
Microwaves Induce Histological Alteration of Ovaries and Testis in Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae)
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing of the Insect
2.2. MW Irradiation
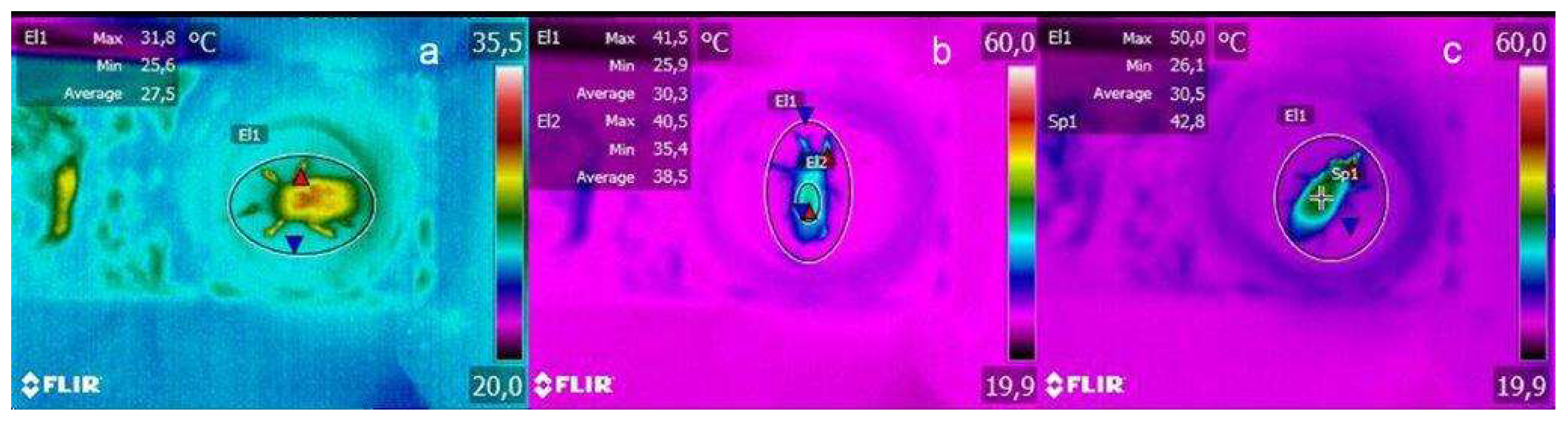
2.3. Thermogram Analysis
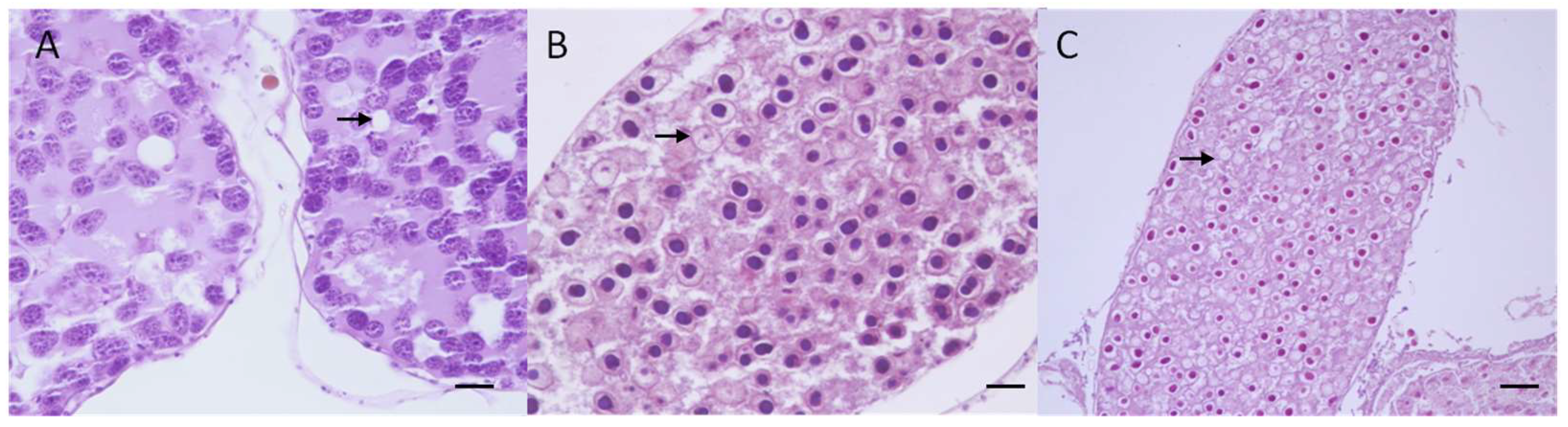
