
Development of Transgenic Maize Tolerant to Both Glyphosate and Glufosinate

Abstract
:1. Introduction
2. Materials and Methods
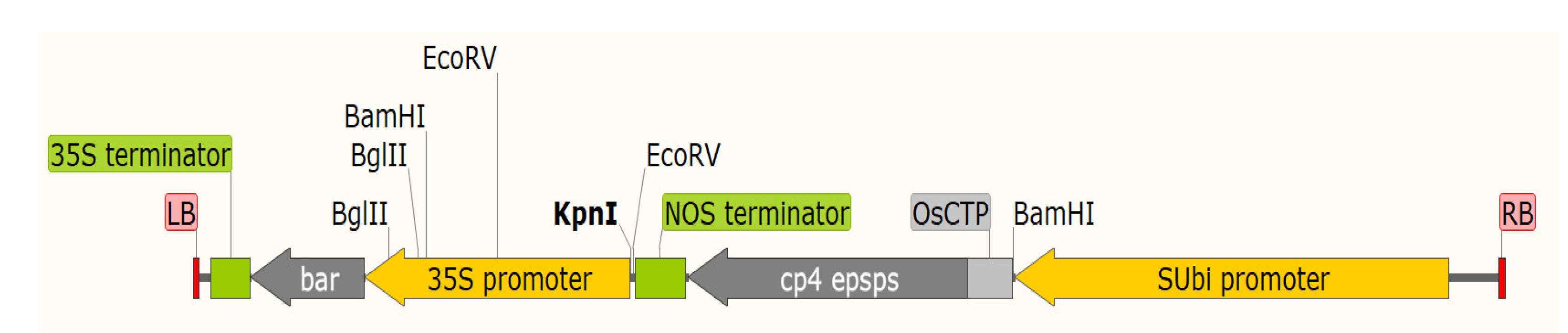
2.1. Maize Transformation and Production of Event SCB-29 Corn
2.2. Amplification of the Unknown Flanking Sequences of T-DNA
2.3. Establishment of Event-Specific PCR for SCB-29
2.4. Southern Blot Analysis
2.5. Western Blot Analysis
2.6. Stability Analysis of Transgenes
2.6.1. Gene-Specific and Event-Specific PCR Analysis
2.6.2. Protein Quantification of CP4 EPSPS and Bar
2.7. Target Trait Detection
2.8. Agronomic Performances of Transgenic Maize with Different Herbicide Treatments
2.9. Weed Control Efficiency
3. Results
3.1. Flanking Sequences Analysis and Event-Specific Detection for Event SCB-29
3.2. Analysis of T-DNA Copy Number in Transgenic Maize SCB-29
3.3. Expression of CP4 EPSPS and Bar in SCB-29
3.4. Genetic Stability of SCB-29 Corn over Three Generations
3.5. Herbicide Tolerance Analysis of SCB-29
3.6. Agronomic Performances of Transgenic Maize SCB-29 with Different Herbicide Treatments
3.7. Weed Control Efficiency and Maize Yield between Different Treatments
4. Discussions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Awika, J.M. Major cereal grains production and use around the world. In Advances in Cereal Science: Implications to Food Processing and Health Promotion; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2011; Volume 1089, pp. 1–13. [Google Scholar]
- Orhun, G.E. Maize for life. Int. J. Food Sci. Nutr. 2013, 3, 13–16. [Google Scholar]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, A.J.; Burke, I.C.; Everman, W.J.; Vangessel, M.J.; Davis, V.M.; Sikkema, P.H. Potential corn yield losses due to weeds in North America. Weed Technol. 2016, 30, 979–984. [Google Scholar] [CrossRef]
- Kannan, S.; Chinnagounder, C. Evaluation of weed control options for herbicide resistant transgenic stacked (TC 1507 X NK603) and conventional maize hybrids for higher productivity. Am. J. Plant Sci. 2013, 4, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.P. Development of chemical weed control and integrated weed management in China. Weed Biol. Manag. 2010, 3, 197–203. [Google Scholar] [CrossRef]
- Rodenburg, J.; Saito, K.; Irakiza, R.; Makokha, D.W.; Onyuka, E.A.; Senthilkumar, K. Labor-saving weed technologies for lowland rice farmers in sub-Saharan Africa. Weed Technol. 2015, 29, 751–757. [Google Scholar] [CrossRef] [Green Version]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier. In ISAAA Brief No. 55; Cornell University Ithaca: New York, NY, USA, 2019. [Google Scholar]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2010, 64, 319–325. [Google Scholar] [CrossRef]
- Steinrücken, H.C.; Amrhein, N. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvyl-shikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun. 1980, 94, 1207–1212. [Google Scholar] [CrossRef]
- Funke, T.; Han, H.; Healy-Fried, M.L.; Fischer, M.; Schoenbrunn, E. Molecular basis for the herbicide resistance of Roundup Ready crops. PNAS 2006, 103, 13010–13015. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.M.; Kroes, R.; Munro, I.C. Safety evaluation and risk assessment of the herbicide Roundup and its active ingredient, glyphosate, for humans. Regul. Toxicol. Pharm. 2000, 31, 117–165. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.J.; Cao, G.Y.; Zhang, Y.W.; Liu, Y.; Liu, Y.J. Overexpression of a modified AM79 aroA gene in transgenic maize confers high tolerance to glyphosate. J. Integr. Agric. 2015, 14, 414–422. [Google Scholar] [CrossRef]
