
Biofertilization with PGP Bacteria Improve Strawberry Plant Performance under Sub-Optimum Phosphorus Fertilization

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Setup
2.2. Bacteria Strain Selection, Characteristics and Inoculum Preparation
2.3. Plant Growth and Physiological Traits Analysis
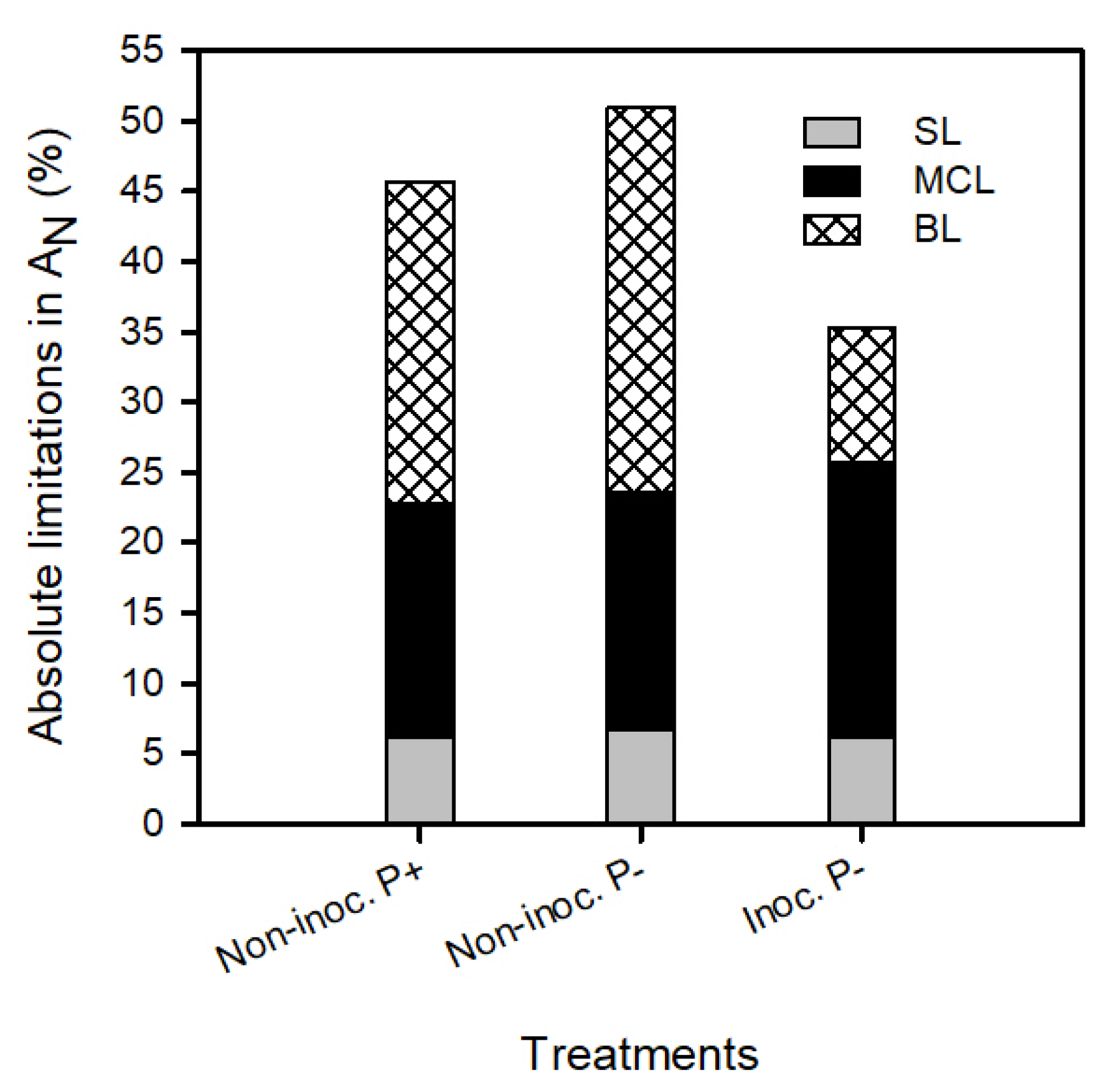
2.4. Quantitative Analysis of Leaf Photosynthetic Limitations
2.5. Ion Concentration in Leaves and Roots
2.6. Statistical Analysis
3. Results
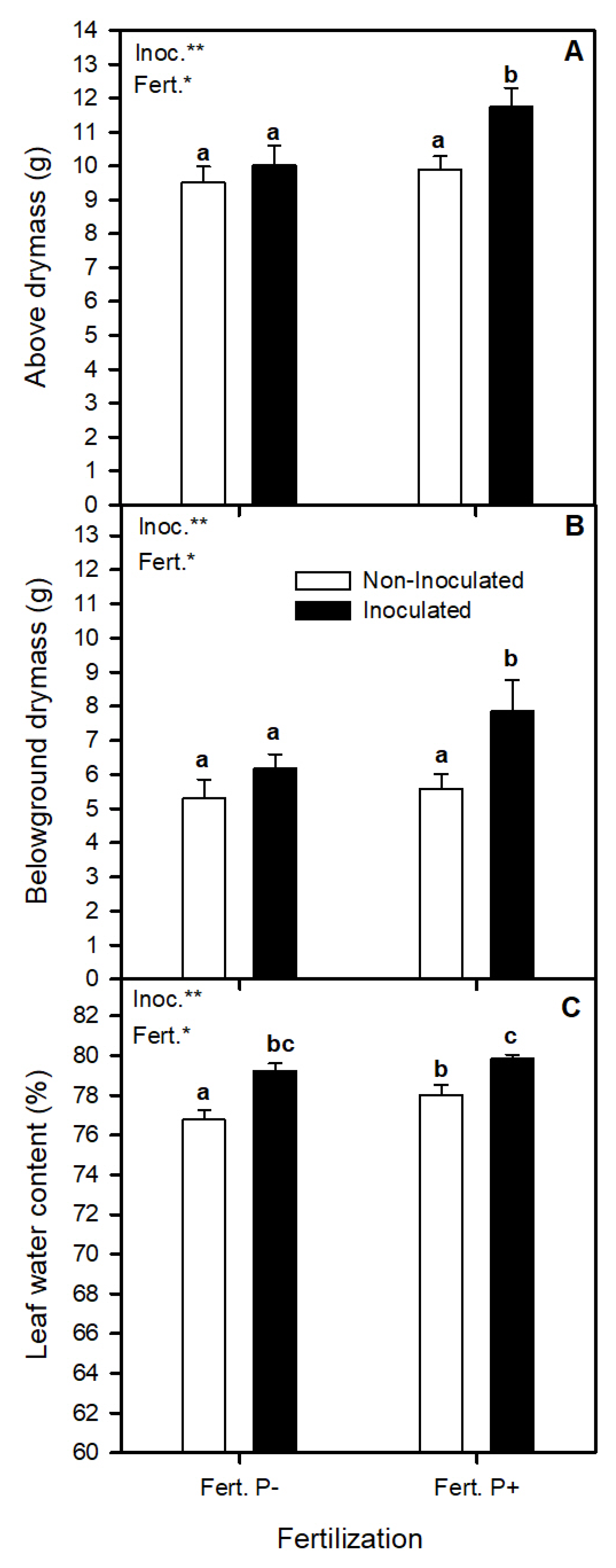
3.1. Bacterial Inoculation and Phosphorus Fertilization Availability Effect on Strawberry Growth
