
Cloning and Disease Resistance Analysis of the Maize ZmBON3 Gene
 ,
,
Abstract
:1. Introduction
2. Results
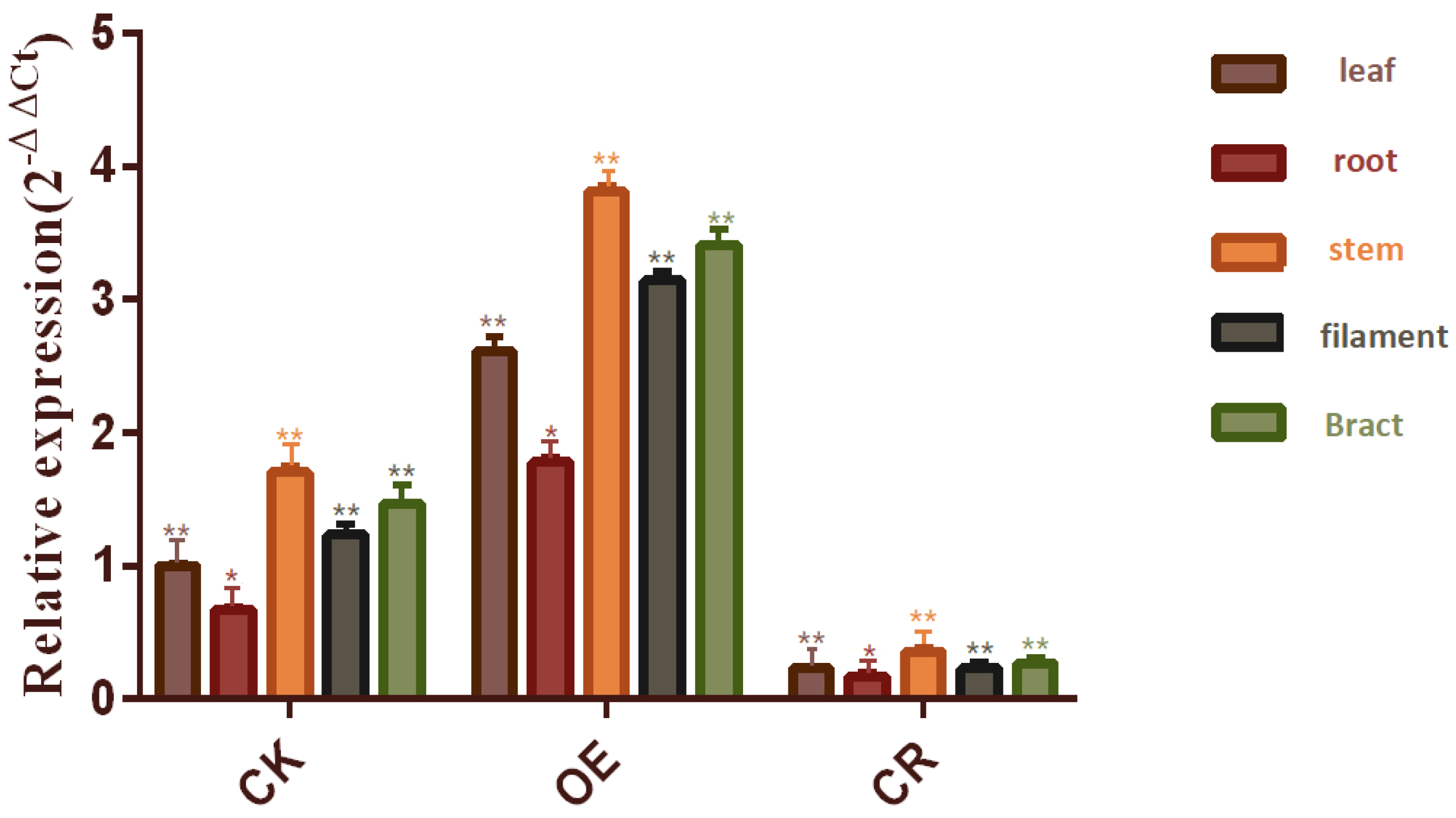
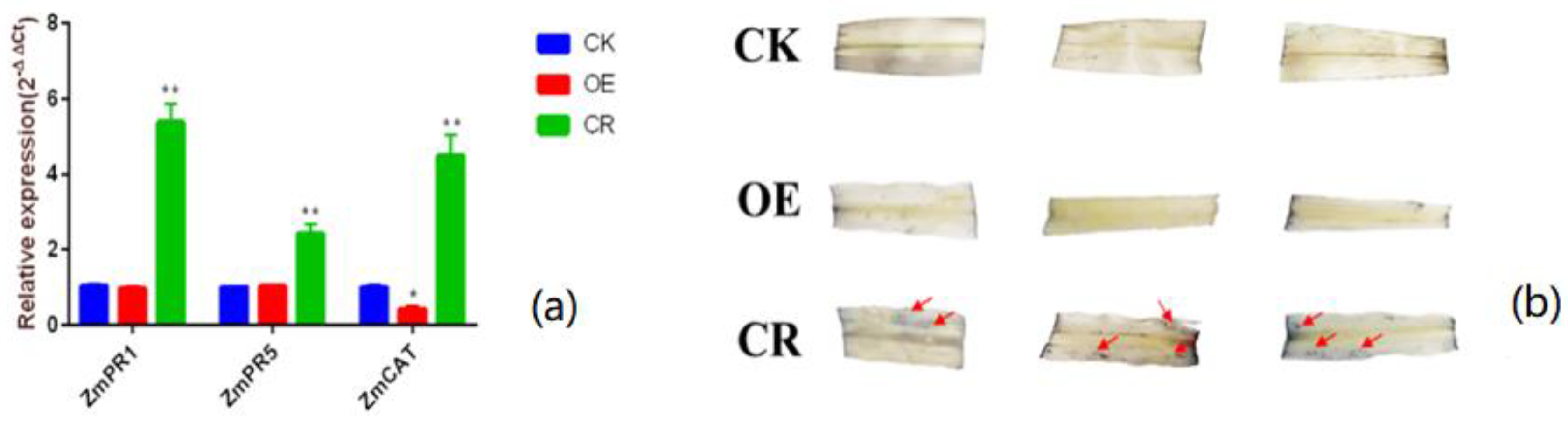
2.1. Detection of Transgenic Plants by Fluorescence Quantitative PCR
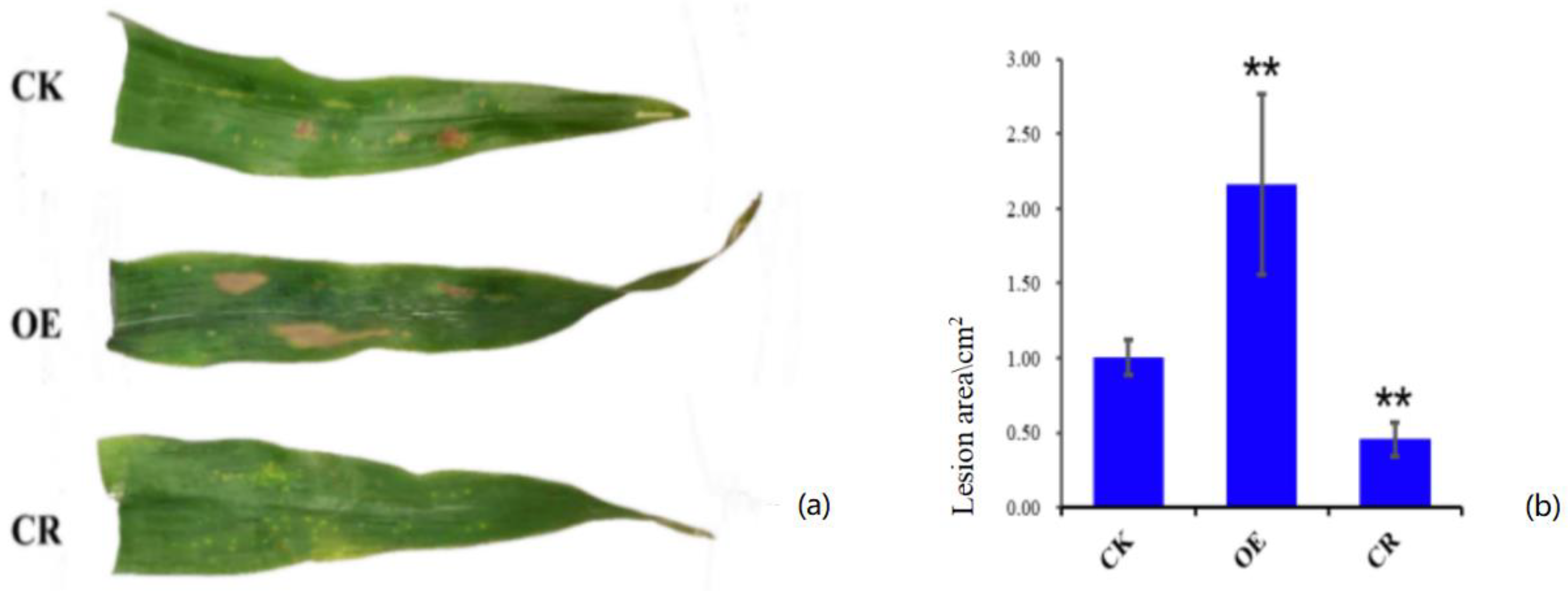
2.2. Effect of ZmBON3 on Disease Resistance of Maize Plants
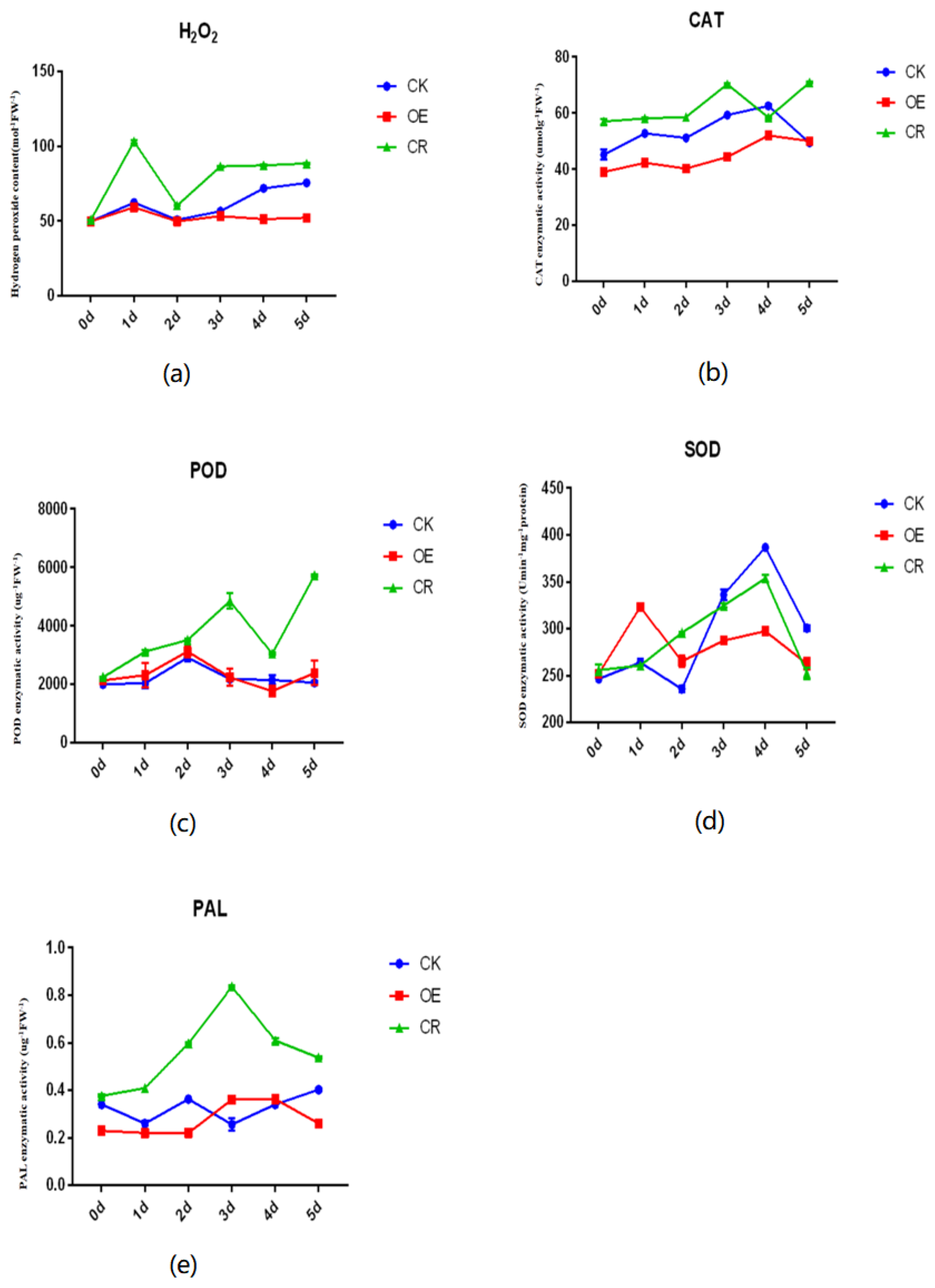
2.3. Changes in Physiological and Biochemical Parameters Related to Defense in Transgenic Plants
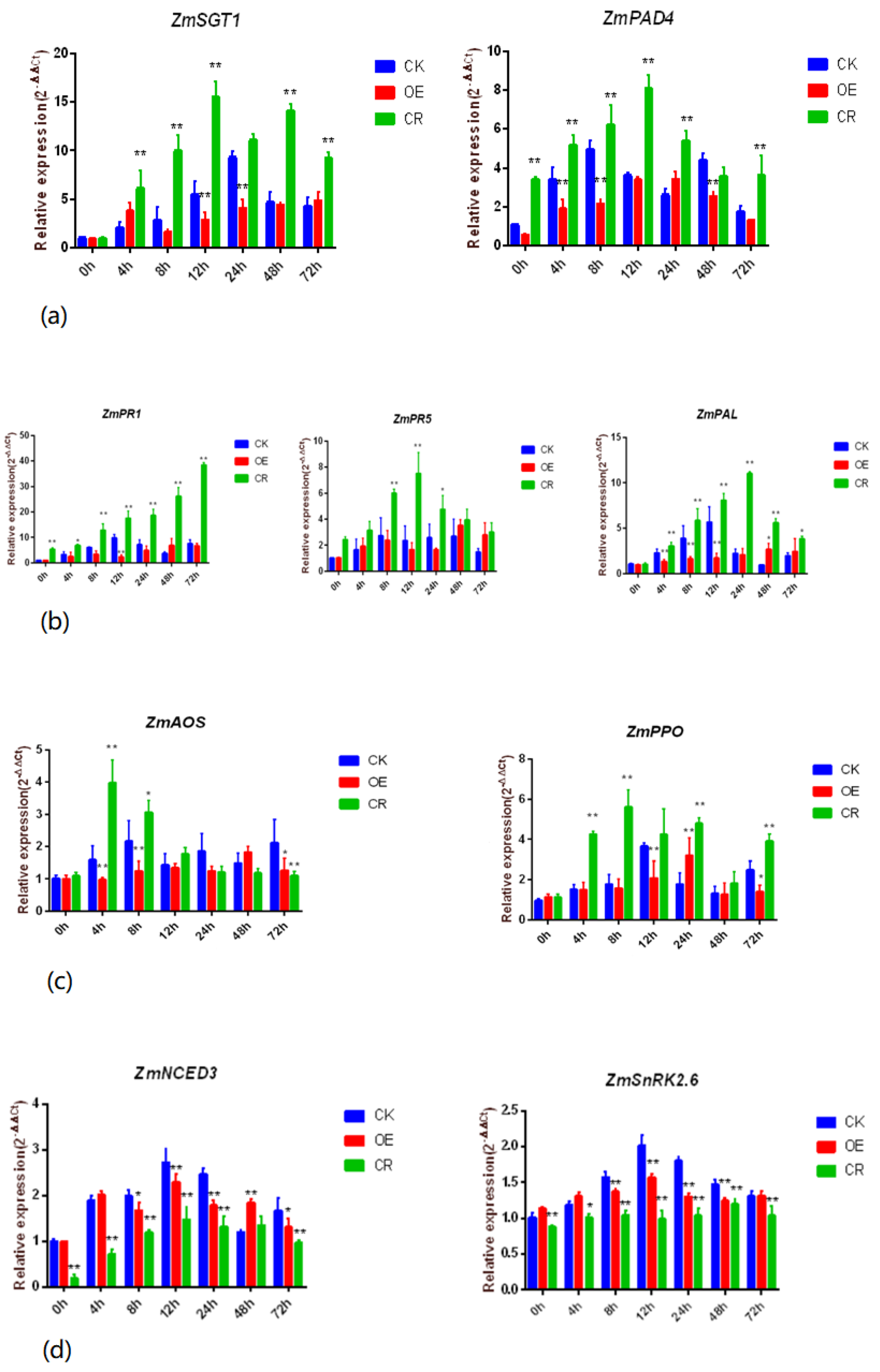
2.4. Changes in Transcription Levels of Key Genes Related to Disease Resistance Pathways
2.5. Effect of ZmBON3 on the Spontaneous Immunity of Maize
2.6. Investigation on Agronomic Characters of Transgenic Plants
3. Discussion
4. Materials and Methods
4.1. Production of Transgenic Plants
4.2. Culture and Inoculation of Northern Corn Leaf Blight
4.3. The Transgenic Plants Were Subjected to Real-Time PCR
4.4. Changes in Physiological and Biochemical Parameters Related to Defense in Transgenic Plants
4.5. Spontaneous Immune Detection and Fluorescence Quantitative Detection of Transgenic Plants
4.6. Changes in Transcription Levels of Key Genes in Pathways Related to Disease Resistance
4.7. Investigation on Agronomic Characters of Transgenic Plants
