
Distribution Characteristics of Microbial Residues within Aggregates of Fluvo-Aquic Soil under Biochar Application
Abstract
:
1. Introduction
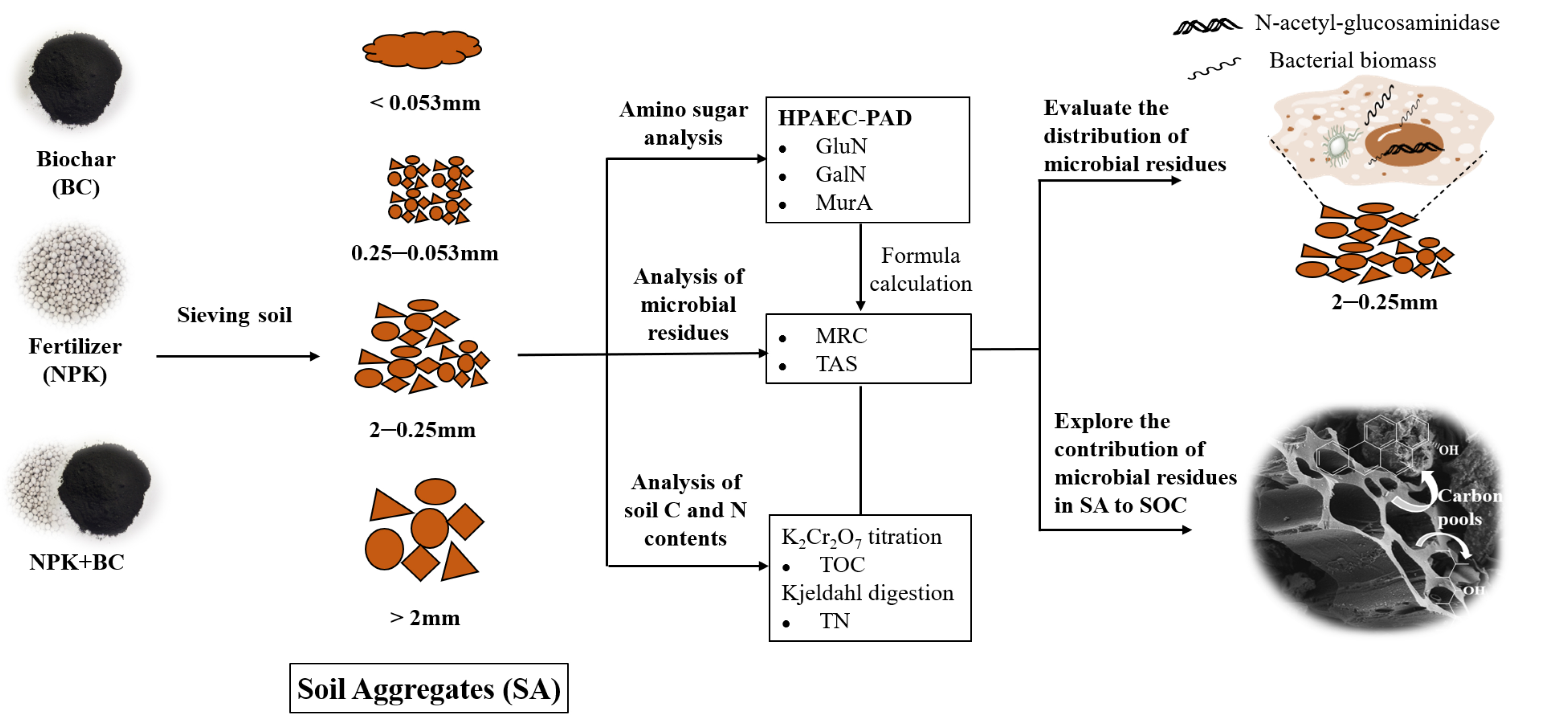
2. Materials and Methods
2.1. Soil and Biochar
2.2. Experimental Design
2.3. Soil Sample Harvesting and Aggregate Classification
2.4. Soil C- and N-Related Properties and Amino Sugar Extraction
2.5. Data and Statistical Analysis
3. Results
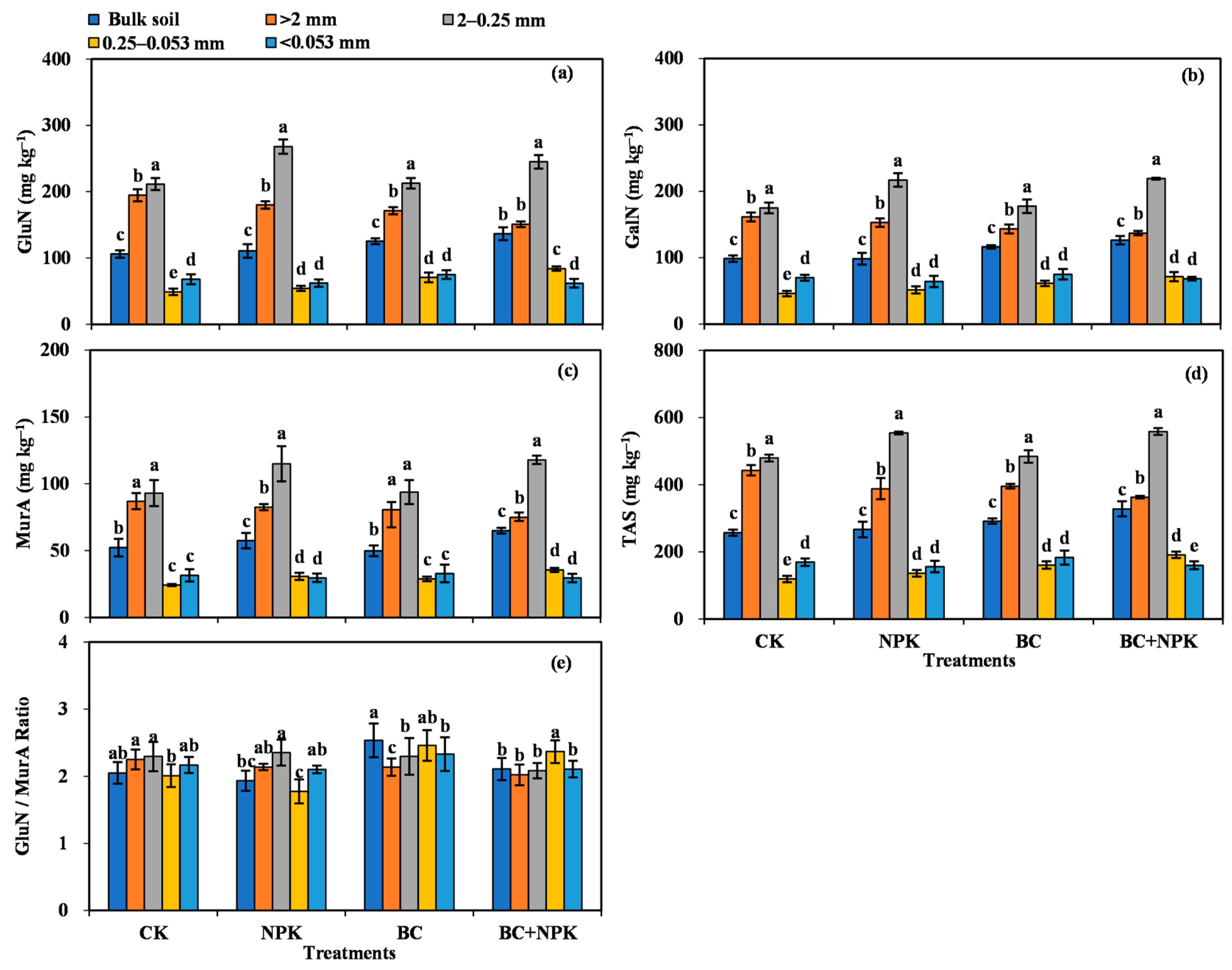
3.1. Amino Sugar Content in Bulk Soil and SA
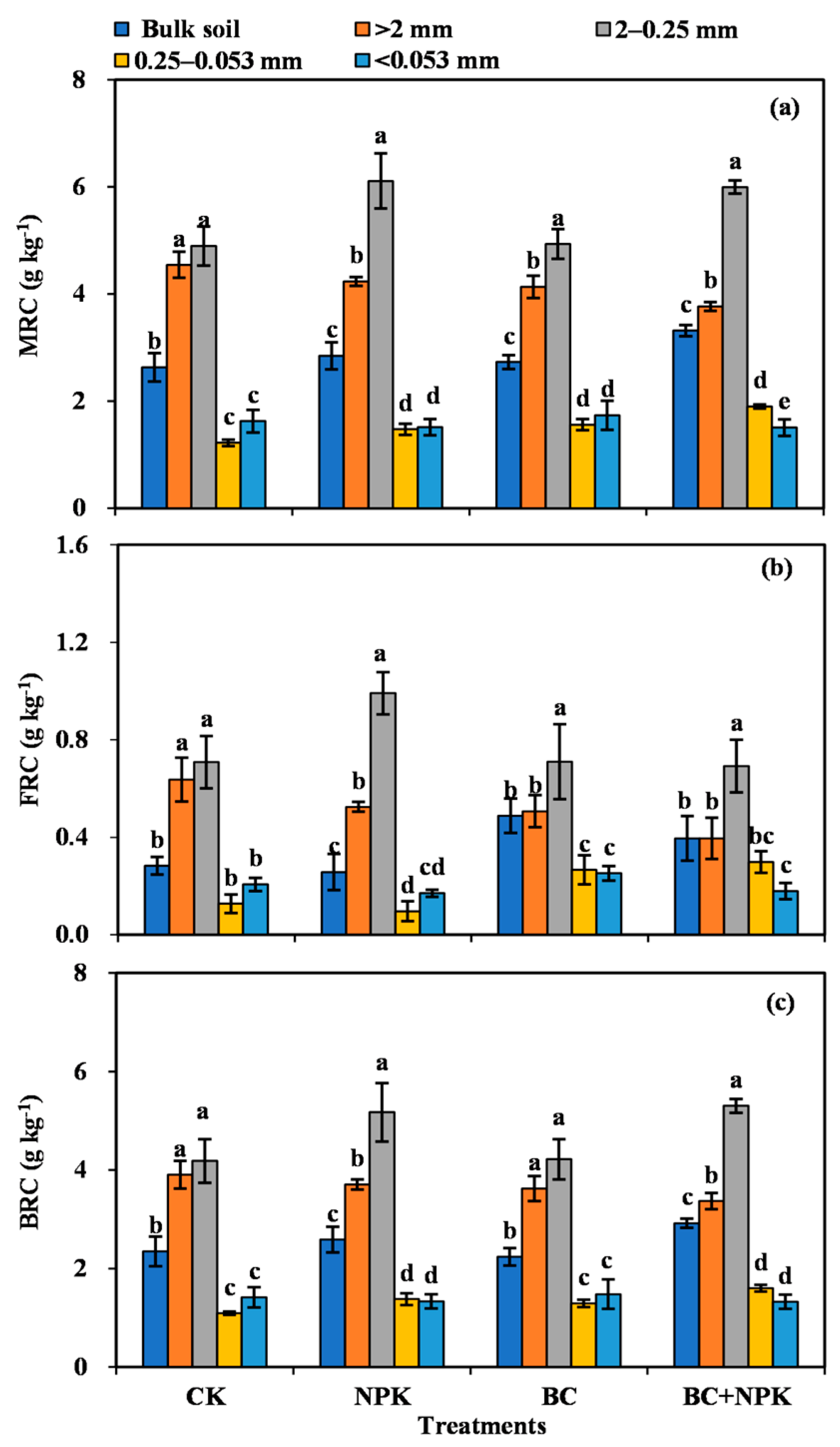
3.2. Content of MRC in Bulk Soil and SA
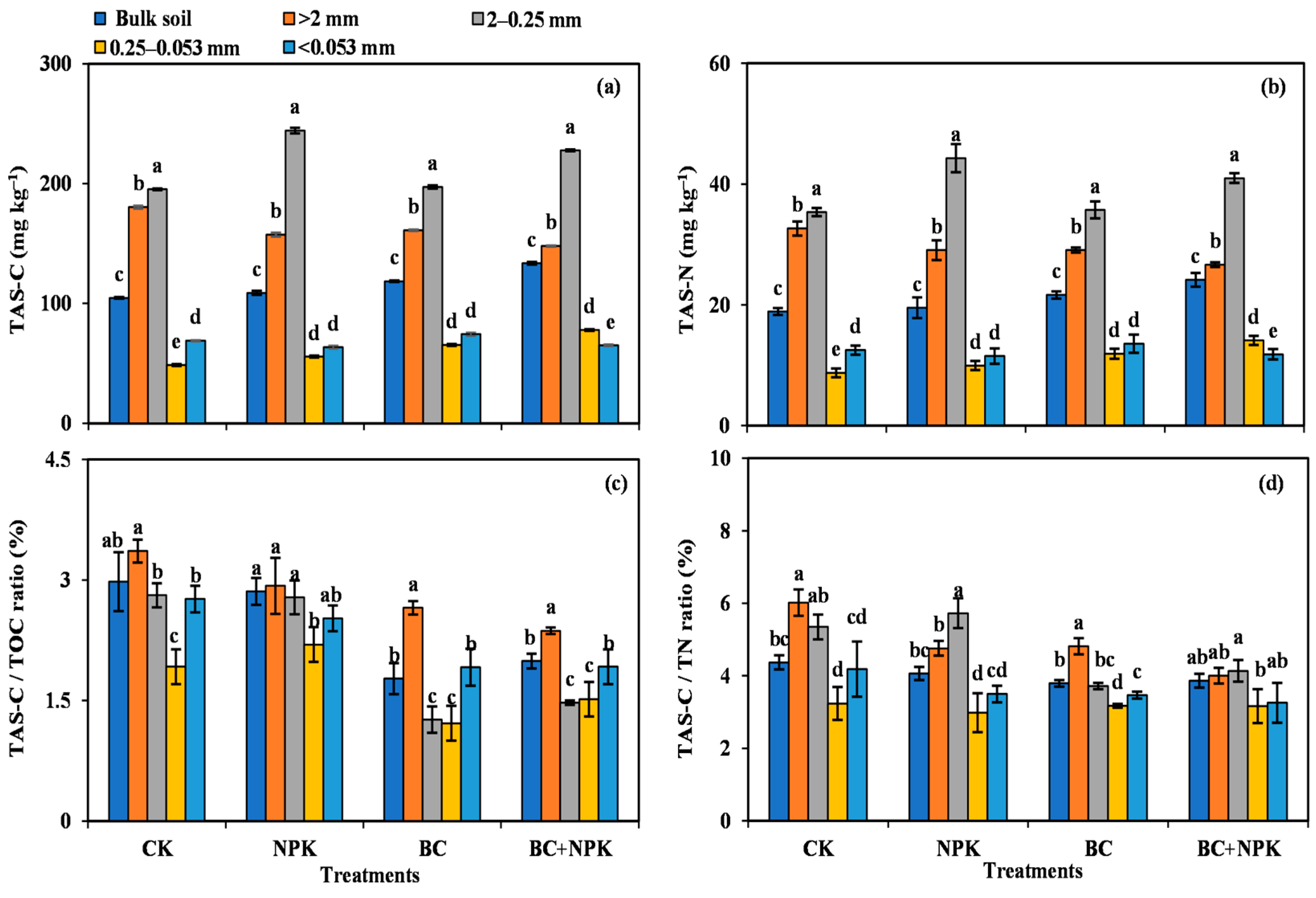
3.3. TAS-C and TAS-N Contents and Contribution to TOC and TN
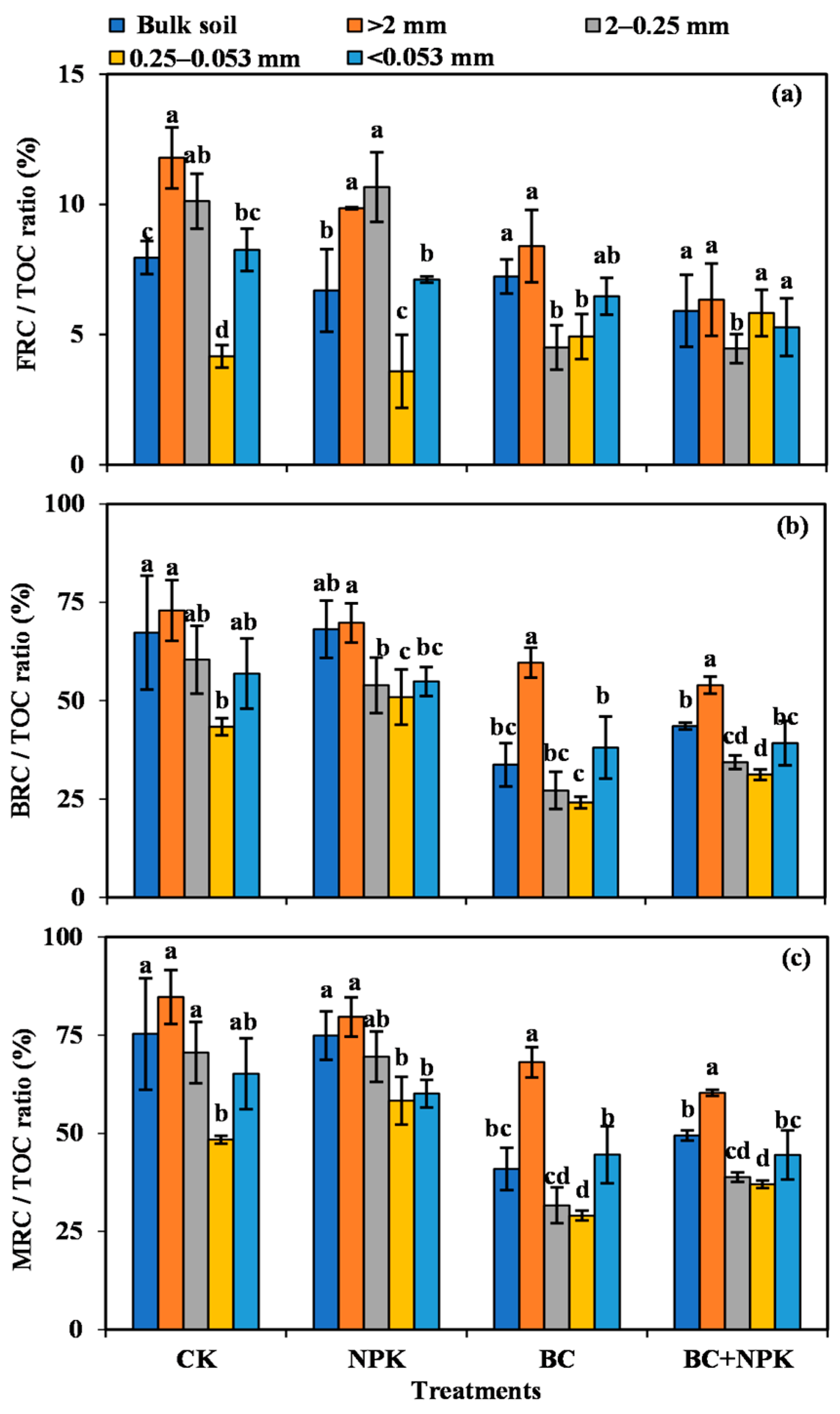
3.4. Ratios of FRC, BRC, and MRC to TOC
3.5. Association between Microbial Residues and Soil Properties
4. Discussion
4.1. Distribution of Microbial Residues within the SA
4.2. Effects of the Application of Biochar and Fertilizer on the Response of Microbial Residues in SA
4.3. Contribution of Microbial Residues to the SOC within SA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Cui, J.; Wu, H.; Zheng, Q.; Song, D.; Wang, X.; Zhang, S. Organic carbon, total nitrogen, and microbial community distributions within aggregates of calcareous soil treated with biochar. Agric. Ecosyst. Environ. 2021, 314, 107408. [Google Scholar] [CrossRef]
- Chai, Y.J.; Zeng, X.B.; E, S.Z.; Bai, L.Y.; Su, S.M.; Huang, T. Effects of freeze-thaw on aggregate stability and the organic carbon and nitrogen enrichment ratios in aggregate fractions. Soil Use Manag. 2014, 30, 507–516. [Google Scholar] [CrossRef]
- Angst, G.; Mueller Kevin, E.; Nierop Klaas, G.J.; Simpson Myrna, J. Plant-or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Ma, S.; Chen, G.; Du, E.; Tian, D.; Xing, A.; Shen, H.; Ji, C.; Zheng, C.; Zhu, J.; Zhu, J.; et al. Effects of nitrogen addition on microbial residues and their contribution to soil organic carbon in China’s forests from tropical to boreal zone. Environ. Pollut. 2020, 268, 115941. [Google Scholar] [CrossRef]
