
Effect of the Altitude Gradient on the Physiological Performance of Quinoa in the Central Region of Colombia


Abstract
:1. Introduction
2. Materials and Methods
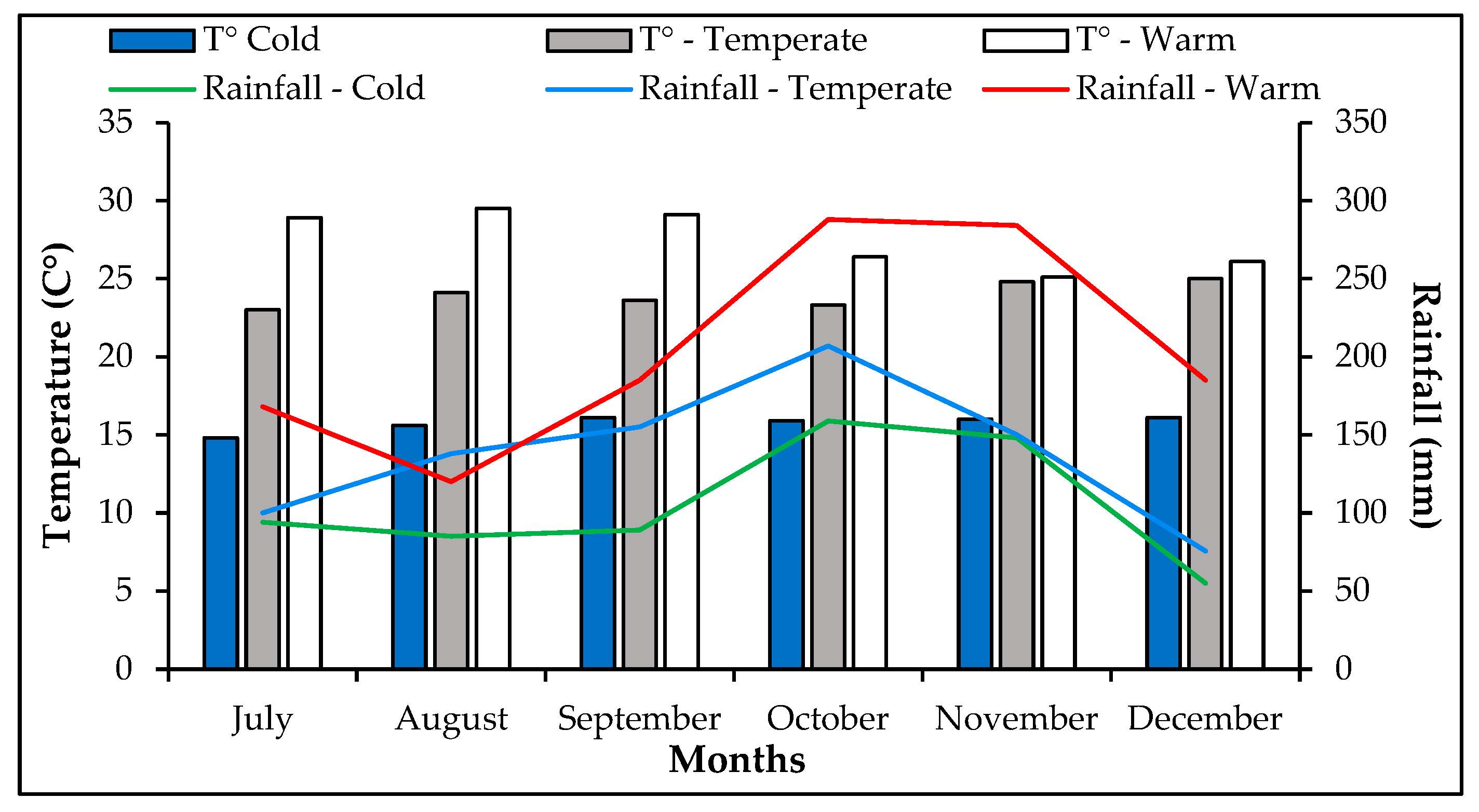
2.1. Environmental Conditions
2.2. Plant Growth and Development
2.3. Physiological Activity
2.4. Grain Yield
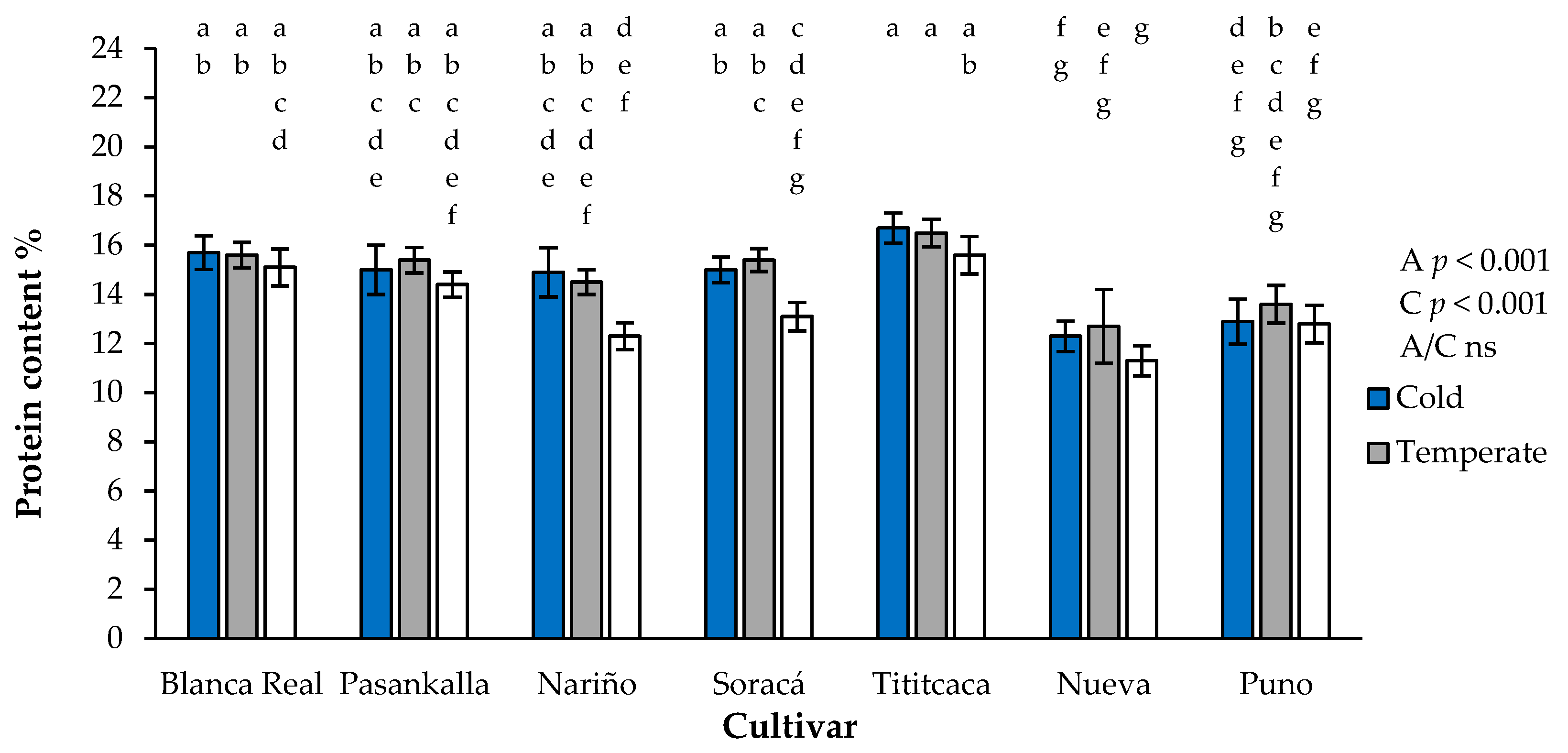
2.5. Protein Content
2.6. Statistical Analysis
3. Results and Discussions
3.1. Growth and Development
3.2. Total Chlorophyll Content
