
New Insights from Soil Microorganisms for Sustainable Double Rice-Cropping System with 37-Year Manure Fertilization
 , , and
, , and
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Site Description
2.2. Experiment Design
2.3. Crop Management
2.4. Sample Collection
2.5. Measurement of Soil Properties
2.6. Measurement of Soil Bacterial Community
2.7. Statistical Analysis
3. Results
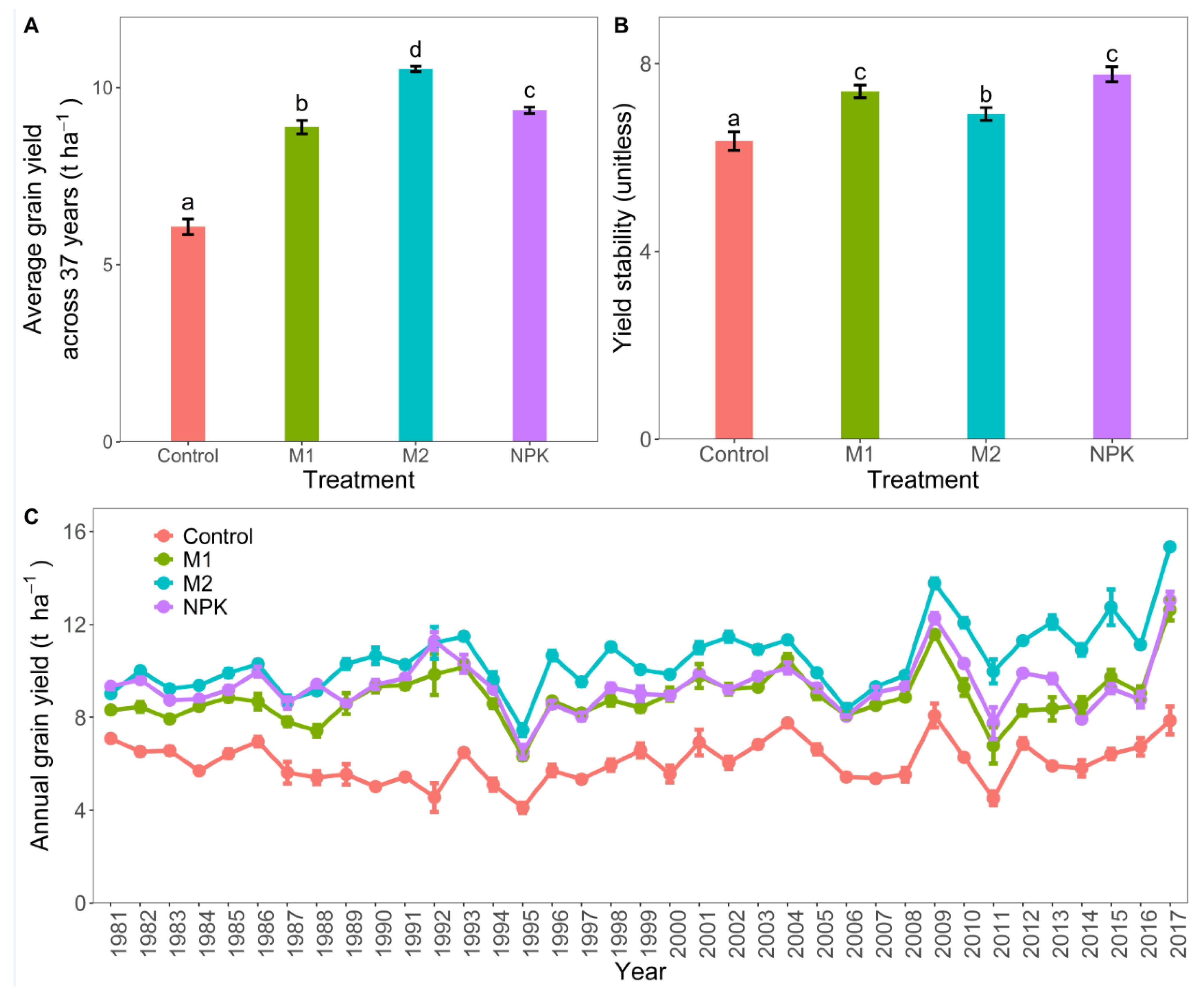
3.1. Rice Grain Yield and Yield Stability
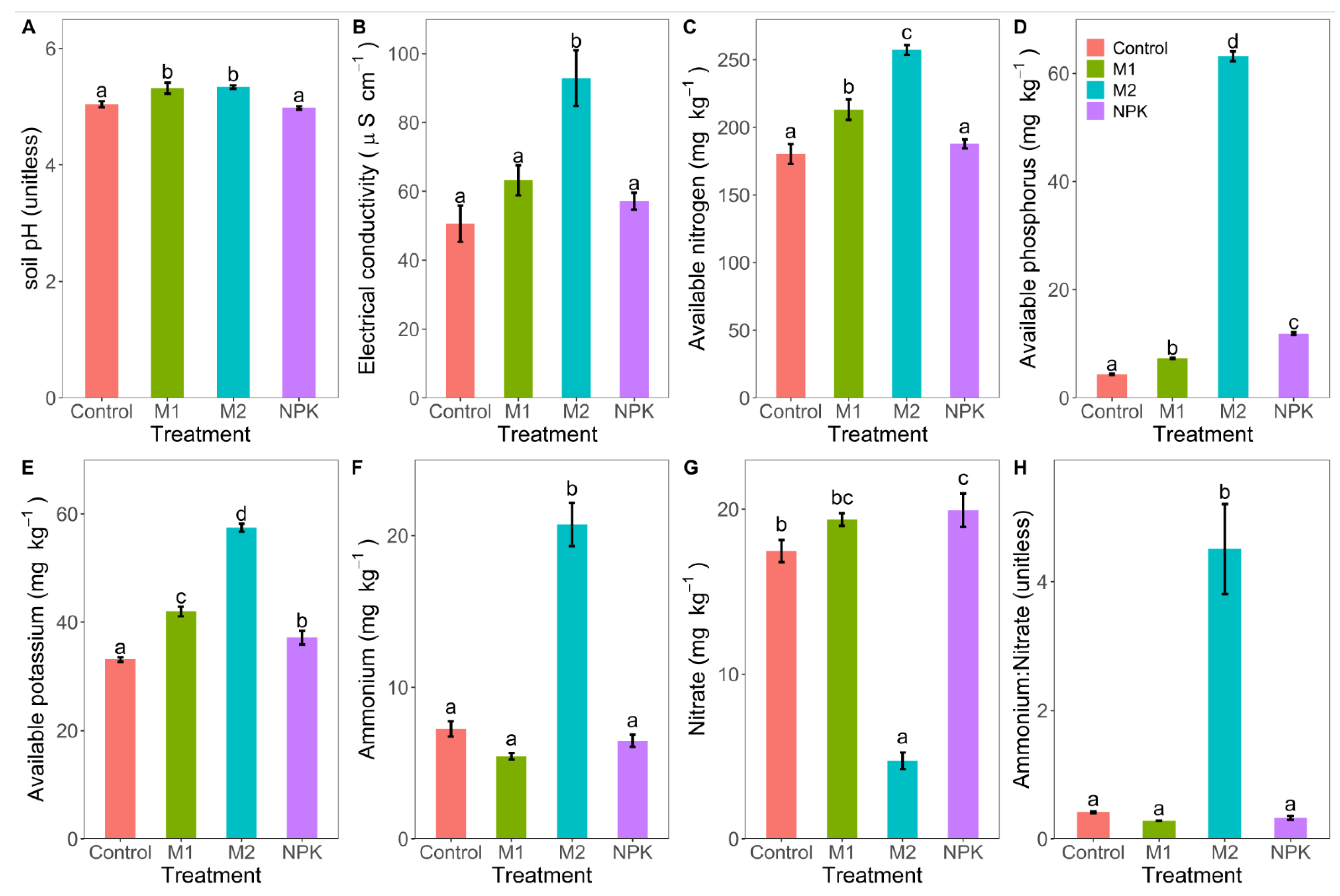
3.2. Soil pH, EC and Nutrient Availability
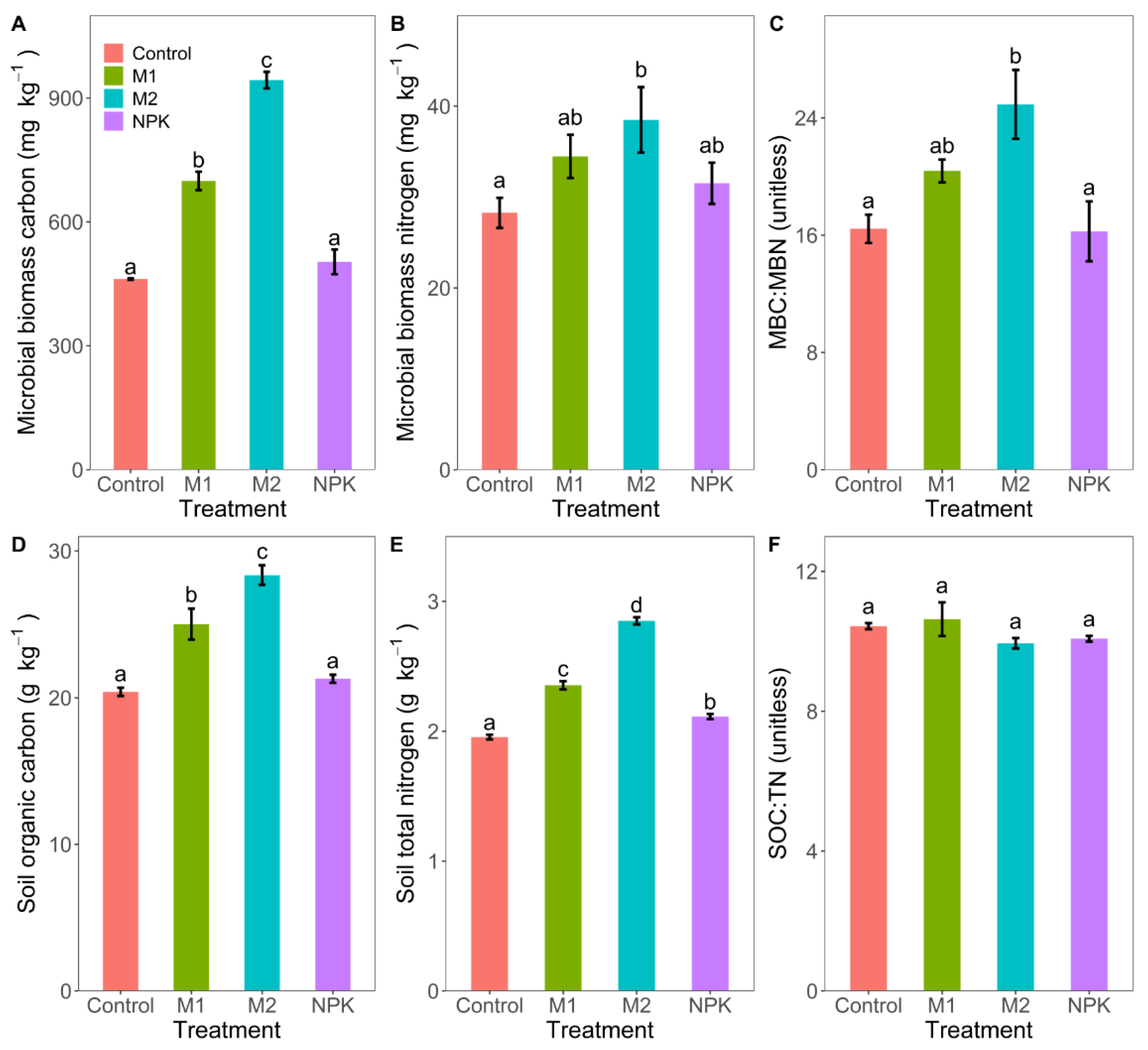
3.3. Microbial Biomass Carbon and Nitrogen, Soil Organic Carbon and Total Nitrogen
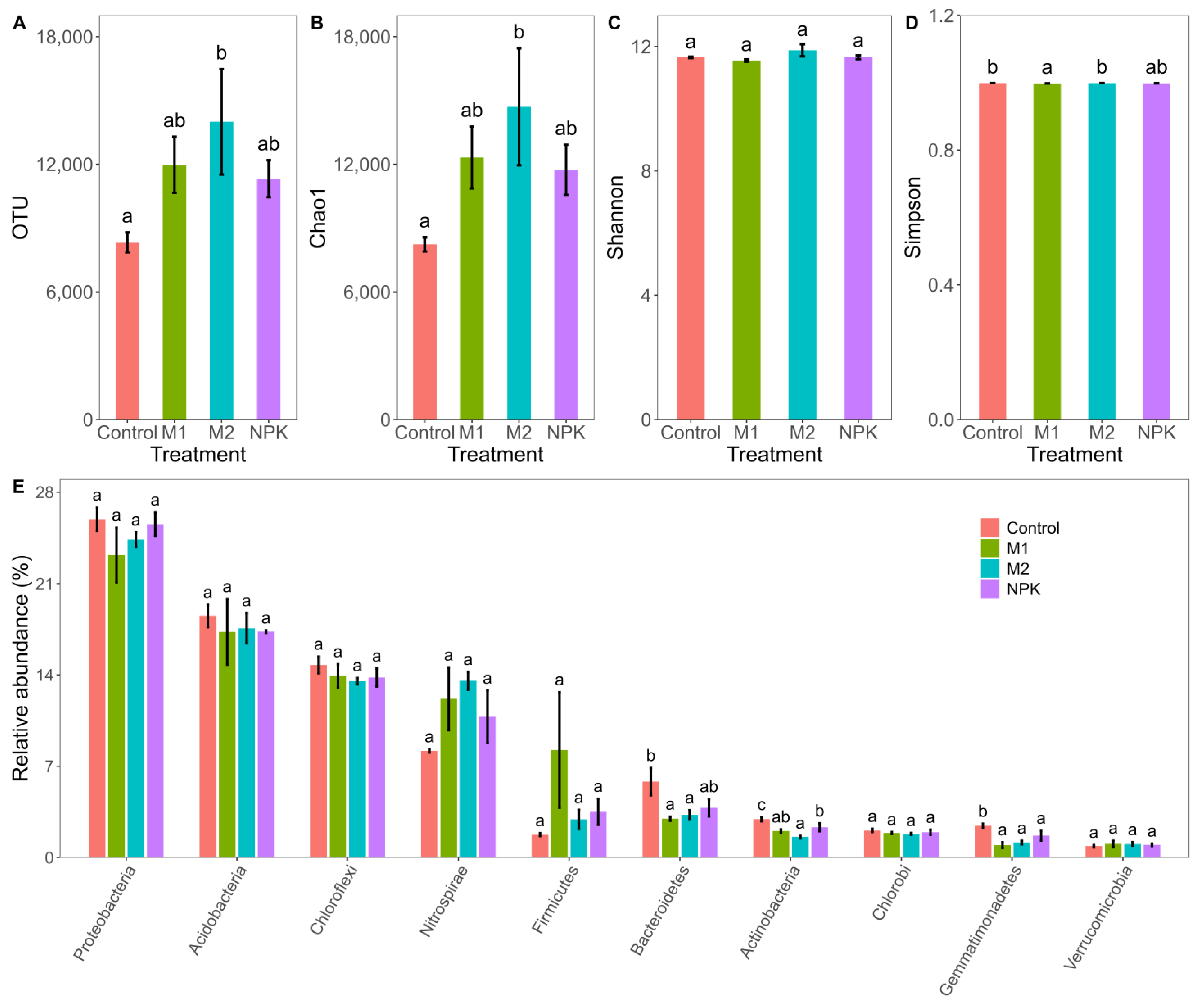
3.4. Bacterial Community Diversity and Composition
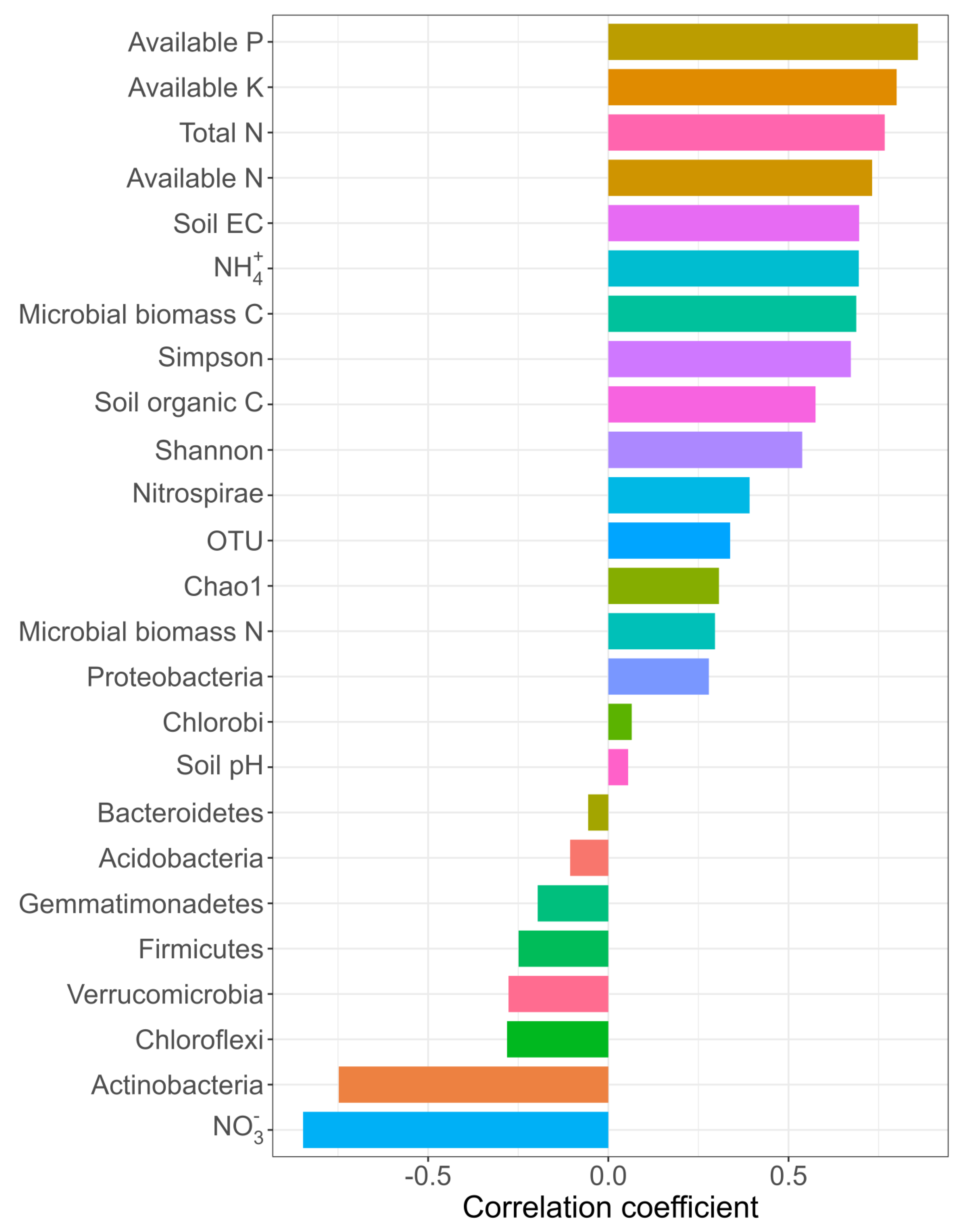
3.5. Factors Affecting Grain Yield, Yield Stability, and Bacterial Community
4. Discussion
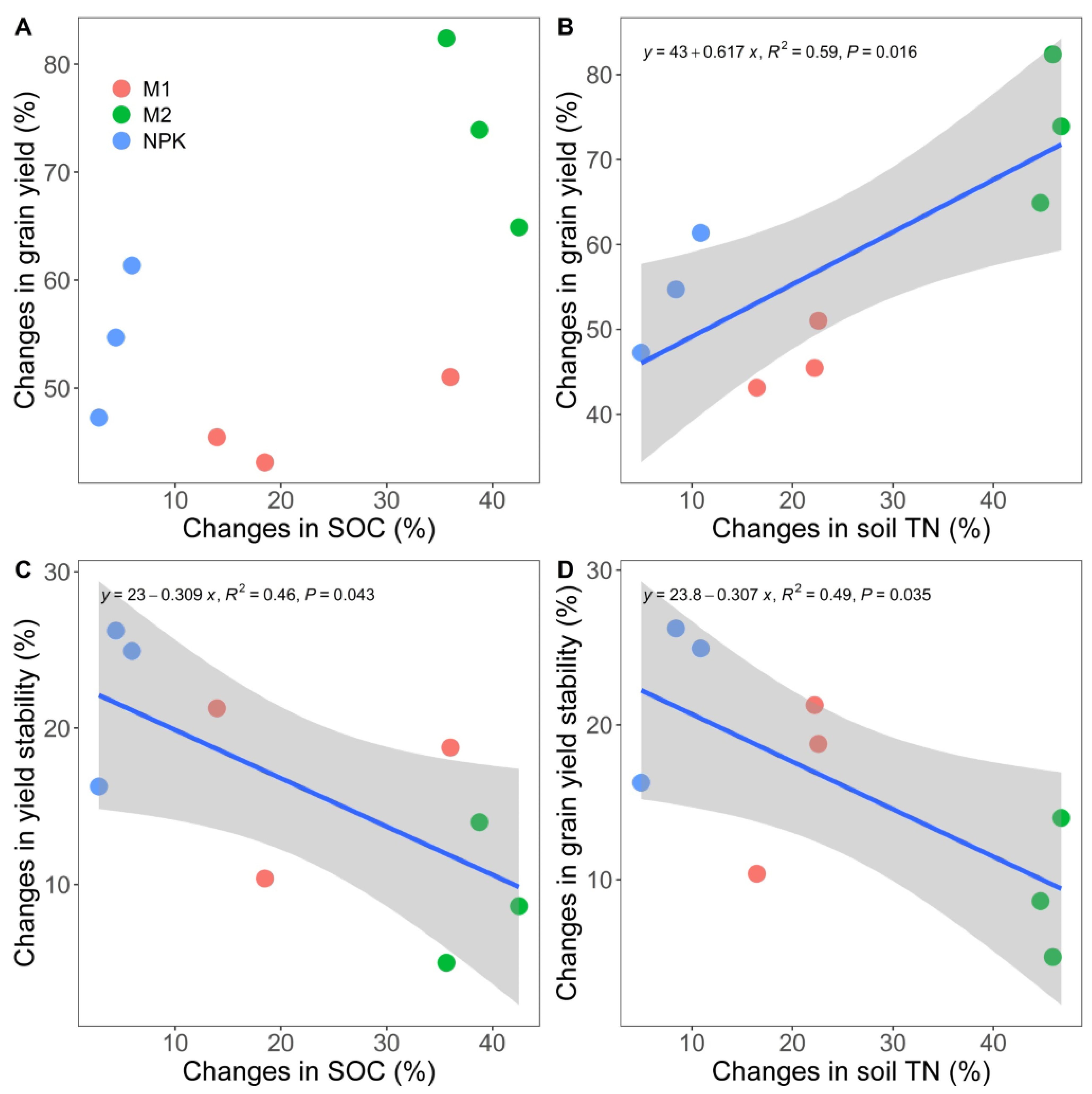
4.1. Increased Rice Grain Yield and Yield Stability with Long-Term Fertilization
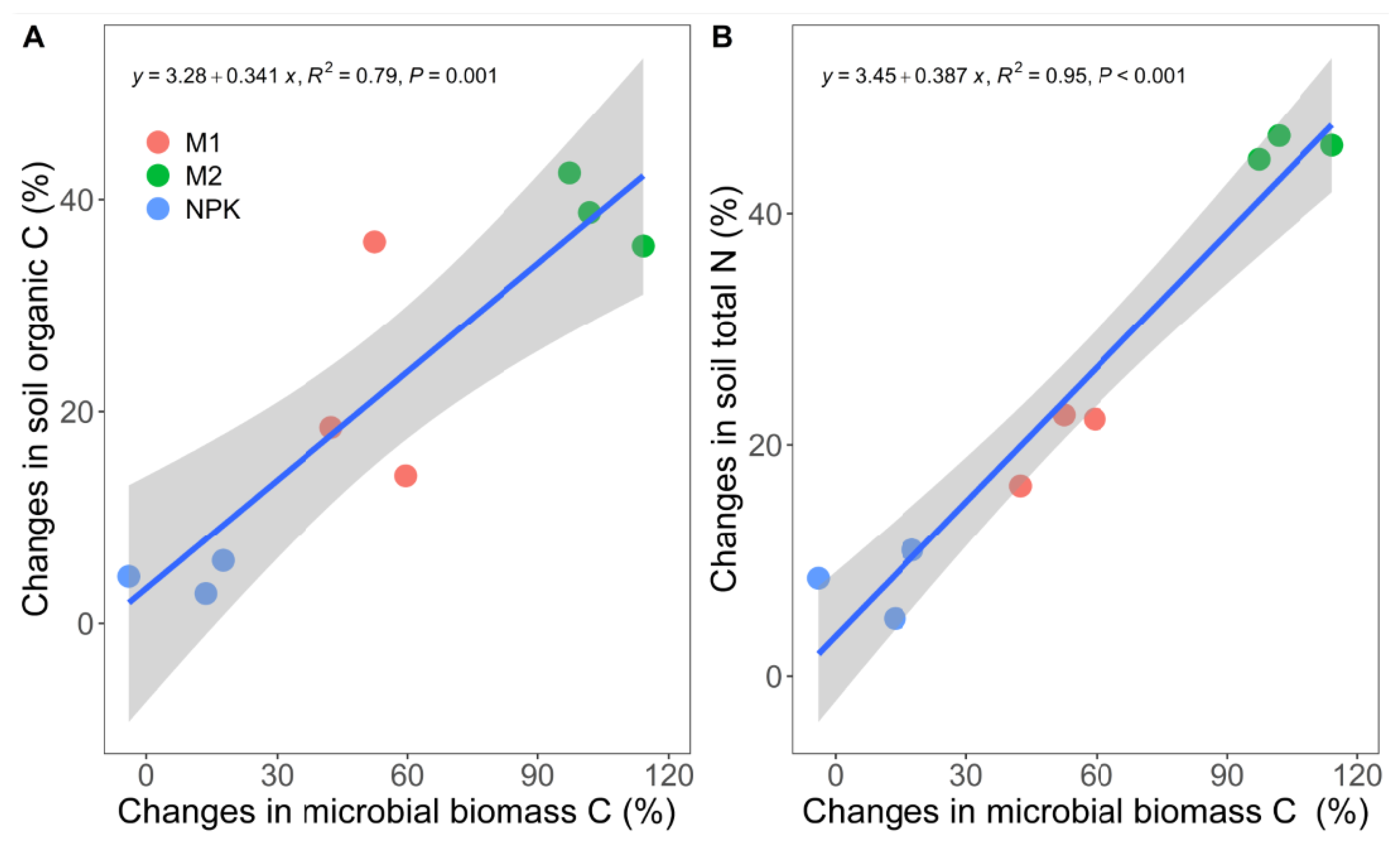
4.2. Enhanced Soil Organic Carbon and Total Nitrogen Content with Long-Term Fertilization
4.3. Shifts in Bacterial Community
4.4. Changes in Soil Physicochemical Variables
4.5. Uncertainties and Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van Nguyen, N.; Ferrero, A. Meeting the challenges of global rice production. Paddy Water Environ. 2006, 4, 1–9. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, P.; Patel, S.C. Pattern recognition method to detect two diseases in rice plants. Imaging Sci. J. 2008, 56, 319–325. [Google Scholar] [CrossRef]
- Tariq, A.; Vu, Q.D.; Jensen, L.S.; de Tourdonnet, S.; Sander, B.O.; Wassmann, R.; Van Mai, T.; de Neergaard, A. Mitigating CH 4 and N 2 O emissions from intensive rice production systems in northern Vietnam: Efficiency of drainage patterns in combination with rice residue incorporation. Agric. Ecosyst. Environ. 2017, 249, 101–111. [Google Scholar] [CrossRef]
- Shang, Q.; Yang, X.; Gao, C.; Wu, P.; Liu, J.; Xu, Y.; Shen, Q.; Zou, J.; Guo, S. Net annual global warming potential and greenhouse gas intensity in Chinese double rice-cropping systems: A 3-year field measurement in long-term fertilizer experiments. Glob. Chang. Biol. 2010, 17, 2196–2210. [Google Scholar] [CrossRef]
- Kirk, G.; George, T.; Courtois, B.; Senadhira, D. Opportunities to improve phosphorus efficiency and soil fertility in rainfed lowland and upland rice ecosystems. Field Crop. Res. 1998, 56, 73–92. [Google Scholar] [CrossRef]