- Ni, X.; Liao, S.; Tan, S.; Wang, D.; Peng, Y.; Yue, K.; Wu, F.; Yang, Y. A quantitative assessment of amino sugars in soil profiles. Soil Biol. Biochem. 2020, 143, 107762. [Google Scholar] [CrossRef]
- Li, L.; Wilson, C.B.; He, H.; Zhang, X.; Zhou, F.; Schaeffer, S.M. Physical, biochemical, and microbial controls on amino sugar accumulation in soils under long-term cover cropping and no-tillage farming. Soil Biol. Biochem. 2019, 135, 369–378. [Google Scholar] [CrossRef]
- Ding, X.; Liang, C.; Zhang, B.; Yuan, Y.; Han, X. Higher rates of manure application lead to greater accumulation of both fungal and bacterial residues in macroaggregates of a clay soil. Soil Biol. Biochem. 2015, 84, 137–146. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and Fungal Contributions to Carbon Sequestration in Agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Amelung, W.; Miltner, A.; Zhang, X.; Zech, W. Fate of microbial residues during litter decomposition as affected by minerals. Soil Sci. 2001, 166, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Murugan, R.; Djukic, I.; Keiblinger, K.; Zehetner, F.; Bierbaumer, M.; Zechmeister-Bolternstern, S.; Joergernsen, R.G. Spatial distribution of microbial biomass and residues across soil aggregate fractions at different elevations in the Central Austrian Alps. Geoderma 2019, 339, 1–8. [Google Scholar] [CrossRef]
- Joergensen, R.G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biol. Fert. Soils 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Hu, G.; Liu, X.; He, H.; Chen, W.; Zhuge, Y.; Dong, Y.; Wang, H. Accumulation Characteristics of Amino Sugars in Salinized Soils of Different Types in the Yellow River Delta. Acta Pedol. Sin. 2018, 55, 390–398. (In Chinese) [Google Scholar]