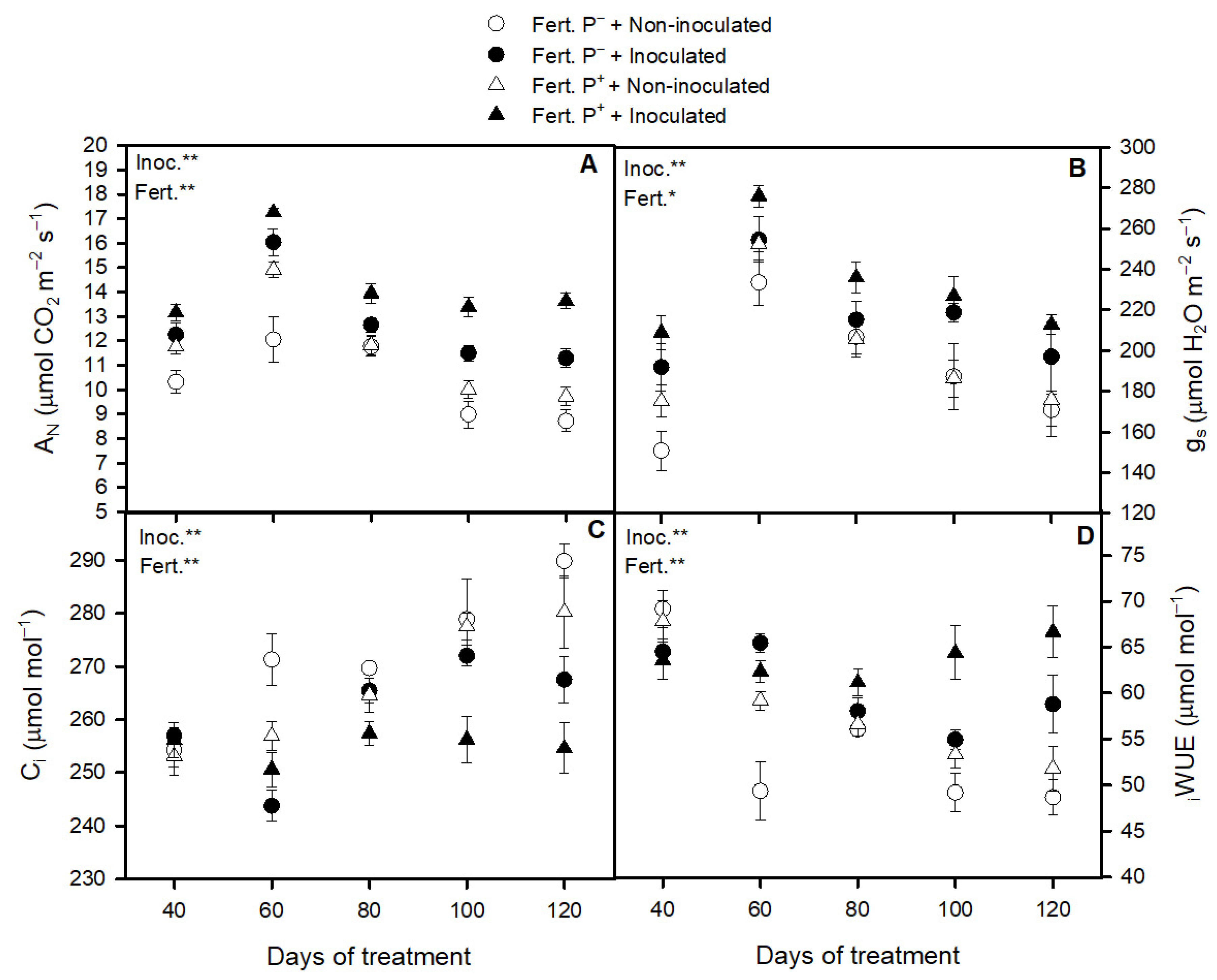
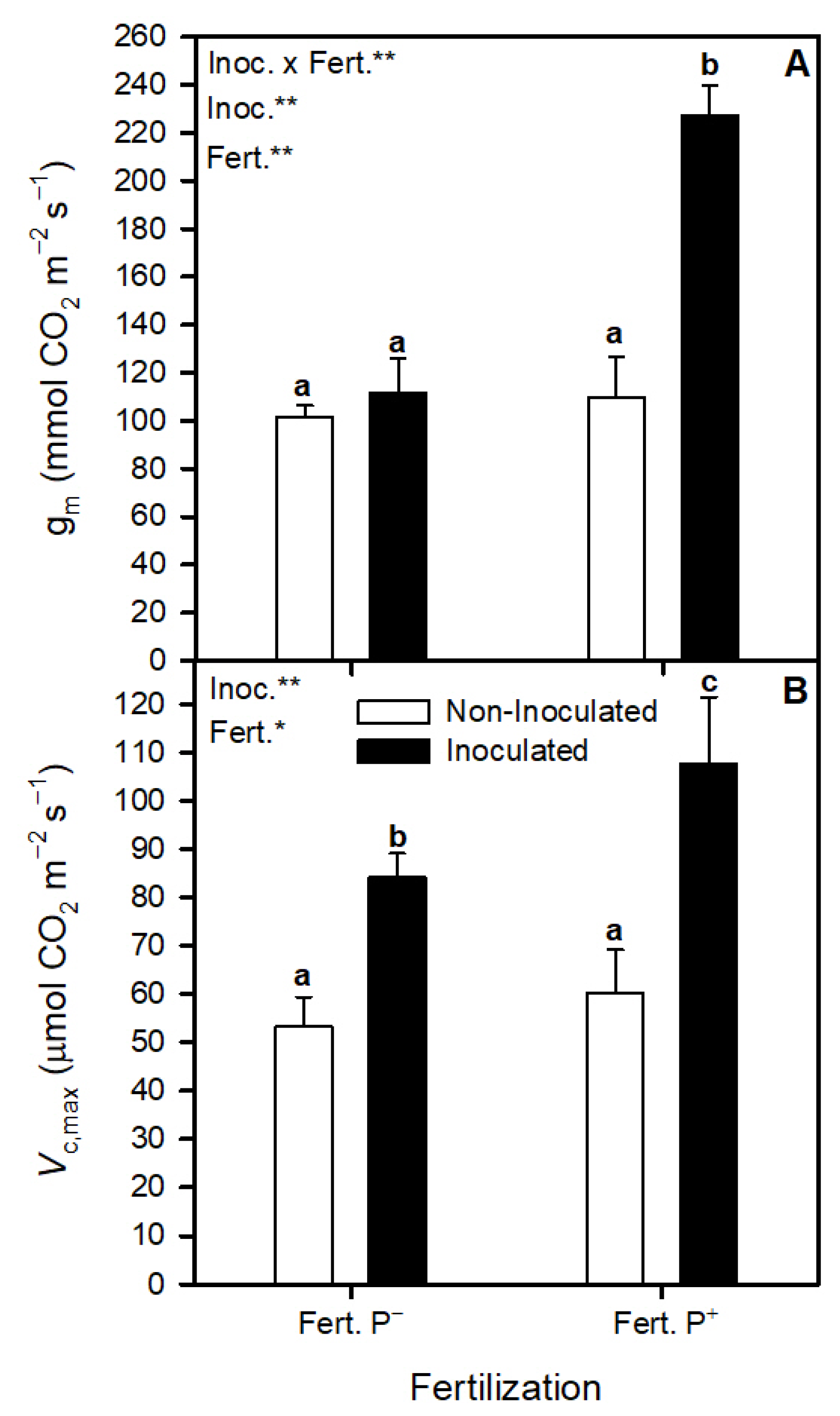
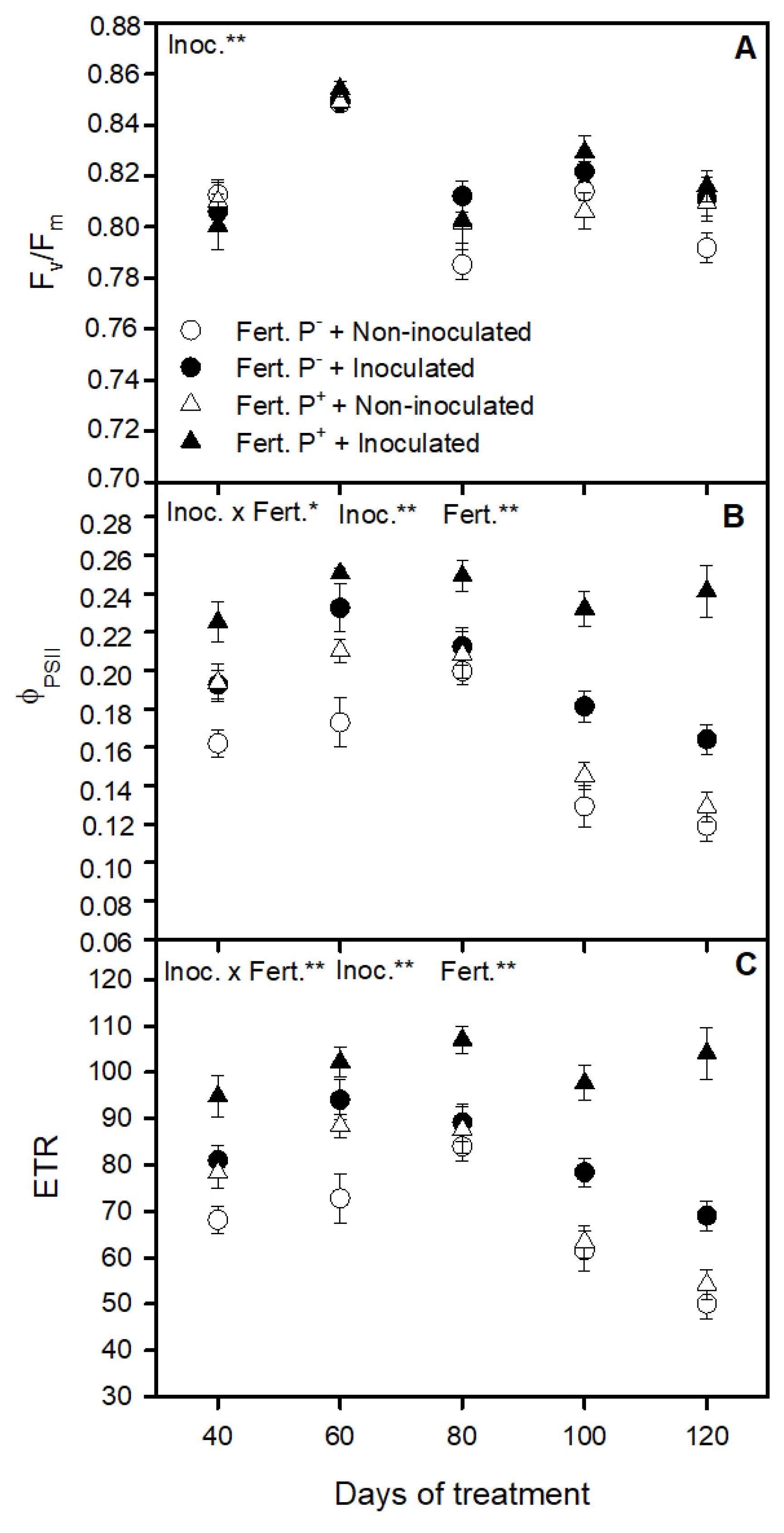
3.2. Bacterial Inoculation and Phosphorus Fertilization Availability Effect on Strawberry Photosynthetic Apparatus Performance
3.3. Bacterial Inoculation and Phosphorus Fertilization Availability Effect on Strawberry Chlorophyll Fluorescence
3.4. Bacterial Inoculation and Phosphorus Fertilization Availability Effect on Strawberry Ion and C/N Ratio
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agricultural Organization (FAO). Available online: http://www.fao.org (accessed on 1 December 2022).