4.8. Statistics and Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jiao, P.; Jiang, Z.; Wei, X.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Overexpression of the homeobox-leucine zipper protein ATHB-6 improves the drought tolerance of maize (Zea mays L.). Plant Sci. 2022, 316, 111159. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.G.; Jabloun, M.; Olesen, J.E.; Öztürk, I.; Wang, M.; Chen, F. Effects of climatic factors, drought risk and irrigation requirement on maize yield in the Northeast Farming Region of China. J. Agric. Sci. 2016, 154, 1171–1189. [Google Scholar] [CrossRef]
- Wu, F.; Shu, J.; Jin, W. Identification and validation of miRNAs associated with the resistance of maize (Zea mays L.) to Exserohilum turcicum. PLoS ONE 2014, 9, e87251. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, H.; Deutsch, J.A.; Gutierrez, H. Recurrent Selection for Resistance to Exserohilum turcicum in Eight Subtropical Maize Populations. Crop Sci. 1991, 31, 964–971. [Google Scholar] [CrossRef]
- Wise, K. Diseases of Corn: Northern Corn Leaf Blight; Purdue University Extension Publication; Purdue University: West Lafayette, IN, USA, 2011. [Google Scholar]
- Ullstrup, A.J. The Impacts of the Southern Corn Leaf Blight Epidemics of 1970–1971. Annu. Rev. Phytopathol. 1972, 10, 37–50. [Google Scholar] [CrossRef]
- Brown, J.K. Durable resistance of crops to disease: A Darwinian perspective. Annu. Rev. Phytopathol. 2015, 53, 513–539. [Google Scholar] [CrossRef]
- Macho, A.P.; Zipfel, C. Plant PRRs and the activation of innate immune signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef]