- Luan, H.; Yuan, S.; Gao, W.; Tang, J.; Li, R.; Zhang, H.; Huang, S. Aggregate-related changes in living microbial biomass and microbial necromass associated with different fertilization patterns of greenhouse vegetable soils. Eur. J. Soil Biol. 2021, 103, 103291. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, F.; Hu, G.; Shao, S.; He, H.; Zhang, W.; Zhang, X.; Li, L. Dynamic contribution of microbial residues to soil organic matter accumulation influenced by maize straw mulching. Geoderma 2019, 333, 35–42. [Google Scholar] [CrossRef]
- Wang, X.; Eltohamy, K.M.; Liu, C.; Li, F.; Fang, Y.; Kawasaki, A.; Liang, X. Biochar reduces colloidal phosphorus in soil aggregates: The role of microbial communities. J. Environ. Manag. 2023, 326, 116745. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Xi, X.; Zheng, Q.; Liang, G.; Zhou, W.; Wang, X. Soil nutrient and microbial activity responses to two years after maize straw biochar application in a calcareous soil. Ecotoxicol. Environ. Saf. 2019, 180, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Fang, H.; Zhang, Q.; Wang, Q.; Chen, C.; Mooney, S.J.; Peng, X.; Du, Z. Biochar enhances soil hydraulic function but not soil aggregation in a sandy loam. Eur. J. Soil Sci. 2019, 70, 291–300. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, Y.; Wu, Z.; Yan, X.; Gunina, A.; Kuzyakov, Y.; Xiong, Z. Effects of six-year biochar amendment on soil aggregation, crop growth, and nitrogen and phosphorus use efficiencies in a rice-wheat rotation. J. Clean. Prod. 2020, 242, 118435. [Google Scholar] [CrossRef]
- Obia, A.; Mulder, J.; Martinsen, V.; Cornelissen, G.; Børresen, T. In situ effects of biochar on aggregation, water retention and porosity in light-textured tropical soils. Soil Till. Res. 2016, 155, 35–44. [Google Scholar] [CrossRef]