- Balasooriya, H.N.; Dassanayake, K.B.; Seneweera, S.; Ajlouni, S. Interaction of Elevated Carbon Dioxide and Temperature on Strawberry (Fragaria × ananassa) Growth and Fruit Yield. Int. J. Agric. Biosyst. Eng. 2018, 12, 279–287. [Google Scholar]
- Lozano, D.; Ruiz, N.; Gavilán, P. Consumptive water use and irrigation performance of strawberries. Agri. Water Manag. 2016, 169, 44–51. [Google Scholar] [CrossRef]
- Castro, A.J.; Lopez-Rodriguez, M.D.; Giagnocavo, C.; Gimenez, M.; Cespedes, L.; La Calle, A.; Gallardo, M.; Pumares, P.; Cabello, J.; Rodriguez, E.; et al. Six collective challenges for sustainability of Almería greenhouse horticulture. Int. J. Environ. Res. Public Health 2019, 16, 4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phogat, V.; Skewes, M.; Cox, J.W.; Sanderson, G.; Alam, J.; Simunek, J. Seasonal simulation of water, salinity, chloride and nitrate dynamics under drip irrigated mandarin (Citrus reticulata) and assessing management options for drainage and nitrate leaching. J. Hydrol. 2014, 513, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Latrou, M.; Papadopoulos, A. Influence of nitrogen nutrition on nitrate levels of strawberry leaf blade and petioles. J. Plant. Nutr. 2016, 39, 1131–1136. [Google Scholar]
- Aguilera, A.; Tereucán, G.; Ercoli, S.; Cornejo, P.; Gomez, M.R.; Uhlmann, L.; Guigas, C.; Esatbeyoglu, T.; Ruiz, A. Influence of Organic and Chemical Fertilisation on Antioxidant Compounds Profiles and Activities in Fruits of Fragaria ananassa var. Camarosa. J. Soil Sci. Plant Nutr. 2020, 20, 715–724. [Google Scholar] [CrossRef]
- Yadav, S.K.; Khokhar, U.U.; Sharma, S.D.; Kumar, P. Response of strawberry to organic versus inorganic fertilizers. J. Plant. Nutr. 2016, 39, 194–203. [Google Scholar] [CrossRef]
- Pokhrel, B.; Laursen, K.H.; Petersen, K.K. Yield, quality, and nutrient concentrations of strawberry (Fragaria ×ananassa Duch. cv. ‘Sonata’) grown with different organic fertilizer strategies. J. Agric. Food. Chem. 2015, 63, 5578–5586. [Google Scholar] [CrossRef]
- Sayğı, H. Effects of organic fertilizer application on strawberry (Fragaria vesca L.) cultivation. Agronomy 2022, 12, 1233. [Google Scholar] [CrossRef]
- Reddy, G.C.; Goyal, R.K.; Godara, A.K. Effect of phosphorous in combination with biofertilizers on growth, yield and quality of strawberry cv. winter dawn. J. Environ. Biol. 2021, 42, 727–734. [Google Scholar] [CrossRef]
- Saqib, S.; Zaman, W.; Ullah, F.; Majeed, I.; Ayaz, A.; Munis, M.F.H. Organometallic assembling of chitosan-Iron oxide nanoparticles with their antifungal evaluation against Rhizopus oryzae. Appl. Organomet. Chem. 2019, 33, e5190. [Google Scholar] [CrossRef]
- Barea, J.M.; Azcon, R.; Azcon-Aguilar, C. Mycorrhizosphere interactions to improve plant fitness and soil quality. Antonie Van Leewenhoek 2002, 81, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Horticul. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Zaidi, A.; Ahmad, E.; Khan, M.S.; Saif, S.; Rizvi, A. Role of plant growth promoting rhizobacteria in sustainable production of vegetables: Current perspective. Sci. Horticul. 2015, 193, 231–239. [Google Scholar] [CrossRef]
- Gianinazzi, S.; Gollotte, A.; Binet, M.N.; van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology: The key of arbuscular mycorrhizas in ecosystem services. Micorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.; et al. Consortia of plant-growth-promoting rhizobacteria isolated from halophytes improve response of eight crops to soil salinization and climate change conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Romano-Rodríguez, E.; Mesa-Marín, J.; Sola-Elías, C.; Mateos-Naranjo, E. Consortia of plant-growth-promoting rhizobacteria isolated from halophytes improve the response of swiss chard to soil salinization. Agronomy 2022, 12, 468. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Functional aspects of plant growth promoting rhizobacteria: Recent advancements. Insight Microbiol. 