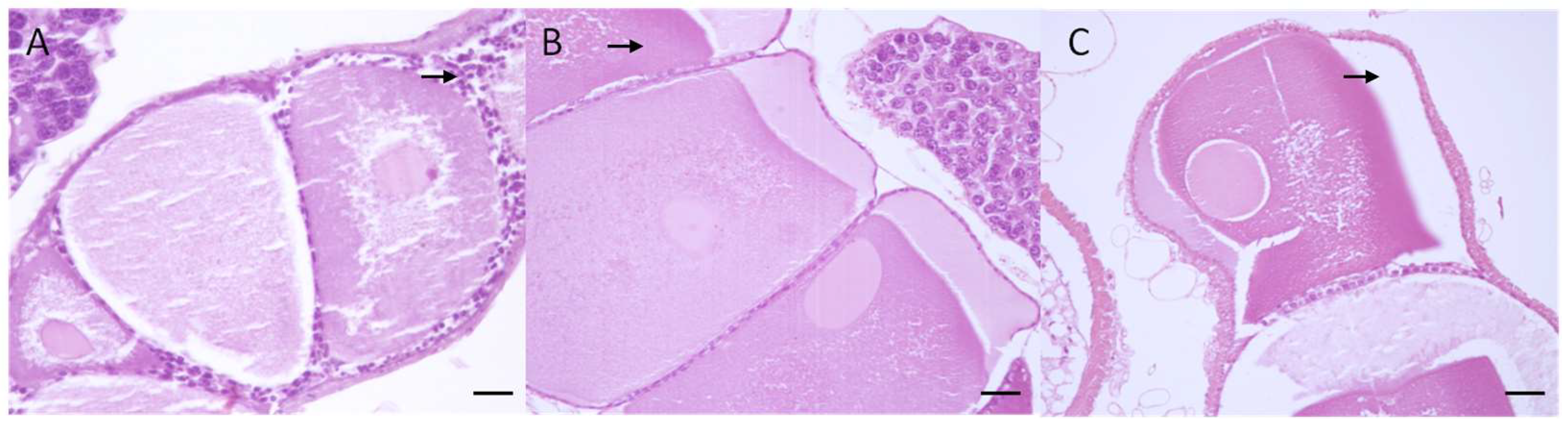
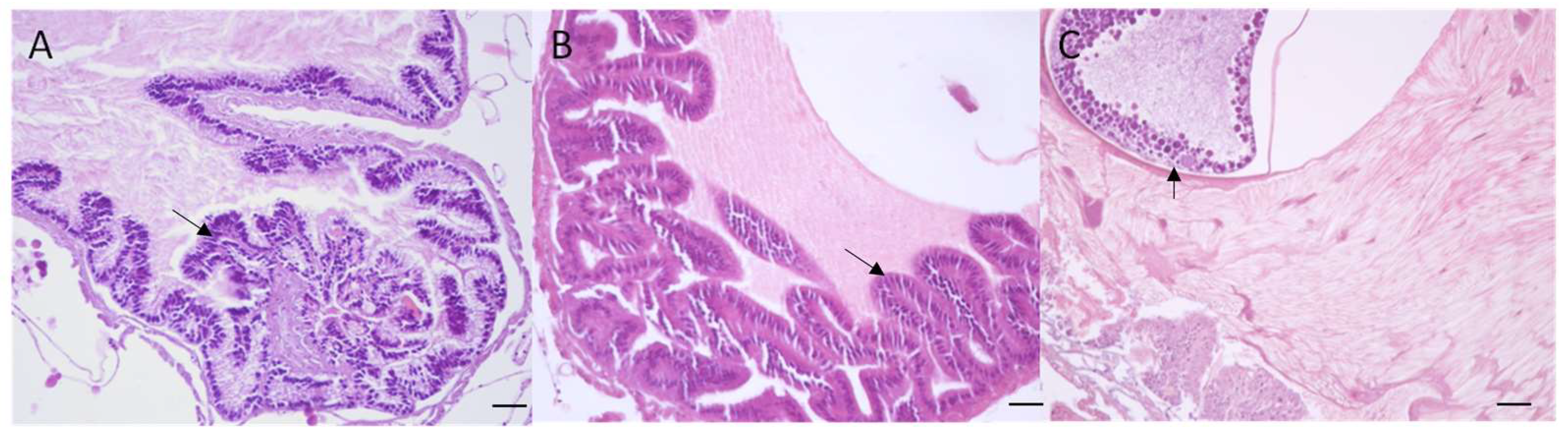
2.4. Macroscopical and Microscopical Analysis
2.5. Statistical Analysis
3. Results
3.1. Thermogram Results
3.2. Survival and Fecundity Data
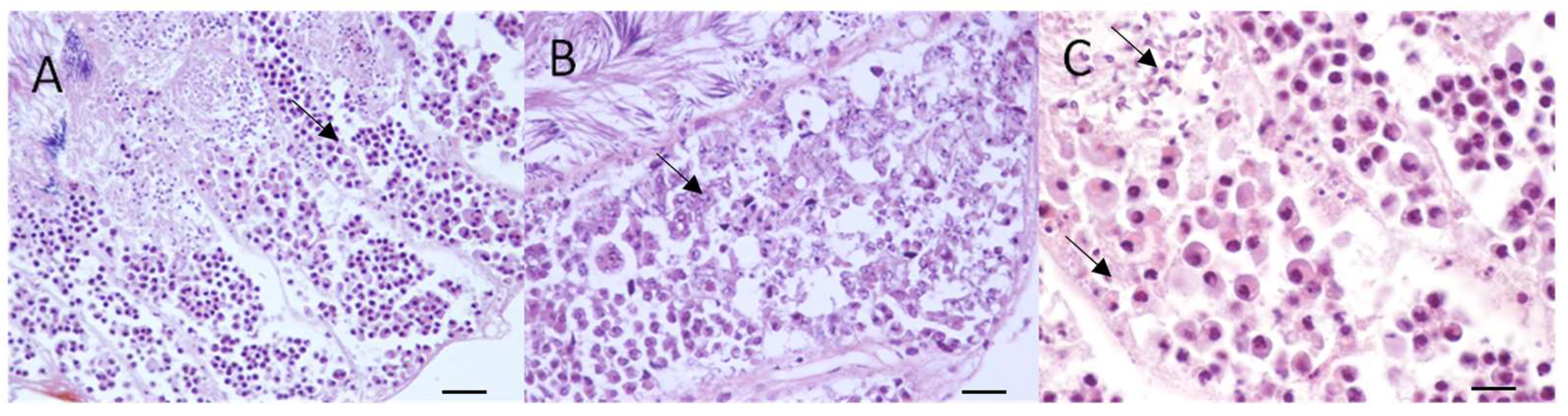
3.3. Macroscopical and Microscopical Results
3.4. Statistical Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffo, R.; Martino, V.; Pesapane, G. Rhynchophorus ferrugineus detected on other species of palms. Il Floric. 2008, 1–2, 37–39. Available online: https://gd.eppo.int/taxon/RHYCFE/hosts (accessed on 14 January 2023). (In Italian).