- Van Schie, C.C.; Takken, F.L. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Belkhadir, Y.; Nimchuk, Z.; Hubert, D.A.; Mackey, D.; Dangl, J.L. Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 2004, 16, 2822–2835. [Google Scholar] [CrossRef] [Green Version]
- Takken, F.L.; Goverse, A. How to build a pathogen detector: Structural basis of NB-LRR function. Curr. Opin. Plant Biol. 2012, 15, 375–384. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Christopoulou, M.; Caldwell, K.S. Impacts of resistance gene genetics, function, and evolution on a durable future. Annu. Rev. Phytopathol. 2013, 51, 291–319. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.; Albrecht, M.; Cornelissen, B.J.; Takken, F.L. Structure-function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef] [Green Version]
- Caplan, J.; Padmanabhan, M.; Dinesh-Kumar, S.P. Plant NB-LRR Immune Receptors: From Recognition to Transcriptional Reprogramming. Cell Host Microbe 2008, 3, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Jambunathan, N.; McNellis, T.W. Transgenic expression of the von Willebrand A domain of the BONZAI 1/COPINE 1 protein triggers a lesion-mimic phenotype in Arabidopsis. Plata 2005, 221, 85–94. [Google Scholar] [CrossRef]
- Lee, T.F.; Mcnellis, T.W. Evidence that the BONZAI1/COPINE1 protein is a calcium- and pathogen-responsive defense suppressor. Plant Mol. Biol. 2009, 69, 155–166. [Google Scholar] [CrossRef]
- Jambunathan, N. Genetic and Molecular Analysis of the Arabidopsis Copine I gene in Plant Defense Responses; The Pennsylvania State University: State College, PA, USA, 2003. [Google Scholar]