- Dai, H.; Chen, Y.; Liu, K.; Li, Z.; Qian, X.; Zang, H.; Yang, X.; Zhao, Y.; Shen, Y.; Li, Z.; et al. Water-stable aggregates and carbon accumulation in barren sandy soil depend on organic amendment method: A three-year field study. J. Clean. Prod. 2019, 212, 393–400. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Q.; Zhan, L.; Xu, X.; Bi, R.; Xiong, Z. Biochar addition stabilized soil carbon sequestration by reducing temperature sensitivity of mineralization and altering the microbial community in a greenhouse vegetable field. J. Environ. Manag. 2022, 313, 114972. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, H.; Chen, X.; Zhang, C.; Ma, W.; Huang, C.; Zhang, W.; Mi, G.; Miao, Y.; Li, X.; et al. Pursuing sustainable productivity with millions of smallholder farmers. Nature 2018, 555, 363–366. [Google Scholar] [CrossRef]
- World Reference Base of Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Recourses Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Zhang, W.L.; Xu, A.G.; Zhang, R.L.; Ji, H.J. Review of Soil Classification and Revision of China Soil Classification System. Sci. Agric. Sin. 2014, 47, 3214–3230. (In Chinese) [Google Scholar]
- Liao, H.; Zhang, Y.; Wang, K.; Hao, X.; Chen, W.; Huang, Q. Complexity of bacterial and fungal network increases with soil aggregate size in an agricultural Inceptisol. Appl. Soil Ecol. 2020, 154, 103640. [Google Scholar] [CrossRef]
- Manna, M.C.; Swarup, A.; Wanjari, R.H.; Mishra, B.; Shahi, D.K. Long-term fertilization, manure and liming effects on soil organic matter and crop yields. Soil Till. Res. 2006, 94, 397–409. [Google Scholar] [CrossRef]
- Juliana, H.S.; Cícero, C.D.F.; Robélio, L.M.; Madari, B.E.M.; Luiz, E.C.B.; Jader, G.B.; Diego, M.D.S. Methods of soil organic carbon determination in Brazilian savannah soils. Sci. Agric. 2014, 71, 302–308. [Google Scholar]