- Dong, W.; Zhang, X.; Wang, H.; Dai, X.; Sun, X.; Qiu, W.; Yang, F. Effect of Different Fertilizer Application on the Soil Fertility of Paddy Soils in Red Soil Region of Southern China. PLoS ONE 2012, 7, e44504. [Google Scholar] [CrossRef] [Green Version]
- Dawe, D.; Dobermann, A.; Ladha, J.; Yadav, R.; Bao, L.; Gupta, R.; Lal, P.; Panaullah, G.; Sariam, O.; Singh, Y.; et al. Do organic amendments improve yield trends and profitability in intensive rice systems? Field Crop. Res. 2003, 83, 191–213. [Google Scholar] [CrossRef]
- Melandri, G.; AbdElgawad, H.; Riewe, D.; Hageman, J.A.; Asard, H.; Beemster, G.T.S.; Kadam, N.; Jagadish, K.; Altmann, T.; Ruyter-Spira, C.; et al. Biomarkers for grain yield stability in rice under drought stress. J. Exp. Bot. 2020, 71, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Yuan, S.; Man, J. Changes in rice yield and yield stability in China during the past six decades. J. Sci. Food Agric. 2020, 100, 3560–3569. [Google Scholar] [CrossRef] [PubMed]
- Tollenaar, M.; Lee, E. Yield potential, yield stability and stress tolerance in maize. Field Crop. Res. 2002, 75, 161–169. [Google Scholar] [CrossRef]
- Raseduzzaman; Jensen, E.S. Does intercropping enhance yield stability in arable crop production? A meta-analysis. Eur. J. Agron. 2017, 91, 25–33. [Google Scholar] [CrossRef]
- Thierfelder, C.; Chivenge, P.; Mupangwa, W.; Rosenstock, T.S.; Lamanna, C.; Eyre, J.X. How climate-smart is conservation agriculture (CA)?—Its potential to deliver on adaptation, mitigation and productivity on smallholder farms in southern Africa. Food Secur. 2017, 9, 537–560. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.C.; Leon, J. Stability Analysis in Plant Breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Amelung, W.; Bossio, D.; de Vries, W.; Kögel-Knabner, I.; Lehmann, J.; Amundson, R.; Bol, R.; Collins, C.; Lal, R.; Leifeld, J.; et al. Towards a global-scale soil climate mitigation strategy. Nat. Commun. 2020, 11, 5427. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Huang, S.; Yu, X.; Zhang, W. Stability and saturation of soil organic carbon in rice fields: Evidence from a long-term fertilization experiment in subtropical China. J. Soils Sediments 2013, 13, 1327–1334. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Roy, K.; Neogi, S.; Adhya, T.; Rao, K.; Manna, M. Effects of rice straw and nitrogen fertilization on greenhouse gas emissions and carbon storage in tropical flooded soil planted with rice. Soil Tillage Res. 2012, 124, 119–130. [Google Scholar] [CrossRef]