2011, 1, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotech. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Ding, Z.T.; Ishap, M.; Khan, I.; Ahmed, A.A.; Khan, A.Q.; Guo, X.S. Applications of beneficial plant growth promoting rhizobacteria and mycorrhizae in rhizosphere and plant growth: A review. Int. J. Agric. Biol. Eng. 2020, 13, 199–208. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, L.; Hao, R.Q.; Bai, X.X.; Wang, Y.; Yu, X. Drought-tolerant plant growth-promoting rhizobacteria isolated from jujube (Ziziphus jujuba) and their potential to enhance drought tolerance. Plant Soil 2020, 452, 423–440. [Google Scholar] [CrossRef]
- Sunera; Amna; Saqib, S.; Uddin, S.; Zaman, W.; Ullah, F.; Ayaz, A.; Asghar, M.; Rehman, S.U.; Munis, M.F.H.; et al. Characterization and phytostimulatory activity of bacteria isolated from tomato (Lycopersicon esculentum Mill.) rhizosphere. Microb. Pathogenesis 2020, 140, 103966. [Google Scholar] [CrossRef]
- Spolaor, L.T.; Goncalves, L.S.A.; dos Santos, O.J.A.P.; de Oliveira, A.L.M.; Scapim, C.A.; Bertagna, F.A.B.; Kuki, M.C. Plant growth-promoting bacteria associated with nitrogen fertilization at topdressing in popcorn agronomic performance. Bragantia 2016, 75, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Remans, R.; Beebe, S.; Blair, M.; Manrique, G.; Tovar, E.; Rao, I.; Croonenborghs, A.; Torres-Gutierrez, R.; El-Howeity, M.; Michiels, J.; et al. Physiological and genetic analysis of root responsiveness to auxin-producing plant growth-promoting bacteria in common bean (Phaseolus vulgaris L.). Plant Soil 2008, 302, 149–161. [Google Scholar] [CrossRef]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Alijani, Z.; Amini, J.; Ashengroph, M.; Bahramnejad, B. Antifungal activity of volatile compounds produced by Staphylococcus sciuri strain MarR44 and its potential for the biocontrol of Colletotrichum nymphaeae, causal agent strawberry anthracnose. Int. J. Food Microbiol. 2019, 307, 108276. [Google Scholar] [CrossRef]
- Khruengsai, S.; Pripdeevech, P.; Tanapichatsakul, C.; Srisuwannapa, C.; D’Souza, P.E.; Panuwet, P. Antifungal properties of volatile organic compounds produced by Daldinia eschscholtzii MFLUCC 19-0493 isolated from Barleria prionitis leaves against Colletotrichum acutatum and its post-harvest infections on strawberry fruit. PeerJ 2021, 9, e11242. [Google Scholar] [CrossRef]
- Ehrhardt, D.W.; Atkinson, E.M.; Long, S.R. Depolarization of alfalfa root hair membrane potential by Rhizobium meliloti Nod factors. Science 1992, 256, 998–1000. [Google Scholar] [CrossRef] [Green Version]
- Andrades-Moreno, L.; Del Castillo, I.; Parra, R.; Doukkali, B.; Redondo-Gómez, S.; Pérez-Palacios, P.; Caviedes, M.A.; Pajuelo, E.M.; Rodríguez-Llorente, I.D. Prospecting metal-resistant plant-growth promoting rhizobacteria for rhizoremediation of metal contaminated estuaries using Spartina densiflora. Environ. Sci. Pollut. Res. 2014, 21, 3713–3721. [Google Scholar] [CrossRef]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting contaminated estuaries: Heavy metals resistant and plant growth promoting rhizobacteria in the native metal rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Impact of plant growth promoting bacteria on Salicornia ramosissima ecophysiology and heavy metal phytoremediation capacity in estuarine soils. Front. Microbiol. 2020, 17, 553018. [Google Scholar] [CrossRef] [PubMed]
- Flores-Duarte, N.J.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Pajuelo, E.; Rodriguez-Llorente, I.D.; Navarro-Torre, S. Role of nodulation-enhancing rhizobacteria in the promotion of Medicago sativa development in nutrient-poor soils. Plants 2022, 11, 1164. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of plant-growth-promoting and metal-resistant cultivable bacteria from Arthrocnemum macrostachyum in the Odiel marshes with potential use in phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef]