- Liu, Y.J.; Zhang, Y.W.; Liu, Y.; Lu, W.; Wang, G.Y. Metabolic effects of glyphosate on transgenic maize expressing a G2-EPSPS gene from Pseudomonas fluorescens. J. Plant Biochem. Biot. 2015, 24, 233–241. [Google Scholar] [CrossRef]
- Jerry, M.G.; Theresa, H.; Pagano, M.A.; Andreassi, J.L., II; Gutteridge, S.A. Response of 98,140 corn with Gat4621 and hra transgenes to glyphosate and ALS-Inhibiting herbicides. Weed Sci. 2009, 57, 142–148. [Google Scholar]
- Hernández, M.; Esteve, T.; Prat, S.; Pla, M. Development of real-time PCR systems based on SYBR Green I, Amplifluor and TaqMan technologies for specific quantitative detection of the transgenic maize event GA21. J. Cereal Sci. 2004, 39, 99–107. [Google Scholar] [CrossRef]
- Matthews, B.A.; Launis, K.L.; Bauman, P.A.; Juba, N.C. Double-mutated 5-Enol Pyruvylshikimate-3-phosphate synthase protein expressed in MZHG0JG Corn (Zea mays L.) has no impact on toxicological safety and nutritional composition. J. Agric. Food Chem. 2017, 65, 8459–8465. [Google Scholar] [CrossRef]
- Mesnage, R.; Agapito-Tenfen, S.Z.; Vilperte, V.; Renney, G.; Ward, M.; Séralini, G.; Nodari, R.O.; Antoniou, M.N. An integrated multi-omics analysis of the NK603 Roundup-tolerant GM maize reveals metabolism disturbances caused by the transformation process. Sci. Rep. 2016, 6, 37855. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.M.; Zhang, X.J.; Gao, Y.; Shen, Z.C.; Lin, C.Y. Molecular characterization and efficacy evaluation of a transgenic corn event for insect resistance and glyphosate tolerance. J. Zhejiang Univ.- SC B 2018, 19, 610–619. [Google Scholar] [CrossRef]
- International Service for the Acquisition of Agri-biotech Applications (ISAAA). 2022. Available online: www.isaaa.org (accessed on 9 November 2022).
- Eelen, G.; Dubois, C.; Cantelmo, A.R.; Goveia, J.; Carmeliet, P. Role of glutamine synthetase in angiogenesis beyond glutamine synthesis. Nature 2018, 561, 63–69. [Google Scholar] [CrossRef]
- Liu, X.J.; Huan, Z.Y.; Zhang, Q.F.; Zhong, M.Q.; Du, H. Glutamine Synthetase (GS): A key enzyme for nitrogen assimilation in macroalga Gracilariopsis lemaneiformis (Rhodophyta). J. Phycol. 2019, 55, 1059–1070. [Google Scholar] [CrossRef]
- Kutlesa, N.J.; Caveney, S. Insecticidal activity of glufosinate through glutamine depletion in a caterpillar. Pest Manag. Sci. 2015, 57, 25–32. [Google Scholar] [CrossRef]
- Thompson, C.J.; Movva, N.R.; Tizard, R.; Crameri, R.; Botterman, J. Characterization of the herbicide-resistance gene bar from Streptomyces hygroscopicus. Embo J. 1987, 6, 2519–2523. [Google Scholar] [CrossRef]
- Wohlleben, W.; Arnold, W.; Broer, I.; Hillemann, D.; Strauch, E.; Pühler, A. Nucleotide sequence of phosphinothricin-N-acetyl-transferase gene from Streptomyces viridochromogenes Tue H 94 and its expression in Nicotiane tabacum. Gene 1988, 70, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Lutz, K.A.; Maliga, K.P. Expression of bar in the plastid genome confers herbicide resistance. Plant Physiol. 2001, 125, 1585–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florian, W.; Cristina, B.; Claudia, P.; Maddalena, Q.; Simon, K.; Marc, D.B.; Guy, V.D.E. Real-time polymerase chain reaction-based approach for quantification of the pat gene in the T25 Zea mays event. J. Aoac. Int. 2004, 87, 1342–1355. [Google Scholar]