- Jiang, Y.; Carrijo, D.; Huang, S.; Chen, J.; Balaine, N.; Zhang, W.; van Groenigen, K.J.; Linquist, B. Water management to mitigate the global warming potential of rice systems: A global meta-analysis. Field Crop. Res. 2019, 234, 47–54. [Google Scholar] [CrossRef]
- Gerber, J.S.; Carlson, K.M.; Makowski, D.; Mueller, N.D.; de Cortazar-Atauri, I.G.; Havlík, P.; Herrero, M.; Launay, M.; O’Connell, C.S.; Smith, P.; et al. Spatially explicit estimates of N2O emissions from croplands suggest climate mitigation opportunities from improved fertilizer management. Glob. Chang. Biol. 2016, 22, 3383–3394. [Google Scholar] [CrossRef]
- Nayak, A.; Gangwar, B.; Shukla, A.K.; Mazumdar, S.P.; Kumar, A.; Raja, R.; Kumar, A.; Kumar, V.; Rai, P.; Mohan, U. Long-term effect of different integrated nutrient management on soil organic carbon and its fractions and sustainability of rice–wheat system in Indo Gangetic Plains of India. Field Crop. Res. 2012, 127, 129–139. [Google Scholar] [CrossRef]
- Zhu, Y.; Waqas, M.A.; Li, Y.; Zou, X.; Jiang, D.; Wilkes, A.; Qin, X.; Gao, Q.; Wan, Y.; Hasbagan, G. Large-scale farming operations are win-win for grain production, soil carbon storage and mitigation of greenhouse gases. J. Clean. Prod. 2018, 172, 2143–2152. [Google Scholar] [CrossRef]
- Chen, J.; Lærke, P.E.; Jørgensen, U. Optimized crop rotations increase biomass production without significantly changing soil carbon and nitrogen stock. Ecol. Indic. 2020, 117, 106669. [Google Scholar] [CrossRef]
- Iovieno, P.; Morra, L.; Leone, A.; Pagano, L.; Alfani, A. Effect of organic and mineral fertilizers on soil respiration and enzyme activities of two Mediterranean horticultural soils. Biol. Fertil. Soils 2009, 45, 555–561. [Google Scholar] [CrossRef]
- Kimura, M.; Murase, J.; Lu, Y. Carbon cycling in rice field ecosystems in the context of input, decomposition and translocation of organic materials and the fates of their end products (CO2 and CH4). Soil Biol. Biochem. 2004, 36, 1399–1416. [Google Scholar] [CrossRef]
- Yadvinder, S.; Bijay, S.; Ladha, J.K.; Khind, C.S.; Khera, T.S.; Bueno, C.S. Effects of residue decomposition on productivity and soil fertility in rice–wheat rotation. Soil Sci. Soc. Am. J. 2004, 68, 854–864. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Pirdashti, H.; Gilani, Z. Application of mathematical models to describe rice growth and nutrients uptake in the presence of plant growth promoting microorganisms. Appl. Soil Ecol. 2018, 124, 171–184. [Google Scholar] [CrossRef]
