.3. Changes in the Root Architecture under Different Treatments
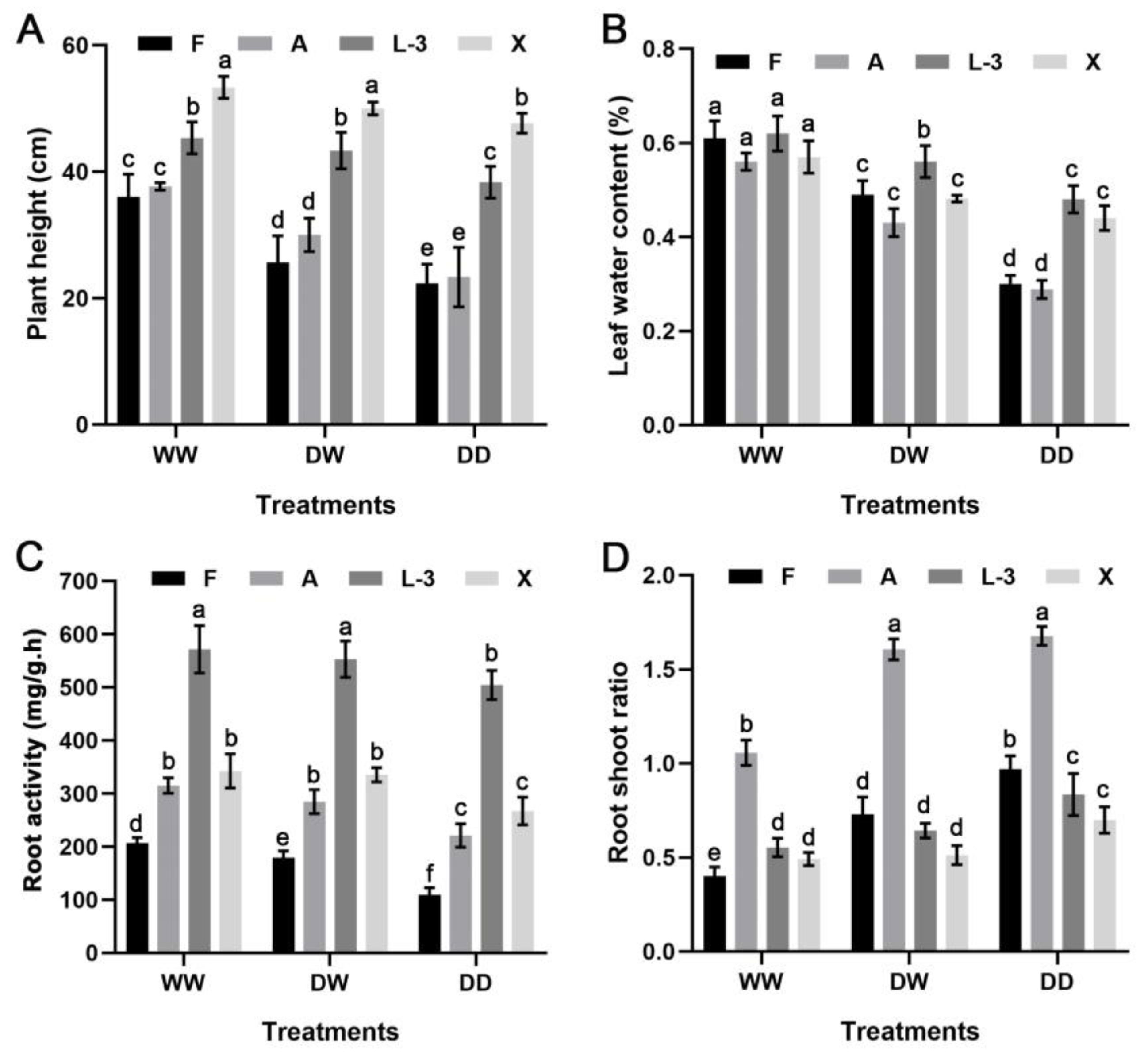
3.4. Changes in the Plant Height and LWC under Different Treatments
3.5. Changes in the Root Activity and Root–Shoot Ratio under Different Treatments
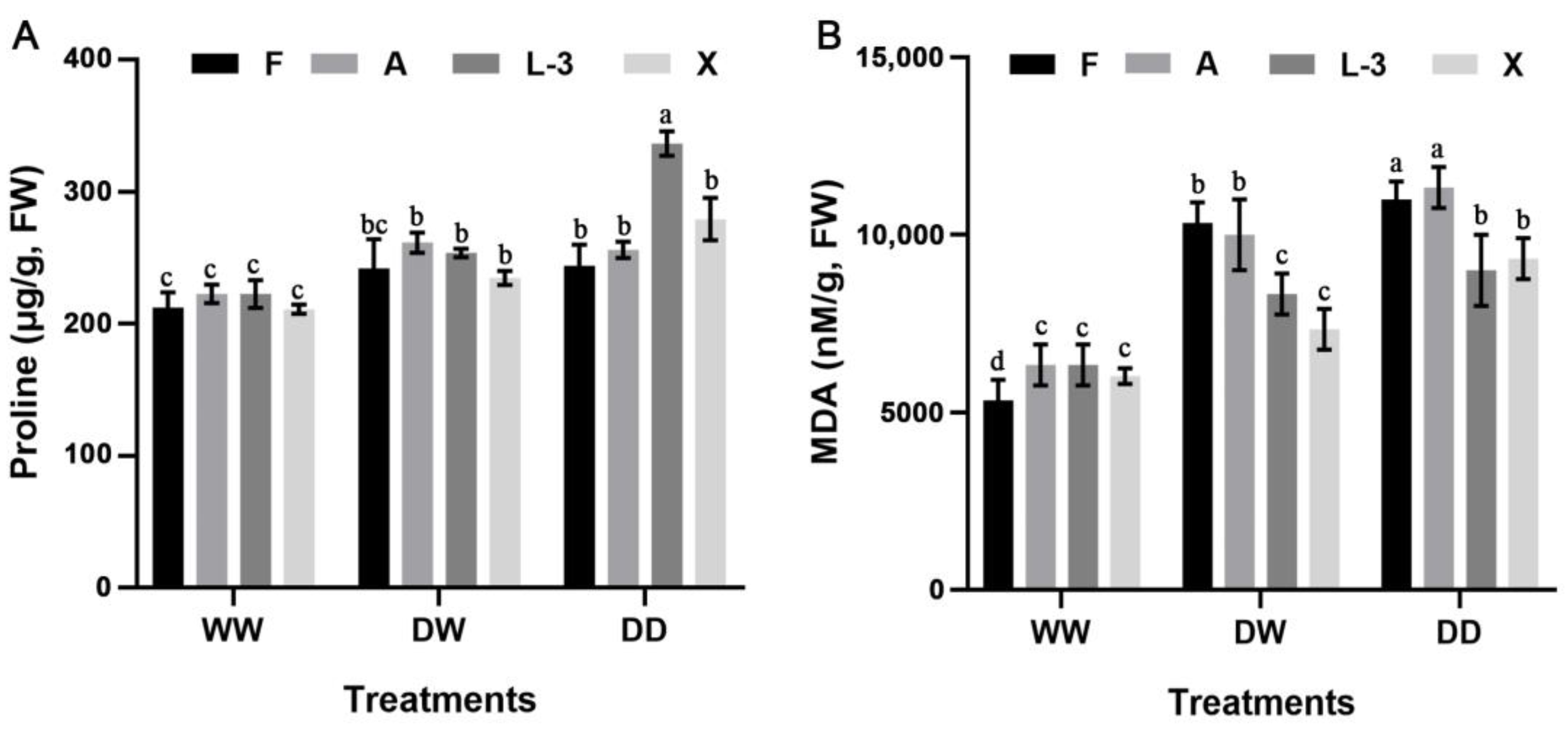
3.6. Changes in the Proline Content under Different Treatments
3.7. Changes in the MDA Content under Different Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alagele, S.M.; Jose, S.; Anderson, S.H.; Udawatta, R.P. Hydraulic lift: Processes, methods, and practical implications for society. Agrofor. Syst. 2021, 95, 641–657. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Zwiazek, J.J. Hydraulic redistribution in slender wheatgrass (Elymus trachycaulus Link Malte) and yellow sweet clover (Melilotus officinalis L.): Potential benefits for land reclamation. Agronomy 2018, 8, 308. [Google Scholar] [CrossRef]
- Caldwell, M.; Richards, J. Hydraulic lift: Water efflux from upper roots improves effectiveness of water uptake by deep roots. Oecologia 1989, 79, 1–5. [Google Scholar] [CrossRef]
- Caldwell, M.M.; Dawson, T.E.; Richards, J.H. Hydraulic lift: Consequences of water efflux from the roots of plants. Oecologia 1998, 113, 151–161. [Google Scholar] [CrossRef]
- Domec, J.; Warren, J.; Meinzer, F.; Brooks, J.; Coulombe, R. Native root xylem embolism and stomatal closure in stands of Douglas-fir and ponderosa pine: Mitigation by hydraulic redistribution. Oecologia 2004, 141, 7–16. [Google Scholar] [CrossRef]
- Xu, W.; Niu, F.; Kang, F.; Xu, B.; Deng, X.; Palta, J.A.; Chen, Y. Biomass allocation and competitive ability of a semiarid perennial grass and a legume in Mixtures under periodical soil water decreasing conditions. Agronomy 2022, 12, 2256. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Dawson, T.E.; Burgess, S.S.; Nepstad, D.C. Hydraulic redistribution in three Amazonian trees. Oecologia 2005, 145, 354–363. [Google Scholar] [CrossRef]
- Mohammadi, A.S.; Zahra, N.; Hajiaghaei, K.M.; Asgari, L.B.; Nobaharan, K.; Astatkie, T.; Siddique, K.H.; Farooq, M. Role of root hydraulics in plant drought tolerance. J. Plant Growth Regul. 2022, 17, 1–16. [Google Scholar] [CrossRef]