- Qayyum, M.A.; Miao, Y.; Hou, Y. Integration of entomopathogenic fungi and eco-friendly insecticides for management of red palm weevil, Rhynchophorus ferrugineus (Olivier). Saudi J. Biol. Sci. 2019, 27, 1811–1817. [Google Scholar] [PubMed]
- Singh, S.P.; Rethinam, P. Trapping—A major tactic of BIPM strategy of palm weevils. Coconut Res. Dev. J. 2005, 21, 34. [Google Scholar] [CrossRef]
- Yuezhong, L.; Zeng-Rong, Z.; Ruiting, J.; Lian-Sheng, W. The red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), newly reported from Zhejiang, China and update of geographical distribution. Fla. Entomol. 2009, 92, 386–387. [Google Scholar] [CrossRef] [Green Version]
- Shaarawi, F.A.; Kamel, K.E.; Al Qahtani, A.M. Toxicity and histopathologic effects of two plant extracts, as compared to an insecticide, on the midgut of the fourth instar larvae of the red palm weevil, Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). J. Union Arab. Biol. 2000, 13, 491–503. [Google Scholar]
- Giblin-Davis, R.M.; Faleiro, J.R.; Jacas, J.A.; Peña, J.E.; Vidyasagar, P.S.P.V. Biology and Management of the Red Palm Weevil, Rhynchophorus ferrugineus. In Potential Invasive Pests of Agricultural Crops, 1st ed.; CABI: Wallingford, UK, 2013; p. 33. [Google Scholar]
- Darrag, H.M.; Almuhanna, H.T.; Hakami, E.H. Secondary Metabolites in Basil, Bio-Insecticide, Inhibition Effect, and In Silico Molecular Docking against Proteolytic Enzymes of the Red Palm Weevil (Rhynchophorus ferrugineus). Plants 2022, 11, 1087. [Google Scholar] [CrossRef]
- Shar, M.U.; Rustamani, M.A.; Nizamani, S.M.; Bhutto, L.A. Red palm weevil (Rhynchophorus ferrugineus Olivier) infestation and its chemical control in Sindh province of Pakistan. Afr. J. Agric. Res. 2012, 11, 1666–1673. [Google Scholar]
- Llácer, E.; Jacas, J.A. Efficacy of phosphine as a fumigant against Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in palms. Span. J. Agric. Res. 2010, 8, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Vidyasagar, P.; Bhat, S.K. Pest management in coconut gardens. J. Plant. Crops 1991, 19, 163–182. [Google Scholar]
- Aldawood, A. Guidelines on curative pesticide treatments (chemical trunk injection). In Red Palm Weevil: Guidelines on Management Practices; Elkakhy, M., Faleiro, J.R., Eds.; FAO: Rome, Italy, 2020. [Google Scholar]
- Hallett, R.H.; Gries, G.; Gries, R.; Borden, J.H.; Czyzewska, E.; Oehlschlager, A.C.; Pierce, H.D.; Angerilli, N.D.P.; Rauf, A. Aggregation pheromones of two Asian palm weevils, Rhynchophorus ferrugineus and R. vulneratus. Naturwissenschaften 1993, 80, 328–331. [Google Scholar] [CrossRef]
- Guarino, S.; Bue, P.L.; Peri, E.; Colazza, S. Responses of Rhynchophorus ferrugineus adults to selected synthetic palm esters: Electroantennographic studies and trap catches in an urban environment. Pest Manag. Sci. 2010, 67, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Abraham, V.A.; Al Shuaibi, M.A.; Faleiro, J.R.; Abuzuhairah, R.A.; Vidyasagar, P.S.P.V. An integrated management approach for the red palm weevil, Rhynchophorus ferrugineus Oliv., a key pest of date palm in the Middle East. Int. J. Sci. Res. Agric. Sci. 1998, 3, 77–84. [Google Scholar]