- Mäder, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A.; et al. Inoculation of root microorganisms for sustainable wheat–rice and wheat–black gram rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Johnston, A.E.; Poulton, P.R. The importance of long-term experiments in agriculture: Their management to ensure continued crop production and soil fertility; the Rothamsted experience. Eur. J. Soil Sci. 2018, 69, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Suter, H.; He, J.-Z.; Hu, H.-W.; Chen, D. Dissimilatory nitrate reduction to ammonium dominates nitrate reduction in long-term low nitrogen fertilized rice paddies. Soil Biol. Biochem. 2019, 131, 149–156. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E.; Andersen, A.; Bowman, D.; Bull, C.M.; Burns, E.; Dickman, C.R.; Hoffmann, A.A.; Keith, D.A.; Liddell, M.J.; et al. Value of long-term ecological studies. Austral Ecol. 2012, 37, 745–757. [Google Scholar] [CrossRef]
- Bi, L.; Zhang, B.; Liu, G.; Li, Z.; Liu, Y.; Ye, C.; Yu, X.; Lai, T.; Zhang, J.; Yin, J.; et al. Long-term effects of organic amendments on the rice yields for double rice cropping systems in subtropical China. Agric. Ecosyst. Environ. 2009, 129, 534–541. [Google Scholar] [CrossRef]
- Hughes, B.B.; Beas-Luna, R.; Barner, A.K.; Brewitt, K.; Brumbaugh, D.R.; Cerny-Chipman, E.B.; Close, S.L.; Coblentz, K.E.; De Nesnera, K.L.; Drobnitch, S.T.; et al. Long-term studies contribute disproportionately to ecology and policy. BioScience 2017, 67, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.G.; Menalled, F.D.; Robertson, G.P. Temporal Yield Variability under Conventional and Alternative Management Systems. Agron. J. 2007, 99, 1629–1634. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, H.; Dong, Q.; Xia, L.; Li, J.; Li, C.; Zang, H.; Andersen, M.N.; Olesen, J.E.; Jørgensen, U.; et al. Ammoniated straw incorporation increases wheat yield, yield stability, soil organic carbon and soil total nitrogen content. Field Crop. Res. 2022, 284, 108558. [Google Scholar] [CrossRef]
- Wang, H.; Shen, M.; Hui, D.; Chen, J.; Sun, G.; Wang, X.; Lu, C.; Sheng, J.; Chen, L.; Luo, Y.; et al. Straw incorporation influences soil organic carbon sequestration, greenhouse gas emission, and crop yields in a Chinese rice (Oryza sativa L.) –wheat (Triticum aestivum L.) cropping system. Soil Tillage Res. 2019, 195, 104377. [Google Scholar] [CrossRef]
- Pedraza, R.; Bellone, C.H.; de Bellone, S.C.; Sorte, P.M.F.B.; Teixeira, K.R.D.S. Azospirillum inoculation and nitrogen fertilization effect on grain yield and on the diversity of endophytic bacteria in the phyllosphere of rice rainfed crop. Eur. J. Soil Biol. 2009, 45, 36–43. [Google Scholar] [CrossRef]