- Zapater, M.; Hossann, C.; Bréda, N.; Bréchet, C.; Bonal, D.; Granier, A. Evidence of hydraulic lift in a young beech and oak mixed forest using 18O soil water labelling. Trees 2011, 25, 885–894. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, B.; Xie, Z. Impacts of hydraulic redistribution on eco-hydrological cycles: A case study over the Amazon basin. Sci. China Earth Sci. 2018, 61, 1330–1340. [Google Scholar] [CrossRef]
- Neumann, R.B.; Cardon, Z.G. The magnitude of hydraulic redistribution by plant roots: A review and synthesis of empirical and modeling studies. New Phytol. 2012, 194, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Bauerle, T.; Richards, J.; Smart, D.; Eissenstat, D. Importance of internal hydraulic redistribution for prolonging the lifespan of roots in dry soil. Plant Cell Environ. 2008, 31, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Smart, D.; Carlisle, E.; Goebel, M.; Nuñez, B.A. Transverse hydraulic redistribution by a grapevine. Plant Cell Environ. 2005, 28, 157–166. [Google Scholar] [CrossRef]
- Ruan, S.; Wang, L.; Li, Y.; Li, P.; Ren, Y.; Gao, R.; Ma, H. Staple food and health: A comparative study of physiology and gut microbiota of mice fed with potato and traditional staple foods (corn, wheat and rice). Food Funct. 2021, 12, 1232–1240. [Google Scholar] [CrossRef]
- Dahal, K.; Li, X.-Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving potato stress tolerance and tuber yield under a climate change scenario—A current overview. Front. Plant Sci. 2019, 10, 563. [Google Scholar] [CrossRef]
- Ji, A.; It, B.; Id, A. Influence of water-based and contact heating preparation methods on potato mechanical properties, mastication, and sensory perception. Int. J. Gastron. Food Sci. 2021, 25, 100401. [Google Scholar]
- Nasir, M.W.; Toth, Z. Effect of drought stress on potato production: A review. Agronomy 2022, 12, 635. [Google Scholar] [CrossRef]
- Boguszewska-Mańkowska, D.; Zarzyńska, K.; Nosalewicz, A. Drought differentially affects root system size and architecture of potato cultivars with differing drought tolerance. Am. J. Potato Res. 2020, 97, 54–62. [Google Scholar] [CrossRef]