- Yin, X.; Zou, B.; Hong, X.; Gao, M.; Yang, W.; Zhong, X.; He, Y.; Kuai, P.; Lou, Y.; Huang, J.; et al. Rice copine genes OsBON1 and OsBON3 function as suppressors of broad-spectrum disease resistance. Plant Biotechnol. J. 2018, 16, 1476–1487. [Google Scholar] [CrossRef]
- Zou, B.; Ding, Y.; Liu, H.; Hua, J. Silencing of copine genes confers common wheat enhanced resistance to powdery mildew. Mol. Plant Pathol. 2018, 19, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gou, M.; Sun, Q.; Hua, J. Requirement of calcium binding, myristoylation, and protein-protein interaction for the Copine BON1 function in Arabidopsis. J. Biol. Chem. 2010, 285, 29884–29891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, J.; Grisafi, P.; Cheng, S.H.; Fink, G.R. Plant growth homeostasis is controlled by the Arabidopsis BON1 and BAP1 genes. Genes Dev. 2001, 15, 2263–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomsig, J.L.; Creutz, C.E. Copines: A ubiquitous family of Ca2+-dependent phospholipid-binding proteins. Cell. Mol. Life Sci. 2002, 59, 1467–1477. [Google Scholar] [CrossRef]
- Liu, P.; Khvotchev, M.; Li, Y.C.; Chanaday, N.L.; Kavalali, E.T. Copine-6 Binds to SNAREs and Selectively Suppresses Spontaneous Neurotransmission. J. Neurosci. 2018, 38, 5888–5899. [Google Scholar] [CrossRef]