- Oehlschlager, A.C.; Chinchilla, C.M.; Gonzalez, L.M.; Jiron, L.F.; Mexzon, R.; Morgan, B. Development of a Pheromone-based trap for the American Palm Weevil, Rhynchophorus palmarum (L.). J. Econ. Entomol. 1993, 86, 1381–1392. [Google Scholar] [CrossRef]
- Nassar, M.M.I.; Abdullah, M.A. Evaluation of azadirachtin for control of the red palm weevil, Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). J. Egypt. Ger. Soc. Zool. 2001, 36, 163–173. [Google Scholar]
- Nurashikin-Khairuddin, W.; Abdul-Hamid, S.N.A.; Mansor, M.S.; Bharudin, I.; Othman, Z.; Jalinas, J. A Review of Entomopathogenic Nematodes as a Biological Control Agent for Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Insects 2022, 13, 245. [Google Scholar] [PubMed]
- Faleiro, J.R. A review of the issues and management of the red palm weevil. Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar]
- Mazza, G.; Inghilesi, A.F.; Stasolla, G.; Cini, A.; Cervo, R.; Benvenuti, C.; Francardi, V.; Cristofaro, M.; Arnone, S.; Roversi, P.F. Sterile Rhynchophorus ferrugineus males efficiently impair reproduction while maintaining their sexual competitiveness in a social context. J. Pest Sci. 2016, 89, 459–468. [Google Scholar]
- El Naggar, S.E.M.; Mohamed, H.F.; Mahmoud, E.A. Studies on the morphology and histology of the ovary of red palm weevil female irradiated with gamma rays. J. Asia Pac. Entomol. 2010, 13, 9–16. [Google Scholar]
- Paoli, F.; Dallai, R.; Cristofaro, M.; Arnone, C.S.; Francardi, V.; Roversi, P.F. Morphology of the male reproductive system, sperm ultrastructure and γirradiation of the red palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Dryophthoridae). Tissue Cell 2014, 46, 274–285. [Google Scholar] [CrossRef]
- Al-Ayedh, H.Y.; Rasool, K.G. Sex ratio and the role of mild relative humidity in mating behaviour of red date palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae) gamma-irradiated adults. J. Appl. Entomol. 2010, 134, 157–162. [Google Scholar] [CrossRef]
- Al-Ayedh, H.Y.; Rasool, K.G. Determination of the optimum sterilizing radiation dose for control of the red date palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae). Crop Prot. 2010, 29, 1377–1380. [Google Scholar] [CrossRef]
- Massa, R.; Migliore, M.D.; Panariello, G.; Pinchera, D.; Schettino, F.; Caprio, E.; Griffo, R. Wide Band Permittivity Measurements of Palm (Phoenix canariensis) and Rhynchophorus ferrugineus (Coleoptera: Curculionidae) for RF Pest Control. J. Microw. Power Electromagn. Energy 2014, 3, 158–169. [Google Scholar] [CrossRef]
- Massa, R.; Panariello, G.; Migliore, M.D.; Pinchera, D.; Schettino, F.; Griffo, R.; Martano, M.; Power, K.; Maiolino, P.; Caprio, E. Microwave heating: A promising and eco-compatible solution to fight the spread of red palm weevil. Arab. J. Plant Prot. 2019, 37, 143–148. [Google Scholar] [CrossRef]
- Mady, H.Y.; Ahmed, M.M.; El Namaky, A.H. Electromagnetic wave and Microwave heating: An eco-compatible solution to control Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). J. Biopestic. 2021, 14, 132–140. [Google Scholar]
- Rmili, H.; Alkhalifeh, K.; Zarouan, M.; Zouch, W.; Islam, M.T. Numerical analysis of the microwave treatment of palm trees infested with the red palm weevil pest by using a circular array of vivaldi antennas. IEEE Access 2020, 8, 152342–152350. [Google Scholar] [CrossRef]