- Hill, D.; Nelson, D.; Hammond, J.; Bell, L. Morphophysiology of potato (Solanum tuberosum) in response to drought stress: Paving the way forward. Front. Plant Sci. 2021, 11, 597554. [Google Scholar] [CrossRef]
- Liu, F.; Shahnazari, A.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J. Exp. Bot. 2006, 57, 3727–3735. [Google Scholar] [CrossRef]
- Satchithanantham, S.; Krahn, V.; Sri Ranjan, R.; Sager, S. Shallow groundwater uptake and irrigation water redistribution within the potato root zone. Agric. Water Manag. 2014, 132, 101–110. [Google Scholar] [CrossRef]
- Aschmedes, G.; Hlmer. A new thiobarbituric acid (TBA) method for determining free malondialdehyde (MDA) and hydroperoxides selectively as a measure of lipid peroxidation. J. Am. Oil Chem. Soc. 1989, 66, 813–817. [Google Scholar] [CrossRef]
- Whalley, W.R.; Binley, A.; Watts, C.; Shanahan, P.; Dodd, I.C.; Ober, E.; Ashton, R.; Webster, C.; White, R.; Hawkesford, M.J. Methods to estimate changes in soil water for phenotyping root activity in the field. Plant Soil 2017, 415, 407–422. [Google Scholar] [CrossRef]
- Yang, Y.; Reddy, K.R.; Du, Y.; Fan, R. Short-term hydraulic conductivity and consolidation properties of soil-bentonite backfills exposed to CCR-impacted groundwater. J. Geotech. Geoenviron. 2018, 144, 04018025. [Google Scholar] [CrossRef]
- Müllers, Y.; Postma, J.A.; Poorter, H.; van Dusschoten, D. Stomatal conductance tracks soil-to-leaf hydraulic conductance in faba bean and maize during soil drying. Plant Physiol. 2022, 1, 2279–2294. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Sack, L.; Ort, D. The causes and consequences of leaf hydraulic decline with dehydration. J. Exp. Bot. 2017, 68, 4479–4496. [Google Scholar] [CrossRef]
- Ishikawa, C.M.; Bledsoe, C. Seasonal and diurnal patterns of soil water potential in the rhizosphere of blue oaks: Evidence for hydraulic lift. Oecologia 2000, 125, 459–465. [Google Scholar] [CrossRef]
- Colombi, T.; Torres, L.C.; Walter, A.; Keller, T. Feedbacks between soil penetration resistance, root architecture and water uptake limit water accessibility and crop growth–A vicious circle. Sci. Total Environ. 2018, 626, 1026–1035. [Google Scholar] [CrossRef]