- Damer, C.K.; Bayeva, M.; Kim, P.S.; Ho, L.K.; Eberhardt, E.S.; Socec, C.I.; Lee, J.S.; Bruce, E.A.; Goldman-Yassen, A.E.; Naliboff, L.C. Copine A is required for cytokinesis, contractile vacuole function, and development in Dictyostelium. Eukaryot. Cell 2007, 6, 430–442. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.Y.; Park, N.; Lee, B.; Choe, Y.I.; Woo, D.K.; Park, J.Y.; Yoo, J.C. CPNE1-mediated neuronal differentiation can be inhibited by HAX1 expression in HiB5 cells. Biochem. Biophys. Res. Commun. 2020, 533, 319–324. [Google Scholar] [CrossRef]
- Zou, B.; Hong, X.; Ding, Y.; Wang, X.; Liu, H.; Hua, J. Identification and analysis of copine/BONZAI proteins among evolutionarily diverse plant species. Genome 2016, 59, 565–573. [Google Scholar] [CrossRef]
- Jambunathan, N.; Siani, J.M.; Mcnellis, T.W. A humidity-sensitive Arabidopsis copine mutant exhibits precocious cell death and increased disease resistance. Plant Cell 2001, 13, 2225–2240. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Shen, X.; Tang, Y.; Shi, X.; Gu, Y. A karyopherin constrains nuclear activity of the NLR protein SNC1 and is essential to prevent autoimmunity in Arabidopsis. Mol. Plant 2021, 14, 1733–1744. [Google Scholar] [CrossRef]
- Yang, S.; Yang, H.; Grisafi, P.; Sanchatjate, S.; Fink, G.R.; Sun, Q.; Hua, J. The BON/CPN gene family represses cell death and promotes cell growth in Arabidopsis. Plant J. 2006, 45, 166–179. [Google Scholar] [CrossRef]
- Chen, K.; Gao, J.; Sun, S.; Zhang, Z.; Yu, B.; Li, J.; Xie, C.; Li, G.; Wang, P.; Song, C.P.; et al. BONZAI Proteins Control Global Osmotic Stress Responses in Plants. Curr. Biol. 2020, 30, 4815–4825. [Google Scholar] [CrossRef]