- Buresh, R.J.; Pampolino, M.F.; Witt, C. Field-specific potassium and phosphorus balances and fertilizer requirements for irrigated rice-based cropping systems. Plant Soil 2010, 335, 35–64. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Sheng, H.; Li, Y.; Tong, C.; Ge, T.; Wu, J. Lower C sequestration and N use efficiency by straw incorporation than manure amendment on paddy soils. Agric. Ecosyst. Environ. 2016, 219, 93–100. [Google Scholar] [CrossRef]
- Briones, A.M.; Okabe, S.; Umemiya, Y.; Ramsing, N.-B.; Reichardt, W.; Okuyama, H. Ammonia-oxidizing bacteria on root biofilms and their possible contribution to N use efficiency of different rice cultivars. Plant Soil 2003, 250, 335–348. [Google Scholar] [CrossRef]
- Wong, J.; Ma, K.; Fang, K.; Cheung, C. Utilization of a manure compost for organic farming in Hong Kong. Bioresour. Technol. 1999, 67, 43–46. [Google Scholar] [CrossRef]
- Zhai, Z.; Luo, M.; Yang, Y.; Liu, Y.; Chen, X.; Zhang, C.; Huang, J.; Chen, J. Trade-off between microbial carbon use efficiency and microbial phosphorus limitation under salinization in a tidal wetland. Catena 2021, 209, 105809. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Hassan, S.; Tanveer, M.; Ihsan, M.Z.; Shah, A.N.; Ullah, A.; Khan, F.; Ullah, S.; et al. A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice. Plant Physiol. Biochem. 2016, 103, 191–198. [Google Scholar] [CrossRef]
- Li, Y.; Feng, H.; Chen, J.; Lu, J.; Wu, W.; Liu, X.; Li, C.; Dong, Q.; Siddique, K.H. Biochar incorporation increases winter wheat (Triticum aestivum L.) production with significantly improving soil enzyme activities at jointing stage. Catena 2022, 211, 105979. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, X.; Liu, Z.; Pullens, J.W.; Chen, J.; Marek, G.W.; Chen, Y.; Lv, S.; Sun, S. Greater maize yield improvements in low/unstable yield zones through recommended nutrient and water inputs in the main cropping regions, China. Agric. Water Manag. 2020, 232, 106018. [Google Scholar] [CrossRef]
- Chen, J.; van Groenigen, K.J.; Hungate, B.A.; Terrer, C.; van Groenigen, J.-W.; Maestre, F.T.; Ying, S.C.; Luo, Y.; Jørgensen, U.; Sinsabaugh, R.L.; et al. Long-term nitrogen loading alleviates phosphorus limitation in terrestrial ecosystems. Glob. Chang. Biol. 2020, 26, 5077–5086. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Luo, Y.; van Groenigen, K.J.; Hungate, B.A.; Cao, J.; Zhou, X.; Wang, R.