- Massa, R.; Panariello, G.; Pinchera, D.; Schettino, F.; Caprio, E.; Griffo, R.; Migliore, M.D. Experimental and numerical evaluations on palm microwave heating for Red Palm Weevil pest control. Sci. Rep. 2017, 7, 45299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massa, R.; Greco, A.; Caprio, E.; Panariello, G.; Migliore, M.D.; Pinchera, D.; Schettino, F.; Griffo, R. Experimental Results on the Effectiveness of Microwave Treatment of Phoenix Canariensis Palm Infested by Rhynchophorus ferrugineus. In Proceedings of the 2015 IEEE 15th Mediterranean Microwave Symposium (MMS), Lecce, Italy, 30 November–2 December 2015. [Google Scholar]
- Romeo, S.; D’Avino, C.; Pinchera, D.; Zeni, O.; Scarfì, M.R.; Massa, R. A waveguide applicator for in vitro exposures to single or multiple ICT frequencies. IEEE Trans. Microw. Theory Tech. 2013, 61, 1994–2004. [Google Scholar] [CrossRef]
- Kamel, K.E.; Mohamed, M.I.; Shaarawi, F.A. Histology and ultrastructure of the ovary of the red palm weevil, Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). J. Egypt. Ger. Soc. Zool. 2005, 47, 87–105. [Google Scholar]
- Beckett, S.J.; Fields, P.G.; Subramanyam, B.H. Disinfestation of stored products and associated structures using heat. In Heat Treatments for Post-Harvest Pest Control: Theory and Practice; Tang, J., Mitcham, E., Wang, S., Lurie, S., Eds.; CABI: Wallingford, UK, 2007; pp. 182–237. [Google Scholar]
- Hansen, J.D.; Johnson, J.A.; Winter, D.A. History and use of heat in pest control: A review. Int. J. Pest Manag. 2011, 57, 267–289. [Google Scholar]
- Massa, R.; Caprio, E.; De Santis, M.; Griffo, R.; Migliore, M.D.; Panariello, G.; Pinchera, D.; Spigno, P. Microwave treatment for pest control: The case of Rhynchophorus ferrugineus in Phoenix canariensis. EPPO Bull. 2011, 41, 128–135. [Google Scholar]
- Nelson, S.O. Review and assessment of radio-frequency and microwave energy for stored-grain insect control. Trans. ASAE 1996, 39, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.O.; Stetson, L.E.; Rhine, J.J. Factors influencing effectiveness of radiofrequency electric fields for stored-grain insect control. Trans. Am. Soc. Agric. Eng. 1966, 9, 809–0815. [Google Scholar] [CrossRef]
- Rai, R.S.; Ball, H.J.; Nelson, S.O.; Stetson, L.E. Cytopathological effects of radiofrequency electric fields on reproductive tissue of adult Teneberio molitor (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 1974, 67, 687–690. [Google Scholar] [CrossRef]
- Blanckenhorn, W.U.; Henseler, C. Temperature-dependent ovariole and testis maturation in the yellow dung fly. Entomol. Exp. Et Appl. 2005, 116, 159–165. [Google Scholar] [CrossRef]
- El-Bokl, M.M.; Baker, R.F.A.; El-Gammal, H.L.; Mahmoud, M.Z. Biological and histopathological effects of some insecticidal agents against red palm weevil Rhynchophorus ferrugineus. Egypt. Acad. J. Biol. Sci. 2010, 1, 7–22. [Google Scholar] [CrossRef]
- MAl-Dhafar, Z.M.; Sharaby, A. Effect of Zinc Sulfate against the Red Palm Weevil Rhynchophorus ferrugineus with Reference to Their Histological Changes on the Larval Midgut and Adult Reproductive System. J. Agric. Sci. Technol. 2012, 2, 888–900. [Google Scholar]