- Hamdoun, S.; Liu, Z.; Gill, M.; Yao, N.; Lu, H. Dynamics of defense responses and cell fate change during Arabidopsis-Pseudomonas syringae interactions. PLoS ONE 2013, 8, e83219. [Google Scholar] [CrossRef]
- Lulai, E.; Huckle, L.; Neubauer, J.; Suttle, J. Coordinate expression of AOS genes and JA accumulation: JA is not required for initiation of closing layer in wound healing tubers. J. Plant Physiol. 2011, 168, 976–982. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Qian, C.; Cao, A.; Li, Y.; Jiang, Z.; Li, M.; Jin, X.; Hu, J.; Zhang, Y.; Wang, X.; et al. The Hv-SGT1 gene from Haynaldia villosa contributes to resistances towards both biotrophic and hemi-biotrophic pathogens in common wheat (Triticum aestivum L.). PLoS ONE 2013, 8, e72571. [Google Scholar] [CrossRef]
- Gao, F.; Dai, R.; Pike, S.M.; Qiu, W.; Gassmann, W. Functions of EDS1-like and PAD4 genes in grapevine defenses against powdery mildew. Plant Mol. Biol. 2014, 86, 381–393. [Google Scholar] [CrossRef]
- Bozbuga, R. Expressions of Pathogenesis related 1 (PR1) Gene in Solanum lycopersicum and Influence of Salicylic Acid Exposures on Host-Meloidogyne incognita Interactions. Dokl. Biochem. Biophys. 2020, 494, 266–269. [Google Scholar] [CrossRef]
- Padmanabhan, C.; Ma, Q.; Shekasteband, R.; Stewart, K.S.; Hutton, S.F.; Scott, J.W.; Fei, Z.; Ling, K.S. Comprehensive transcriptome analysis and functional characterization of PR-5 for its involvement in tomato Sw-7 resistance to tomato spotted wilt tospovirus. Sci. Rep. 2019, 9, 7673. [Google Scholar] [CrossRef] [Green Version]