- Milcamps, A.; Rabe, S.; Cade, R.; De Framond, A.J.; Henriksson, P.; Kramer, V.; Lisboa, D.; Pastor-Benito, S.; Willits, M.G.; Lawrence, D. Validity assessment of the detection method of maize event Bt10 through investigation of its molecular structure. J. Agric. Food Chem. 2009, 57, 3156–3163. [Google Scholar] [CrossRef]
- Windels, P.; Bertrand, S.; Depicker, A.; Moens, W.; Bockstaele, E.V.; Loose, M. Qualitative and event-specific PCR real-time detection methods for StarLink maize. Eur. Food Res. Technol. 2003, 216, 259–263. [Google Scholar] [CrossRef]
- Herman, R.A.; Phillips, A.M.; Collins, R.A.; Tagliani, L.A.; Prochaska, L.M. Compositional equivalency of Cry1F corn event TC6275 and conventional corn (Zea mays L.). J. Agric. Food Chem. 2004, 52, 2726. [Google Scholar] [CrossRef]
- Heap. I. The International Herbicide-Resistant Weed Database. 2022. Available online: www.weedscience.org (accessed on 9 November 2022).
- Beckie, H.J. Herbicide-resistant weeds: Management tactics and practices. Weed Technol. 2006, 20, 793–814. [Google Scholar] [CrossRef]
- Shih, M.H.; Doran, P.M. Foreign protein production using plant cell and organ cultures: Advantages and limitations. Biotechnol. Adv. 2009, 27, 1036–1042. [Google Scholar] [CrossRef]
- Fearing, P.L.; Brown, D.; Valchos, D.; Meghji, M.; Privalle, L. Quantitative analysis of Cry1A(b) expression in Bt maize plants, tissues, and silage and stability of expression over successive generations. Mol. Breed. 1997, 3, 169–176. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Jehle, J.A. Quantitative analysis of the seasonal and tissue-specific expression of Cry1Ab in transgenic maize Mon810. J. Plant Dis. Protect 2007, 114, 82–87. [Google Scholar] [CrossRef]
- Gelvin, S.B. Integration of Agrobacterium T-DNA into the plant genome. Annu. Rev. Genet. 2017, 51, 195–217. [Google Scholar] [CrossRef]
- Frame, B.R.; Shou, H.; Chikwamba, R.K.; Zhang, Z.; Xiang, C.; Fonger, T.M.; Pegg, S.; Li, B.; Dan, S.N.; Wang, P.K. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol. 2002, 129, 13–22. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.G.; Chen, Y. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–656. [Google Scholar] [CrossRef]
- Gianessi, L.P. Economic and herbicide use impacts of glyphosate-resistant crops. Pest Manag. Sci. 2005, 61, 241–245. [Google Scholar] [CrossRef]
- Gianessi, L.P. Economic impacts of glyphosate-resistant crops. Pest Manag. Sci. 2008, 64, 346–352. [Google Scholar] [CrossRef]
- Owen, M.D. Diverse approaches to herbicide-resistant weed management. Weed Sci. 2016, 64, 570–584. [Google Scholar] [CrossRef] [Green Version]
- Beckie, H.J. Herbicide-resistant weed management: Focus on glyphosate. Pest Manag. Sci. 2011, 67, 1037–1048. [Google Scholar] [CrossRef]
- Dhirendra, F.; Aakrati, A.; Donald, J.; Bhabesh, B.; Babu, R.; Vijay, S.; Agrawal, P.K.; Murali, A.; Reddy, M.K. Developing dual herbicide tolerant transgenic rice plants for sustainable weed management. Sci. Rep. 2018, 8, 11598. [Google Scholar]
- Bhat, S.; Chopra, V. Choice of technology for herbicide-resistant transgenic crops in India: Examination of issues. Curr. Sci. 2006, 91, 435–438. [Google Scholar]
- Bonny, S. Genetically modified herbicide-tolerant crops, weeds, and herbicides: Overview and impact. Environ. Manag. 2016, 57, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Guo, M.X.; Liu, H.L.; Zhang, C.W.; Ding, D.R.; Jiao, G.W.; Wei, X.S.; Tang, B.; Xia, Z.L.; Xiong, G.J.; et al. Herbicide-tolerant Maize Plant DBN9858, and Nucleotide Sequence and Method for Detecting Same. 2016. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2016173508 (accessed on 9 November 2022).