- Xu, H.; He, H.; Yang, K.; Ren, H.; Zhu, T.; Ke, J.; You, C.; Guo, S.; Wu, L. Application of the nitrogen nutrition index to estimate the yield of indica hybrid rice grown from machine-transplanted bowl seedlings. Agronomy 2022, 12, 742. [Google Scholar] [CrossRef]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef]
- Chen, L.; Wang, C.; Yang, S.; Guan, X.; Zhang, Q.; Shi, M.; Yang, S.T.; Chen, C.; Chang, X.L. Chemical reduction of graphene enhances in vivo translocation and photosynthetic inhibition in pea plants. Environ. Sci. Nano 2019, 6, 1077–1088. [Google Scholar] [CrossRef]
- Tahir, M.H.N.; Imran, M.; Hussain, M.K. Evaluation of sunflower (Helianthus annuus L.) inbred lines for drought tolerance. Int. J. Agric. Biol. 2002, 3, 398–400. [Google Scholar]
- Seiler, J.; Cazell, B. Influence of water stress on the physiology and growth of red spruce seedlings. Tree Physiol. 1990, 6, 69–77. [Google Scholar] [CrossRef]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Proline accumulation is associated with inhibition of rice seedling root growth caused by NaCl. Plant Sci. 1996, 114, 121–128. [Google Scholar] [CrossRef]
- Esfandiari, E.; Shakiba, M.; Mahboob, S.; Alyari, H.; Shahabivand, S. The effect of water stress on the antioxidant content, protective enzyme activities, proline content and lipid peroxidation in wheat seedling. Pak. J. Biol. Sci. 2008, 11, 1916–1922. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Rems, L.; Viano, M.; Kasimova, M.A.; Miklavčič, D.; Tarek, M. The contribution of lipid peroxidation to membrane permeability in electropermeabilization: A molecular dynamics study. Bioelectrochemistry 2019, 125, 46–57. [Google Scholar] [CrossRef]
- Slama, I.; Tayachi, S.; Jdey, A.; Rouached, A.; Abdelly, C. Differential response to water deficit stress in alfalfa (Medicago sativa) cultivars: Growth, water relations, osmolyte accumulation and lipid peroxidation. Afr. J. Biotechnol. 2011, 10, 16250–16259. [Google Scholar]
- Wang, S.; Liang, D.; Li, C.; Hao, Y.; Ma, F.; Shu, H. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks. Plant Physiol. Biochem. 2012, 51, 81–89. [Google Scholar] [CrossRef]