-W. A keystone microbial enzyme for nitrogen control of soil carbon storage. Sci. Adv. 2018, 4, eaaq1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pampolino, M.F.; Laureles, E.V.; Gines, H.C.; Buresh, R.J. Soil Carbon and Nitrogen Changes in Long-Term Continuous Lowland Rice Cropping. Soil Sci. Soc. Am. J. 2008, 72, 798–807. [Google Scholar] [CrossRef]
- Nayak, D.R.; Babu, Y.J.; Adhya, T. Long-term application of compost influences microbial biomass and enzyme activities in a tropical Aeric Endoaquept planted to rice under flooded condition. Soil Biol. Biochem. 2007, 39, 1897–1906. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, C.H.; Jung, K.Y.; Park, K.D.; Lee, D.; Kim, P.J. Changes of soil organic carbon and its fractions in relation to soil physical properties in a long-term fertilized paddy. Soil Tillage Res. 2009, 104, 227–232. [Google Scholar] [CrossRef]
- Rasool, R.; Kukal, S.; Hira, G. Soil physical fertility and crop performance as affected by long term application of FYM and inorganic fertilizers in rice–wheat system. Soil Tillage Res. 2007, 96, 64–72. [Google Scholar] [CrossRef]
- Mitran, T.; Mani, P.K.; Bandyopadhyay, P.K.; Basak, N. Effects of Organic Amendments on Soil Physical Attributes and Aggregate-Associated Phosphorus Under Long-Term Rice-Wheat Cropping. Pedosphere 2018, 28, 823–832. [Google Scholar] [CrossRef]
- Li, J.; Ye, X.; Zhang, Y.; Chen, J.; Yu, N.; Zou, H. Maize Straw Deep-Burying Promotes Soil Bacteria Community Abundance and Improves Soil Fertility. J. Soil Sci. Plant Nutr. 2021, 21, 1397–1407. [Google Scholar] [CrossRef]
- Kumar, U.; Shahid, M.; Tripathi, R.; Mohanty, S.; Kumar, A.; Bhattacharyya, P.; Lal, B.; Gautam, P.; Raja, R.; Panda, B.B.; et al. Variation of functional diversity of soil microbial community in sub-humid tropical rice-rice cropping system under long-term organic and inorganic fertilization. Ecol. Indic. 2017, 73, 536–543. [Google Scholar] [CrossRef]
- Zealand, A.M.; Mei, R.; Papachristodoulou, P.; Roskilly, A.P.; Liu, W.T.; Graham, D.W. Microbial community composition and diversity in rice straw digestion bioreactors with and without dairy manure. Appl. Microbiol. Biotechnol. 2018, 102, 8599–8612. [Google Scholar] [CrossRef]
- Zhang, Q.-C.; Shamsi, I.H.; Xu, D.-T.; Wang, G.-H.; Lin, X.-Y.; Jilani, G.; Hussain, N.; Chaudhry, A.N. Chemical fertilizer and organic manure inputs in soil exhibit a vice versa pattern of microbial community structure. Appl. Soil Ecol. 2012, 57, 1–8. [Google Scholar] [CrossRef]