- Flores-Duarte, N.J.; Pérez-Pérez, J.; Navarro-Torre, S.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Pajuelo, E.; Rodriguez-Llorente, I.D. Improved Medicago sativa nodulation under stress assisted by Variovorax sp. endophytes. Plants 2022, 11, 1091. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; López-Jurado, J.; Redondo-Gómez, S.; Pérez-Romero, S.; Glick, B.R.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Echegoyan, A.; Mesa-Marín, J. Uncovering PGPB Vibrio spartinae inoculation-triggered physiological mechanisms involved in the tolerance of Halimione portulacoides to NaCl excess. Plant Physiol. Biochem. 2020, 154, 151–159. [Google Scholar] [CrossRef]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Flexas, J.; Baron, M.; Bota, J. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri x V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Niinemets, Ü.; Galle, A.; Barbour, M.M.; Centritto, M.; Diaz-Espejo, A.; Medrano, H. Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef]
- Pons, T.L.; Flexas, J.; von Caemmerer, S.; Evans, J.R.; Genty, B.; Ribas-Carbo, M.; Brugnoli, E. Estimating mesophyll conductance to CO2: Methodology, potential errors, and recommendations. J. Exp. Bot. 2009, 60, 2217–2234. [Google Scholar] [CrossRef] [Green Version]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef] [PubMed]
- de Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular-physiological functions of mineral macronutrients and their consequences for deficiency symptons in plants. New Phytol. 2020, 229, 2446–2469. [Google Scholar] [CrossRef]
- Granada, C.E.; Passaglia, L.M.P.; Souza, E.M.; Sperotto, R.A. Is phosphate solubilization the forgotten child of plant growth-promoting rhizobacteria? Front. Microbiol. 2018, 9, 2054. [Google Scholar] [CrossRef]
- Rosa, P.A.L.; Mortinho, E.S.; Jalal, A.; Galindo, F.S.; Buzetti, S.; Fernandes, G.C.; Neto, M.B.; Pavinato, P.S.; Filho, M.C.M.T. Inoculation with growth-promoting bacteria associated with the reduction of phosphate fertilization in sugarcane. Front. Environ. Sci. 2020, 8, 32. [Google Scholar] [CrossRef]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.H.; Hartmann, A.; Traini, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat. Commun. 2018, 9, 1409. [Google Scholar] [CrossRef] [Green Version]
- De Silva, A.; Patterson, K.; Rothrock, C.; Moore, J. Growth promotion of highbush blueberry by fungal and bacterial inoculants. Hortscience 2000, 35, 1228–1230. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Bernabeu-Meana, M.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S. Effect of plant growth-promoting rhizobacteria on Salicornia ramosissima seed germination under salinity, CO2 and temperature stress. Agronomy 2019, 9, 655. [Google Scholar] [CrossRef] [Green Version]
- Esitken, A.; Yildiz, H.E.; Ercisli, S.; Donmez, M.F.; Turan, M.; Gunes, A. Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Sci. Horticul. 2010, 124, 62–66. [Google Scholar] [CrossRef]
- Hinsinger, P.; Brauman, A.; Devau, N.; Gerard, F.; Jourdan, C.; Laclau, J.P.; Le Cadre, E.; Jaillard, B.; Plassard, C. Acquisition of phosphorus and other poorly mobile nutrients by roots. Where do plant nutrition models fail? Plant Soil 2011, 348, 29–61. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kamerlin, S.C.L.; Sharma, P.K.; Prasad, R.B.; Warshel, A. Why nature really choose phosphates. Q. Rev. Biophys. 2013, 46, 1–132. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.X.; Weng, X.Y.; Yang, Y. Effect of phosphorous deficiency on the photosynthesis characteristics of rice plants. R. J. Plant Physiol. 2007, 54, 741–748. [Google Scholar] [CrossRef]
- Morales, F.; Warren, C.R. Photosynthesis Responses to Nutrient Deprivation and Toxicities. In Terrestrial Photosynthesis in a Changing Environment; Flexas, J., Loreto, F., Medrano, I., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 312–330. [Google Scholar]