- Skjemstad, J.O.; Reeve, R. The determination of nitrogen in soils by rapid high-temperature Kjeldahl digestion and autoanalysis. Commun. Soil Sci. Plant Anal. 2008, 7, 229–239. [Google Scholar] [CrossRef]
- Indorf, C.; Dyckmans, J.; Khan, K.S.; Joergensen, R.G. Optimisation of amino sugar quantification by HPLC in soil and plant hydrolysates. Biol. Fertil. Soils 2011, 47, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liang, C.; Duan, X.; Chen, H.; Li, D. Variation of microbial residue contribution to soil organic carbon sequestration following land use change in a subtropical karst region. Geoderma 2019, 353, 340–346. [Google Scholar] [CrossRef]
- Luan, H.; Yuan, S.; Gao, W.; Tang, J.; Li, R.; Zhang, H.; Huang, S. 10-Year fertilization alters soil C dynamics as indicated by amino sugar differentiation and oxidizable organic C pools in a greenhouse vegetable field of Tianjin, China. Appl. Soil Ecol. 2022, 169, 104226. [Google Scholar] [CrossRef]
- Faust, S.; Heinze, S.; Ngosong, C.; Sradnick, A.; Oltmanns, M.; Raupp, J.; Geisseler, D.; Joergensen, R.G. Effect of biodynamic soil amendments on microbial communities in comparison with inorganic fertilization. Appl. Soil Ecol. 2017, 114, 82–89. [Google Scholar] [CrossRef]
- Sun, H.; Koal, P.; Liu, D.; Gerl, G.; Schroll, R.; Gattinger, A.; Joergensen, R.G.; Munch, J.C. Soil microbial community and microbial residues respond positively to minimum tillage under organic farming in Southern Germany. Appl. Soil Ecol. 2016, 108, 16–24. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Wang, B.; An, S.; Liang, C.; Liu, Y.; Kuzyakov, Y. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biol. Biochem. 2021, 162, 108422. [Google Scholar] [CrossRef]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil -concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Hu, G.; Zhao, Y.; Liu, X.; Zhou, F.; Zhang, W.; Shao, S.; He, H.; Zhang, X. Comparing microbial transformation of maize residue-N and fertilizer-N in soil using amino sugar-specific 15N analysis. Eur. J. Soil Sci. 2020, 71, 252–264. [Google Scholar] [CrossRef]