- Nguyen, B.T.; Trinh, N.N.; Bach, Q.-V. Methane emissions and associated microbial activities from paddy salt-affected soil as influenced by biochar and cow manure addition. Appl. Soil Ecol. 2020, 152, 103531. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhu, L. Biochar alters microbial community and carbon sequestration potential across different soil pH. Sci. Total. Environ. 2018, 622–623, 1391–1399. [Google Scholar] [CrossRef]
- Pascual, J.; García-López, M.; Bills, G.; Genilloud, O. Longimicrobium terrae gen. nov., sp. nov., an oligotrophic bacterium of the under-represented phylum Gemmatimonadetes isolated through a system of miniaturized diffusion chambers. Int. J. Syst. Evol. Microbiol. 2016, 66, 1976–1985. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yan, X.; Wang, D.; Siddique, I.A.; Chen, J.; Xu, Q.; Zhao, C.; Yang, L.; Miao, Y.; Han, S. Soil Microbial Community Based on PLFA Profiles in an Age Sequence of Pomegranate Plantation in the Middle Yellow River Floodplain. Diversity 2021, 13, 408. [Google Scholar] [CrossRef]
- Ren, C.; Chen, J.; Deng, J.; Zhao, F.; Han, X.; Yang, G.; Tong, X.; Feng, Y.; Shelton, S.; Ren, G. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fertil. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Wu, J.; Liu, X.; Xu, J. Impact of organic matter addition on pH change of paddy soils. J. Soils Sediments 2012, 13, 12–23. [Google Scholar] [CrossRef]
- Zhou, W.; Guo, Z.; Chen, J.; Jiang, J.; Hui, D.; Wang, X.; Sheng, J.; Chen, L.; Luo, Y.; Zheng, J.; et al. Direct seeding for rice production increased soil erosion and phosphorus runoff losses in subtropical China. Sci. Total. Environ. 2019, 695, 133845. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, F.; Abalos, D.; Luo, Y.; Hui, D.; Hungate, B.A.; García-Palacios, P.; Kuzyakov, Y.; Olesen, J.E.; Jørgensen, U.; et al. Stimulation of ammonia oxidizer and denitrifier abundances by nitrogen loading: Poor predictability for increased soil N2O emission. Glob. Chang. Biol. 2021, 28, 2158–2168. [Google Scholar] [CrossRef]
- Yang, Z.-P.; Zheng, S.-X.; Nie, J.; Liao, Y.-L.; Xie, J. Effects of Long-Term Winter Planted Green Manure on Distribution and Storage of Organic Carbon and Nitrogen in Water-Stable Aggregates of Reddish Paddy Soil Under a Double-Rice Cropping System. J. Integr. Agric. 2014, 13, 1772–1781. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, T.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cooper, J.M.; Lin, Z.; Li, Y.; Yang, X.; Zhao, B. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Chen, J.; Moorhead, D.L. Progressively decreased nitrogen-stimulation of soil phosphatase activity with long-term nitrogen addition. Appl. Soil Ecol. 2021, 169, 104213. [Google Scholar] [CrossRef]