3.3. Chlorophyll Fluorescence (Fv/Fm)
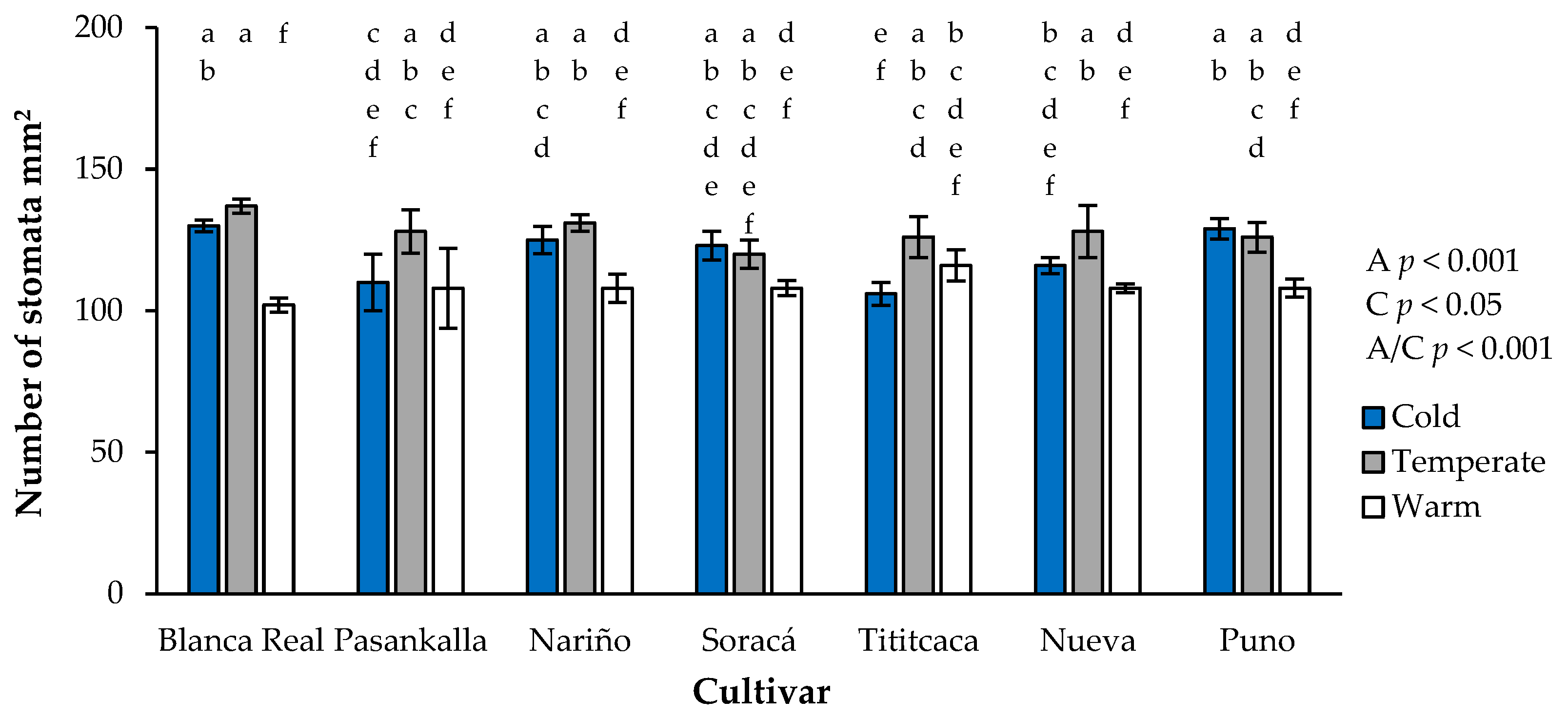
3.4. Stomatal Density
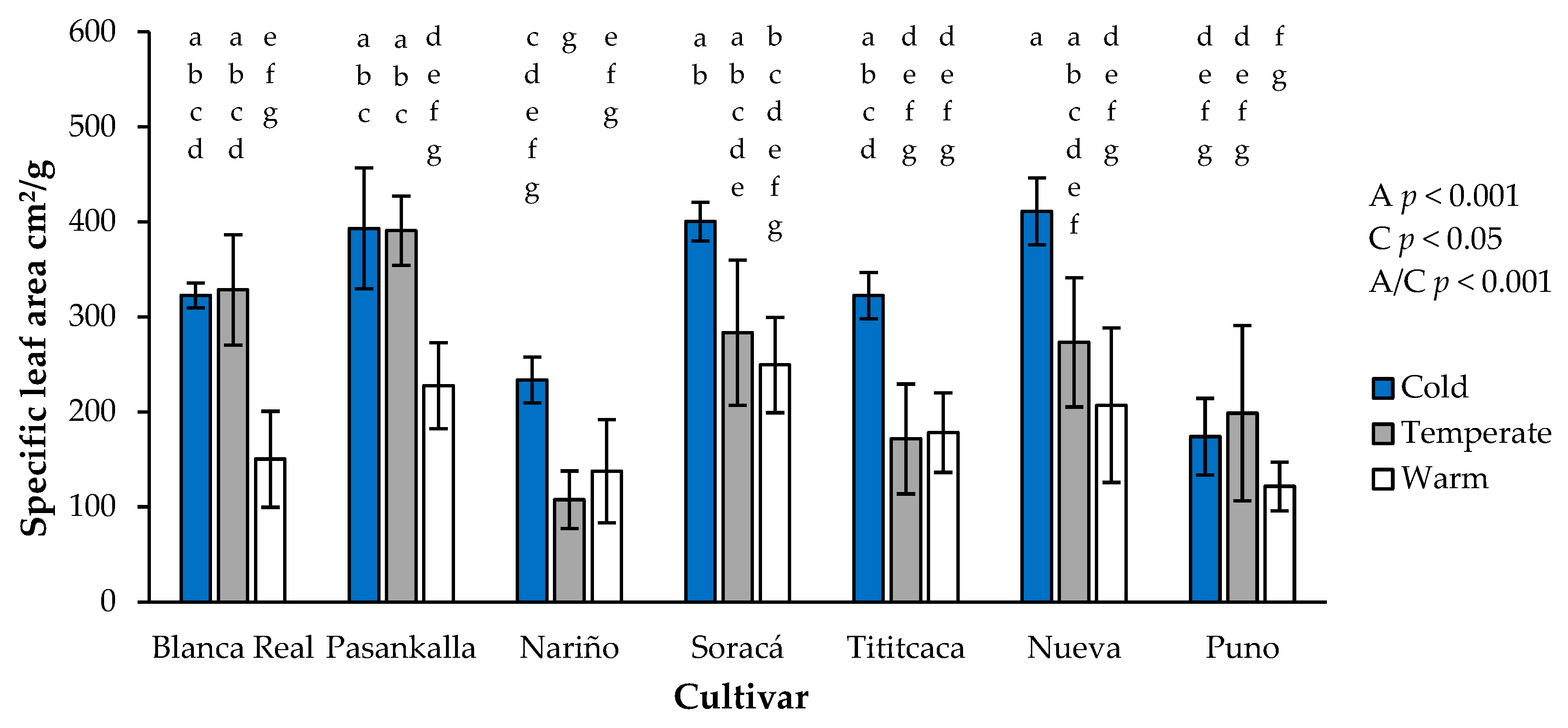
3.5. Specific Leaf Area
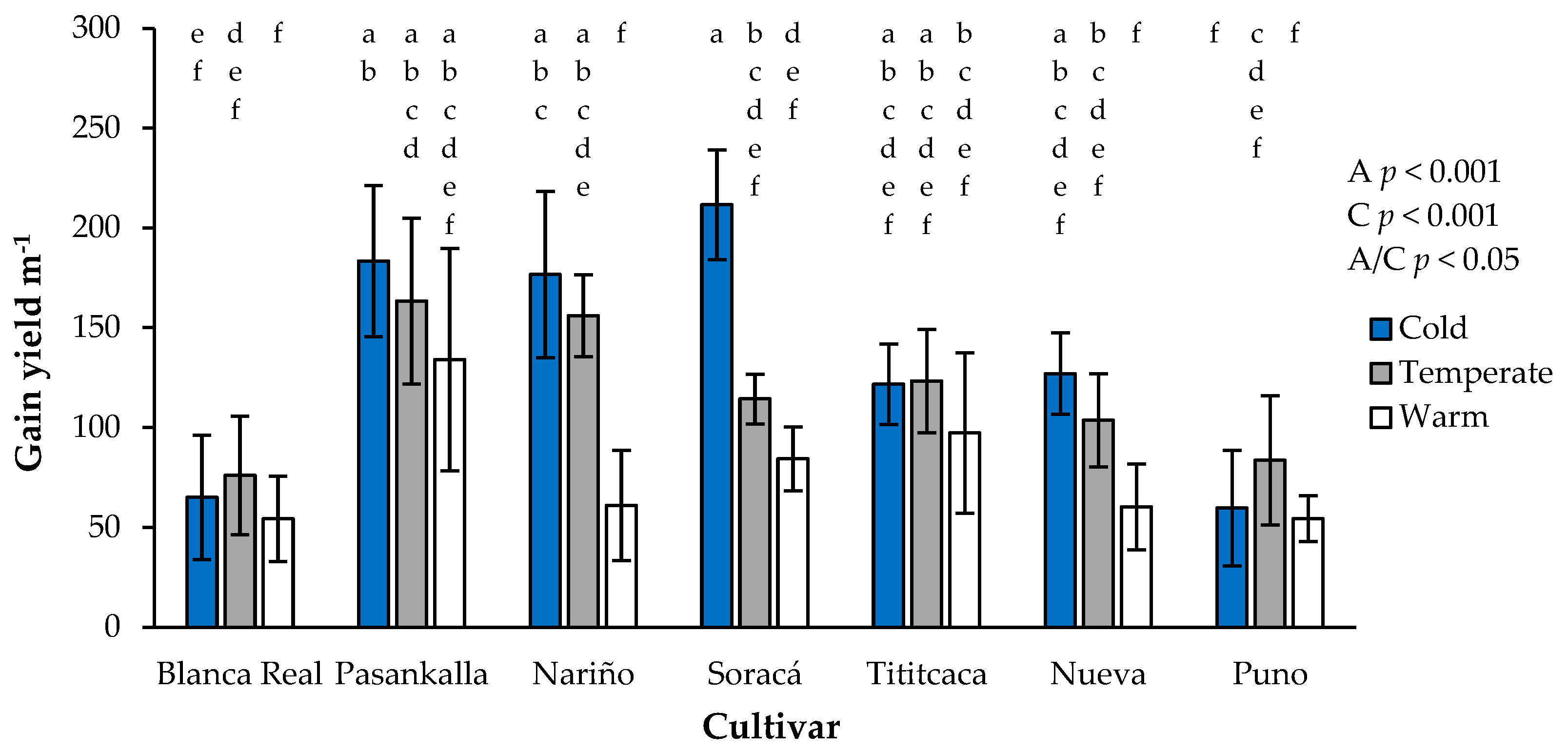
3.6. Grain Yield
3.7. Protein Content
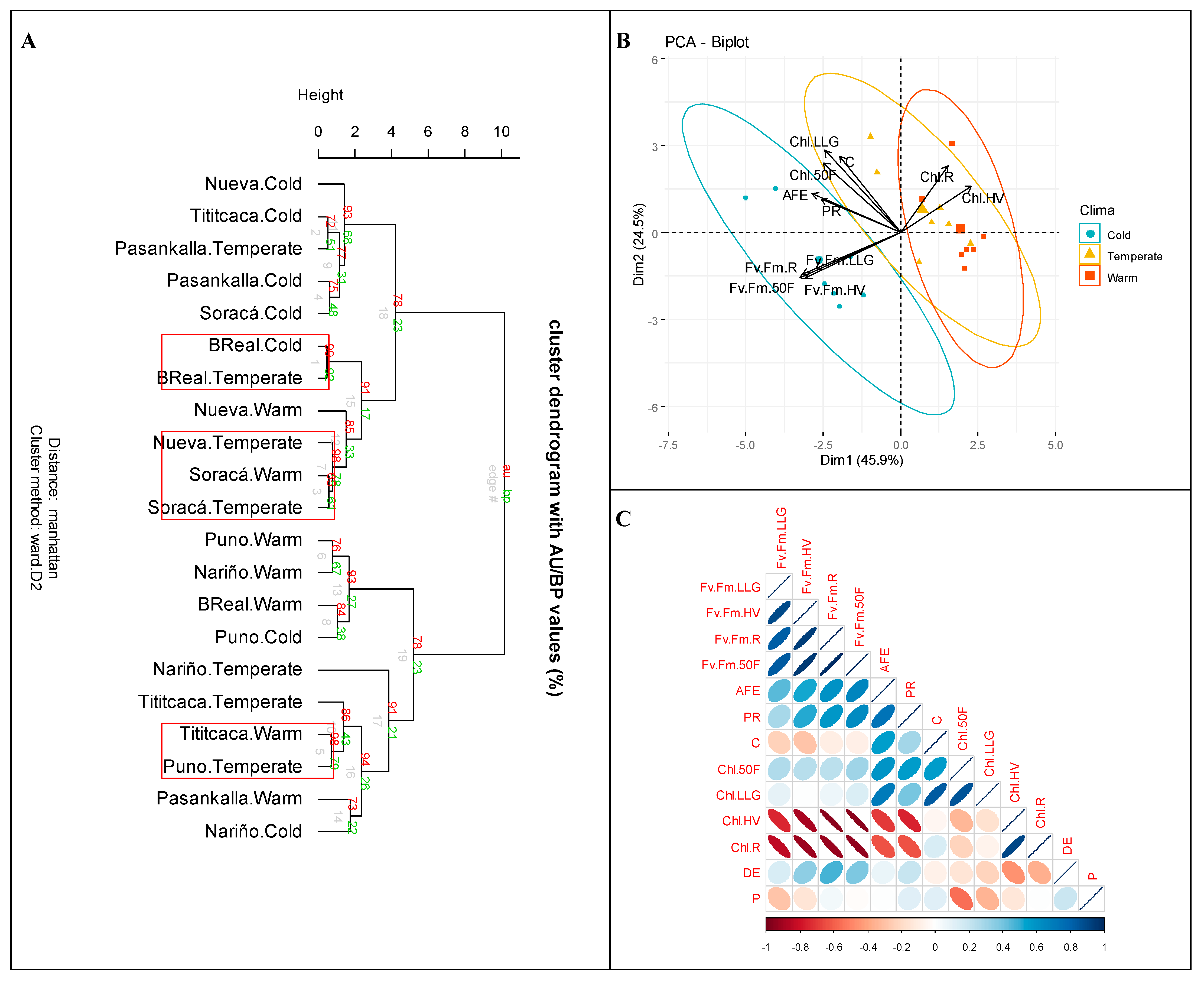
3.8. Altitudinal Gradient Relationship with Seven Quinoa Cultivars
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Parra, M.; Zurita-Silva, A.; Stechauner-Rohringer, R.; Roa-Acosta, D.; Jacobsen, S.