- Ding, X.; Han, X. Effects of long-term fertilization on contents and distribution of microbial residues within aggregate structures of a clay soil. Biol. Fert. Soils 2014, 50, 549–554. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, J.; Yang, F.; Dong, W.; Dai, X.; Yang, Y.; Sun, X. Specific Responses of Soil Microbial Residue Carbon to Long-Term Mineral Fertilizer Applications to Reddish Paddy Soils. Pedosphere 2018, 28, 488–496. [Google Scholar] [CrossRef]
- Chen, X.; Hu, Y.; Xia, Y.; Zheng, S.; Ma, C.; Rui, Y.; He, H.; Huang, D.; Zhang, Z.; Ge, T.; et al. Contrasting pathways of carbon sequestration in paddy and upland soils. Glob. Chang. Biol. 2021, 27, 2478–2490. [Google Scholar] [CrossRef]
- Wang, C.; Chen, D.; Shen, J.; Yuan, Q.; Fan, F.; Wei, W.; Li, Y.; Wu, J. Biochar alters soil microbial communities and potential functions 3–4 years after amendment in a double rice cropping system. Agr. Ecosyst. Environ. 2021, 311, 107291. [Google Scholar] [CrossRef]
- Bicharanloo, B.; Shirvan, M.B.; Keitel, C.; Dijkstra, F.A. Rhizodeposition mediates the effect of nitrogen and phosphorous availability on microbial carbon use efficiency and turnover rate. Soil Biol. Biochem. 2020, 142, 107705. [Google Scholar] [CrossRef]
- Spohn, M.; Klaus, K.; Wanek, W.; Richter, A. Microbial carbon use efficiency and biomass turnover times depending on soil depth—Implications for carbon cycling. Soil Biol. Biochem. 2016, 96, 74–81. [Google Scholar] [CrossRef]