- Sussmilch, F.C.; Brodribb, T.J.; McAdam, S.A. Up-regulation of NCED3 and ABA biosynthesis occur within minutes of a decrease in leaf turgor but AHK1 is not required. J. Exp. Bot. 2017, 68, 2913–2918. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, P.; Guo, P.; Chong, L.; Yu, G.; Sun, X.; Hu, T.; Li, Y.; Hsu, C.C.; Tang, K.; et al. CDK8 is associated with RAP2.6 and SnRK2.6 and positively modulates abscisic acid signaling and drought response in Arabidopsis. New Phytol. 2020, 228, 1573–1590. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.E.; Lee, J.; Williams, S.J.; Choi, S.; Halane, M.K.; Zhou, J.; Solomon, P.; Kobe, B.; Jones, J.D.G.; Segonzac, C.; et al. Autoimmunity and effector recognition in Arabidopsis thaliana can be uncoupled by mutations in the RRS1-R immune receptor. New Phytol. 2019, 222, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Wang, Z.; Xu, L.; Peng, Q.; Liu, F.; Li, Z.; Que, Y. Early Selection for Smut Resistance in Sugarcane Using Pathogen Proliferation and Changes in Physiological and Biochemical Indices. Front. Plant Sci. 2016, 7, 1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garner, C.M.; Spears, B.J.; Su, J.; Cseke, L.J.; Smith, S.N.; Rogan, C.J.; Gassmann, W. Opposing functions of the plant TOPLESS gene family during SNC1-mediated autoimmunity. PLoS Genet. 2021, 17, e1009026. [Google Scholar] [CrossRef]