- Jangid, K.K.; Dwivedi, P. Physiological and biochemical changes by nitric oxide and brassinosteroid in tomato (Lycopersicon esculentum Mill.) under drought stress. Acta Physiol. Plant. 2017, 39, 73. [Google Scholar] [CrossRef]
- Sairam, R.; Srivastava, G.; Agarwal, S.; Meena, R. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant. 2005, 49, 85–91. [Google Scholar] [CrossRef]
- Fazeli, F.; Ghorbanli, M.; Niknam, V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars. Biol. Plant. 2007, 51, 98–103. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RL | RT | RV | DM | Pro | MDA | HL | |
|---|---|---|---|---|---|---|---|
| RL | 1.000 | 0.787 ** | 0.908 ** | 0.826 ** | −0.181 | −0.862 ** | 0.597 * |
| RT | 0.787 ** | 1.000 | 0.948 ** | 0.823 ** | 0.087 | −0.541 | 0.355 * |
| RV | 0.908 ** | 0.948 ** | 1.000 | 0.829 ** | 0.052 | −0.657 * | 0.501 ** |
| DM | 0.826 ** | 0.823 ** | 0.829 ** | 1.000 | −0.395 | −0.817 ** | 0.600 * |
| Pro | −0.181 | 0.087 | 0.052 | −0.395 | 1.000 | 0.540 | −0.441 |
| MDA | −0.862 ** | −0.541 | −0.657 * | −0.817 ** | 0.540 | 1.000 | −0.683 ** |
| HL | 0.597 * | 0.355 * | 0.501 ** | 0.600 * | −0.441 | −0.683** | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, P.; Li, Y.; Ali, K.; Zhang, C.; Qin, T.; Bi, Z.; Liu, Y.; Liu, Z.; Kear, P.; Sun, C.; et al. Study on Root Hydraulic Lift of Drought-Tolerant and Drought-Sensitive Potato Cultivars (Solanum tuberosum L.). Agronomy 2023, 13, 443. https://doi.org/10.3390/agronomy13020443
Yao P, Li Y, Ali K, Zhang C, Qin T, Bi Z, Liu Y, Liu Z, Kear P, Sun C, et al. Study on Root Hydraulic Lift of Drought-Tolerant and Drought-Sensitive Potato Cultivars (Solanum tuberosum L.). Agronomy. 2023; 13(2):443. https://doi.org/10.3390/agronomy13020443
Chicago/Turabian StyleYao, Panfeng, Yajie Li, Kazim Ali, Chunli Zhang, Tianyuan Qin, Zhenzhen Bi, Yuhui Liu, Zhen Liu, Philip Kear, Chao Sun, and et al. 2023. "Study on Root Hydraulic Lift of Drought-Tolerant and Drought-Sensitive Potato Cultivars (Solanum tuberosum L.)" Agronomy 13, no. 2: 443. https://doi.org/10.3390/agronomy13020443