- Liu, W.; Graham, E.B.; Zhong, L.; Zhang, J.; Li, S.; Lin, X.; Feng, Y. Long-Term Stochasticity Combines with Short-Term Variability in Assembly Processes to Underlie Rice Paddy Sustainability. Front. Microbiol. 2020, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Y.; Kuzyakov, Y.; Wang, D.; Olesen, J.E. Challenges in upscaling laboratory studies to ecosystems in soil microbiology research. Global Change Biol. 2023, 29, 569–574. [Google Scholar] [CrossRef]
- Chen, J.; Sinsabaugh, R.L. Linking microbial functional gene abundance and soil extracellular enzyme activity: Implications for soil carbon dynamics. Glob. Change Biol. 2020, 27, 1322–1325. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Early Rice (kg ha−1 y−1) | Late Rice (kg ha−1 y−1) | Milk Vetch | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Milk Vetch Manure | Chemical Fertilizers | Farmyard Manure | Chemical Fertilizers | Rice Straw | |||||
| N | P2O5 | K2O | N | P2O5 | K2O | ||||
| Control | - | - | - | - | - | - | - | - | - |
| M1 | 2250 | 45 a, 45 b, 69 c | 30 a,b,c | 0 a, 37.5 b, 75 c | - | 45 a, 45 b, 69 c | 30 a,b,c | 0 a, 37.5 b, 75 c | - |
| M2 | 2250 | 45 a, 45 b, 69 c | 30 a,b,c | 0 a, 37.5 b, 75 c | 11,250 | 45 a, 45 b, 69 b | 30 a,b,c | 0 a, 37.5 b, 75 c | 4050 |
| NPK | - | 90 | 45 | 75 | - | 90 a, 45 b, 69 c | 60 a, 30 b, 30 c | 0 a, 37.5 b, 75 c | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Liu, K.-L.; Chen, J.; Xie, J.; Jiang, Y.; Deng, G.-Q.; Li, D.-M.; Guan, X.-J.; Liang, X.-H.; Chen, X.-M.; et al. New Insights from Soil Microorganisms for Sustainable Double Rice-Cropping System with 37-Year Manure Fertilization. Agronomy 2023, 13, 261. https://doi.org/10.3390/agronomy13010261
Li J, Liu K-L, Chen J, Xie J, Jiang Y, Deng G-Q, Li D-M, Guan X-J, Liang X-H, Chen X-M, et al. New Insights from Soil Microorganisms for Sustainable Double Rice-Cropping System with 37-Year Manure Fertilization. Agronomy. 2023; 13(1):261. https://doi.org/10.3390/agronomy13010261
Chicago/Turabian StyleLi, Jin, Kai-Luo Liu, Ji Chen, Jiang Xie, Yu Jiang, Guo-Qiang Deng, Da-Ming Li, Xian-Jiao Guan, Xi-Huang Liang, Xian-Mao Chen, and et al. 2023. "New Insights from Soil Microorganisms for Sustainable Double Rice-Cropping System with 37-Year Manure Fertilization" Agronomy 13, no. 1: 261. https://doi.org/10.3390/agronomy13010261