- Peng, L.; Miao, Y.; Hou, Y. Demographic comparison and population projection of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) reared on sugarcane at different temperatures. Sci. Rep. 2016, 6, 31659. [Google Scholar] [CrossRef]
- Mustapha, E.B.; Jose Romeno Hilario, F.D.S. Date Palm Pests and Diseases: Integrated Management Guide; International Center for Agricultural Research in the Dry Areas (ICARDA): Beirut, Lebanon, 2018. [Google Scholar]
- Massa, R.; Schettino, F.; Panariello, G.; Migliore, M.D.; Pinchera, D.; Chirico, G.; D’Silva, C.J.; Griffo, R.; Yaseen, T. Using Microwave Technology against Red Palm Weevil: An Innovative Sustainable Strategy to Contrast a Lethal Pest of Date Palms. Acta Hortic. 2023, in press.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sex | Irradiation Time | ΔT [°C] | Survival Days | Score of Histological Lesions |
|---|---|---|---|---|---|
| 1 | F | UNTREATED | 23 | 0 | |
| 2 | F | UNTREATED | 30 | 0 | |
| 3 | M | UNTREATED | 43 | 0 | |
| 4 | M | UNTREATED | 71 | 0 | |
| 5 | F | 5 s. | 8.9 | 40 | 1 |
| 6 | F | 5 s. | 9.5 | 28 | ½ |
| 7 | F | 5 s. | 9.9 | 7 | 1 |
| 8 | F | 5 s. | 7.6 | 11 | 1 |
| 9 | M | 5 s. | 14.4 | 7 | 2 |
| 10 | M | 5 s. | 5.7 | 12 | 1 |
| 11 | M | 5 s. | 10 | 14 | 1 |
| 12 | M | 5 s. | 10 | 54 | 2 |
| 13 | F | 15 s. | 14.5 | 3 | 2 |
| 14 | F | 15 s. | 26.2 | 3 | 3 |
| 15 | F | 15 s. | 7.9 | 12 | 3 |
| 16 | F | 15 s. | 9.3 | 32 | 2 |
| 17 | M | 15 s. | 13.6 | 32 | 2 |
| 18 | M | 15 s. | 16.9 | 7 | 2 |
| 19 | M | 15 s. | 6.8 | 43 | 1 |
| 20 | M | 15 s. | 16.6 | 10 | 3 |
| 21 | F | 30 s. | 22 | 16 | 3 |
| 22 | F | 30 s. | 26.6 | 28 | 3 |
| 23 | F | 30 s. | 22.4 | 12 | 3 |
| 24 | F | 30 s. | 26 | 13 | 3 |
| 25 | M | 30 s. | 24.6 | 21 | 3 |
| 26 | M | 30 s. | 16.7 | 21 | 2/3 |
| 27 | M | 30 s. | 24.8 | 21 | 3 |
| 28 | M | 30 s. | 13.2 | 14 | 3 |
| 29 | M | 30 s. | 7.4 | 1 | Not assessed |
| Number of Samples (Sex) | Irradiation Time | ΔT [°C] (Mean ± SD) | Survival Days (Mean ± SD) | Score of Histological Lesions |
|---|---|---|---|---|
| 4 (2 females and 2 males) | Untreated | 41.75 ± 21.18 | 0 | |
| 8 (4 females and 4 males) | 5 s. | 9.5 ± 2.4 | 21.62 ± 17.39 | 1 |
| 8 (4 females and 4 males) | 15 s. | 13.9 ± 6.2 | 17.75 ± 15.52 | 2 |
| 8 (4 females and 4 males) | 30 s. | 20.4 1 ± 6.5 | 16.03 ± 7.6 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martano, M.; Massa, R.; Restucci, B.; Caprio, E.; Griffo, R.; Power, K.; Maiolino, P. Microwaves Induce Histological Alteration of Ovaries and Testis in Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae). Agronomy 2023, 13, 420. https://doi.org/10.3390/agronomy13020420
Martano M, Massa R, Restucci B, Caprio E, Griffo R, Power K, Maiolino P. Microwaves Induce Histological Alteration of Ovaries and Testis in Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae). Agronomy. 2023; 13(2):420. https://doi.org/10.3390/agronomy13020420
Chicago/Turabian StyleMartano, Manuela, Rita Massa, Brunella Restucci, Emilio Caprio, Raffaele Griffo, Karen Power, and Paola Maiolino. 2023. "Microwaves Induce Histological Alteration of Ovaries and Testis in Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae)" Agronomy 13, no. 2: 420. https://doi.org/10.3390/agronomy13020420