- Li, X.; Yang, D.L.; Sun, L.; Li, Q.; Mao, B.; He, Z. The Systemic Acquired Resistance Regulator OsNPR1 Attenuates Growth by Repressing Auxin Signaling through Promoting IAA-Amido Synthase Expression. Plant Physiol. 2016, 172, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Seedling Emergence | Hundred Grain Weight/g | Plant Height/cm | Ear Length/cm | Shaft Rough/cm | Grain Length/mm | Ear Rows | Grain Color |
|---|---|---|---|---|---|---|---|---|
| CK | 88% | 25.01 ± 0.71 | 152.66 ± 0.72 | 15.54 ± 0.49 | 3.38 ± 0.11 | 7.24 ± 0.38 | 14 | orange |
| OE | 63% | 26.29 ± 0.26 ** | 152.39 ± 0.77 | 15.25 ± 0.22 | 3.59 ± 0.13 | 8.55 ± 0.34 ** | 14 | orange |
| CR | 54% | 23.57 ± 0.49 ** | 146.82 ± 1.73 ** | 14.93 ± 0.33 | 3.56 ± 0.08 | 6.29 ± 0.17 * | 14 | orange |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Qi, Z.; Jiao, P.; Jiang, Z.; Liu, S.; Fan, S.; Zhao, C.; Ma, Y.; Guan, S. Cloning and Disease Resistance Analysis of the Maize ZmBON3 Gene. Agronomy 2023, 13, 152. https://doi.org/10.3390/agronomy13010152
Zhang C, Qi Z, Jiao P, Jiang Z, Liu S, Fan S, Zhao C, Ma Y, Guan S. Cloning and Disease Resistance Analysis of the Maize ZmBON3 Gene. Agronomy. 2023; 13(1):152. https://doi.org/10.3390/agronomy13010152
Chicago/Turabian StyleZhang, Chen, Zhuo Qi, Peng Jiao, Zhenzhong Jiang, Siyan Liu, Sujie Fan, Chunli Zhao, Yiyong Ma, and Shuyan Guan. 2023. "Cloning and Disease Resistance Analysis of the Maize ZmBON3 Gene" Agronomy 13, no. 1: 152. https://doi.org/10.3390/agronomy13010152