- Bown, H.E.; Watt, M.S.; Clinton, P.W. The influence of N and P supply and genotype on carbon flux and partitioning in potted Pinus radiata plants. Tree Physiol. 2009, 29, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Hammond, J.P.; White, P.J. Sucrose transport in the phloem: Integrating root responses to phosphorus starvation. J. Exp. Bot. 2008, 59, 93–109. [Google Scholar] [CrossRef]
- Warren, C.R. How does P affect photosynthesis and metabolite profiles of Eucalyptus globulus? Tree Physiol. 2011, 31, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. Mapping the impacts of phosphorus deficiency on the photosynthetic electron transport chain in barley. Plant Physiol. 2018, 177, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef]
- de Souza, R.; Meyer, J.; Schoenfeld, R.; da Costa, P.B.; Passaglia, L.M.P. Characterization of plant growth-promoting bacteria associated with rice cropped in iron-stressed soils. Ann. Microbiol. 2015, 65, 951–964. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.H.; Chen, L.S.; Chen, R.B.; Zhang, F.Z.; Jiang, H.X.; Tang, N. CO2 assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport probed by the JIP-test, of tea leaves in response to phosphorus supply. BMC Plant Biol. 2009, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGP | SDT3 | HPJ40 | SMT38 | SRT15 | S110 |
|---|---|---|---|---|---|
| Phosphate solubilizing | + | + | - | + | - |
| Siderophores | + | + | + | - | + |
| IAA | - | - | - | + | + |
| Fixation of nitrogen | - | + | + | + | + |
| Biofilm | - | + | + | + | + |
| ACC activity | n.d | - | + | - | + |
| Treatments | Leaves Concentrations | ||||||
|---|---|---|---|---|---|---|---|
| Inoculation | Fertilization | Ca (mg g−1) | K (mg g−1) | Mg (mg g−1) | P (mg g−1)Inoc. ** | Mn (mg Kg−1) | C/NInoc., Fert. ** |
| − | P− | 9.4 ± 0.2 | 21.5 ± 0.8 | 5.9 ± 0.2 | 1.2 ± 0.0 | 703.5 ± 12.6 | 27.5 ± 0.1 |
| − | P+ | 9.0 ± 0.1 | 19.6 ± 0.1 | 5.6 ± 0.0 | 1.3 ± 0.0 | 705.1 ± 7.2 | 25.4 ± 0.5 |
| + | P− | 8.9 ± 0.1 | 21.1 ± 0.4 | 5.9 ± 0.1 | 1.6 ± 0.0 | 700.0 ± 10.6 | 24.1 ± 0.2 |
| + | P+ | 8.7 ± 0.1xd | 19.3 ± 0.2 | 5.8 ± 0.1 | 1.7 ± 0.0 | 727.0 ± 6.5 | 22.1 ± 0.1 |
| Roots Concentrations | |||||||
| Inoculation | Fertilization | Ca (mg g−1)Inoc. ** | K (mg g−1)Inoc., Fert. ** | Mg (mg g−1) | P (mg g−1)Inoc., Fert. ** | Mn (mg Kg−1) | C/NInoc. x Fert. * |
| − | P− | 4.2 ± 0.1 | 8.9 ± 0.2 | 6.5 ± 0.8 | 1.0 ± 0.1 | 160.0 ± 1.6 | 60.2 ± 0.8 |
| − | P+ | 4.7 ± 0.2 | 10.5 ± 0.9 | 5.3 ± 0.5 | 1.2 ± 0.1 | 160.5 ± 7.1 | 54.3 ± 0.6 |
| + | P− | 5.2 ± 0.2 | 12.9 ± 0.3 | 6.4 ± 0.3 | 1.4 ± 0.0 | 163.3 ± 8.5 | 51.2 ± 1.3 |
| + | P+ | 5.2 ± 0.2 | 14.5 ± 0.7 | 6.4 ± 0.1 | 1.6 ± 0.0 | 180.4 ± 1.0 | 50.8 ± 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valle-Romero, P.; García-López, J.V.; Redondo-Gómez, S.; Flores-Duarte, N.J.; Rodríguez-Llorente, I.D.; Idaszkin, Y.L.; Pajuelo, E.; Mateos-Naranjo, E. Biofertilization with PGP Bacteria Improve Strawberry Plant Performance under Sub-Optimum Phosphorus Fertilization. Agronomy 2023, 13, 335. https://doi.org/10.3390/agronomy13020335
Valle-Romero P, García-López JV, Redondo-Gómez S, Flores-Duarte NJ, Rodríguez-Llorente ID, Idaszkin YL, Pajuelo E, Mateos-Naranjo E. Biofertilization with PGP Bacteria Improve Strawberry Plant Performance under Sub-Optimum Phosphorus Fertilization. Agronomy. 2023; 13(2):335. https://doi.org/10.3390/agronomy13020335
Chicago/Turabian StyleValle-Romero, Pedro, Jesús V. García-López, Susana Redondo-Gómez, Noris J. Flores-Duarte, Ignacio D. Rodríguez-Llorente, Yanina Lorena Idaszkin, Eloisa Pajuelo, and Enrique Mateos-Naranjo. 2023. "Biofertilization with PGP Bacteria Improve Strawberry Plant Performance under Sub-Optimum Phosphorus Fertilization" Agronomy 13, no. 2: 335. https://doi.org/10.3390/agronomy13020335