- Davinic, M.; Fultz, L.M.; Acosta-Martinez, V.; Calderón, F.J.; Cox, S.B.; Dowd, S.E.; Allen, V.G.; Zak, J.C.; Moore-Kucera, J. Pyrosequencing and mid-infrared spectroscopy reveal distinct aggregate stratification of soil bacterial communities and organic matter composition. Soil Biol. Biochem. 2012, 46, 63–72. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, X.; Zheng, X.; Deng, S.; Hu, Y.; Zheng, S.; He, X.; Wu, J.; Kuzyakov, Y.; Su, Y. Preferential uptake of hydrophilic and hydrophobic compounds by bacteria and fungi in upland and paddy soils. Soil Biol. Biochem. 2020, 148, 107879. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Chang. Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Brabcová, V.; Štursová, M.; Davidová, A.; Jansa, J.; Cajthaml, T.; Baldrian, P. Decomposer food web in a deciduous forest shows high share of generalist microorganisms and importance of microbial biomass recycling. ISME J. 2018, 12, 1768–1778. [Google Scholar] [CrossRef] [Green Version]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Indices of Microbial Residues | Fertilization | Aggregate Particle Sizes | Fertilization × Aggregate Particle Sizes | ||||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| Amino sugars | GluN | 0.951 | 0.428 | 324.313 | <0.001 | 4.968 | <0.001 |
| GalN | 1.519 | 0.228 | 430.981 | <0.001 | 5.097 | <0.001 | |
| MurA | 1.795 | 0.168 | 246.366 | <0.001 | 3.366 | 0.005 | |
| TAS | 2.007 | 0.133 | 539.885 | <0.001 | 7.134 | <0.001 | |
| GluN/MurA | 2.786 | 0.057 | 1.671 | 0.193 | 2.019 | 0.070 | |
| Microbial residue carbon | FRC | 0.415 | 0.743 | 70.749 | <0.001 | 2.191 | 0.050 |
| BRC | 1.795 | 0.168 | 246.366 | <0.001 | 3.366 | 0.005 | |
| MRC | 1.658 | 0.196 | 322.604 | <0.001 | 4.422 | 0.001 | |
| FRC/BRC | 2.786 | 0.057 | 1.671 | 0.193 | 2.019 | 0.070 | |
| Bulk Soil Properties | The Indices of Microbial Residues | ||||||
|---|---|---|---|---|---|---|---|
| Amino Sugars | Microbial Residue Carbon | ||||||
| GluN | GalN | MurA | TAS | FRC | BRC | MRC | |