- Ellis, C.M.; Goley, M.E.; Huang, J.; Klingaman, T.E.; Larue, C.T.; Qi, Y.; Sparks, O.C.; Van Scoyoc, B.M.; Yang, H. Maize event MON87429 and Methods of Use Thereof. U.S. Patent No. 10,920,239, 16 February 2021. [Google Scholar]
- Mehrotra, S.; Goyal, V. Evaluation of designer crops for biosafety-A scientist’s perspective. Gene 2012, 515, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.F.; Jin, X.; Tang, Q.L.; Zhang, X.; Yang, J.T.; Liu, X.J.; Cai, J.F.; Zhang, X.B.; Wang, X.J.; Wang, Z.X. Development and event-specific detection of trasngenic glyphosate-resistant rice expressing the G2-EPSPS gene. Front. Plant Sci. 2017, 8, 885. [Google Scholar] [CrossRef] [Green Version]
- Koger, C.H.; Price, A.J.; Faircloth, J.C.; Wilcut, J.W.; Nichols, S.P. Effect of residual herbicides used in the last post-directed application on weed control and cotton yield in glyphosate- and glufosinate-resistant cotton. Weed Technol. 2007, 21, 378–383. [Google Scholar] [CrossRef]
- Smith, A.E. Persistence and transformation of the herbicide [14C] glufosinate-ammonium in prairie soils under laboratory conditions. J. Agric. Food Chem. 1988, 36, 393–397. [Google Scholar] [CrossRef]
- Tebbe, C.C.; Reber, H.H. Degradation of [14C] phosphinothricin (glufosinate) in soil under laboratory conditions: Effects of concentration and soil amendments on 14CO2 production. Biol. Fertil. Soils 1991, 11, 62–67. [Google Scholar] [CrossRef]
- Torstensson, N.T.L.; Aamisepp, A. Detoxification of glyphosate in soil. Weed Res. 1977, 17, 209–212. [Google Scholar] [CrossRef]
- Komarlingam, M.S. Refuge-in-bag for Bt cotton. Curr. Sci. 2018, 114, 726–727. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5’-3’) | Amplicon (bp) |
|---|---|---|
| LB-MG | 5′-GTGCCAAAGTAGCCCTAAAACGGCC-3′ | 478 |
| LB-TDNA | 5′-TTTCTCCATAATAATGTGTGAGTAGTTCCC-3′ | |
| RB-TDNA | 5′-CGTGACTGGGAAAACCCTGGCGTT-3′ | 416 |
| RB-MG | 5′-CATCGCAACCAGCGTGTGCGTAC-3′ | |
| zSSIIb-F | 5′-CTCCCAATCCTTTGACATCTGC-3′ | 151 |
| zSSIIb-R | 5′-TCGATTTCTCTCTTGGTGACAGG-3′ | |
| CP4-GF | 5′-CAGGATTCCGGGCGATAAGTCCATATC-3′ | 1068 |
| CP4-GR | 5′-GAGGTCTCACCCTCGTCGCAGTCAAC-3′ | |
| Bar-GF | 5′-TGCACCATCGTCAACCACTACATCGAG-3′ | 455 |
| Bar-GR | 5′-GGTACAGGCAGGCTGAAGTCCAGCTG-3′ |
| Treatments | Weed Control Efficiency (%) | Maize Yield (t/ha) |
|---|---|---|
| SCB-29 with glyphosate at 900 g a.e. ha−1 | 59.16 c | 8.49 |
| SCB-29 with glufosinate at 900 g a.i. ha−1 | 79.14 b | 8.58 |
| SCB-29 with gly at 900 followed glu at 900 | 96.43 a | 8.83 |
| SCB-29 with glu at 900 followed gly at 900 | 95.66 a | 8.62 |
| SCB-29 with no herbicide spraying and no manual weeding | — | 6.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Sun, Y.; Lin, C.; Wang, P.; Shen, Z.; Zhao, Y. Development of Transgenic Maize Tolerant to Both Glyphosate and Glufosinate. Agronomy 2023, 13, 226. https://doi.org/10.3390/agronomy13010226
Yu X, Sun Y, Lin C, Wang P, Shen Z, Zhao Y. Development of Transgenic Maize Tolerant to Both Glyphosate and Glufosinate. Agronomy. 2023; 13(1):226. https://doi.org/10.3390/agronomy13010226
Chicago/Turabian StyleYu, Xiaoxing, Yongzheng Sun, Chaoyang Lin, Pengfei Wang, Zhicheng Shen, and Yu Zhao. 2023. "Development of Transgenic Maize Tolerant to Both Glyphosate and Glufosinate" Agronomy 13, no. 1: 226. https://doi.org/10.3390/agronomy13010226