| MWD | 0.805 ** | 0.681 * | 0.614 * | 0.812 ** | 0.382 | 0.614 * | 0.772 ** |
| R0.25 | 0.972 ** | 0.907 ** | 0.465 | 0.953 ** | 0.695 * | 0.465 | 0.727 ** |
| TOC | 0.779 ** | 0.777 ** | 0.096 | 0.721 ** | 0.790 ** | 0.221 | 0.378 |
| TN | 0.930 ** | 0.941 ** | 0.344 | 0.921 ** | 0.749 ** | 0.284 | 0.621 * |
| C/N | 0.521 | 0.514 | −0.108 | 0.439 | 0.673 * | −0.108 | 0.125 |
| MBC | 0.829 ** | 0.785 ** | 0.553 | 0.842 ** | 0.460 | 0.517 | 0.736 ** |
| MBN | 0.698 * | 0.551 | 0.774 ** | 0.743 ** | 0.128 | 0.720 ** | 0.848 ** |
| The Indices of Microbial Residues | Soil Physicochemical Properties | p | R2 | |
|---|---|---|---|---|
| Amino sugars | GluN | +Bacteria, +NAG, −βX, +TN, −TOC | <0.001 | 0.907 |
| GalN | +Bacteria, +NAG, −βX, +TN, −TOC,+αG, −Fungi, −U | <0.001 | 0.927 | |
| MurA | +Bacteria, +NAG, −βX | <0.001 | 0.885 | |
| TAS | +Bacteria, +NAG, −βX | <0.001 | 0.896 | |
| Microbial residue carbon | FRC | +Actinomycetes, +NAG, −Fungi, −U | <0.001 | 0.768 |
| BRC | +Bacteria, +NAG, −βX | <0.001 | 0.885 | |
| MRC | +Bacteria, +NAG, −βX | <0.001 | 0.895 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Zhang, S.; Song, D.; Wu, H.; Wang, L.; Wang, X. Distribution Characteristics of Microbial Residues within Aggregates of Fluvo-Aquic Soil under Biochar Application. Agronomy 2023, 13, 392. https://doi.org/10.3390/agronomy13020392
Cheng Y, Zhang S, Song D, Wu H, Wang L, Wang X. Distribution Characteristics of Microbial Residues within Aggregates of Fluvo-Aquic Soil under Biochar Application. Agronomy. 2023; 13(2):392. https://doi.org/10.3390/agronomy13020392
Chicago/Turabian StyleCheng, Yuyang, Shuai Zhang, Dali Song, Hang Wu, Linxuan Wang, and Xiubin Wang. 2023. "Distribution Characteristics of Microbial Residues within Aggregates of Fluvo-Aquic Soil under Biochar Application" Agronomy 13, no. 2: 392. https://doi.org/10.3390/agronomy13020392