-E. Quinoa (Chenopodium quinoa Willd.) and its relationship with agroclimatic characteristics: A Colombian perspective. Chil. J. Agric. Res. 2020, 80, 290–302. [Google Scholar] [CrossRef]
- Reguera, M.; Conesa, C.M.; Gil-Gómez, A.; Haros, C.M.; Pérez-Casas, M.Á.; Briones-Labarca, V.; Bolaños, L.; Bonilla, I.; Álvarez, R.; Pinto, K.; et al. The impact of different agroecological conditions on the nutritional composition of quinoa seeds. PeerJ 2018, 14, e4442. [Google Scholar] [CrossRef] [PubMed]
- Lafitte, H.R.; Courtois, B. Interpreting cultivar X environment interactions for yield in upland rice: Assigning value to drought-adaptive traits. Crop Sci. 2002, 42, 1409–1420. [Google Scholar] [CrossRef]
- Subira, J.; Álvaro, F.; García del Moral, L.F.; Royo, C. Breeding effects on the cultivar×environment interaction of durum wheat yield, Eur. J. Agron. 2015, 68, 78–88. [Google Scholar] [CrossRef]
- Tonk, F.A.; Ilker, E.; Tosun, M. Evaluation of genotype X environment interactions in maize hybrids using GGE biplot analysis. Crop Breed. Appl. Biotechnol. 2011, 11, 1–9. [Google Scholar] [CrossRef]
- Bazile, D.; Pulvento, C.; Verniau, A.; Al-Nusairi, M.S.; Ba, D.; Breidy, J.; Hassan, L.; Mohammed, M.I.; Mambetov, O.; Otambekova, M.; et al. Worldwide evaluations of quinoa: Preliminary results from post international year of quinoa FAO projects in nine countries. Front. Plant Sci. 2016, 7, 850. [Google Scholar] [CrossRef]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef]
- García-Parra, M.; Plazas-Leguizamón, N. La quinua (Chenopodium quinoa Willd) en los sistemas de producción agraria. Prod.+ Limpia 2018, 13, 112–119. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Berlin, Germany, 2012. [Google Scholar]
- Granado-Rodríguez, S.; Vilariño-Rodríguez, S.; Maestro-Gaitán, I.; Matías, J.; Rodríguez, M.J.; Calvo, P.; Cruz, V.; Bolaños, L.; Reguera, M. Genotype-dependent variation of nutritional quality-related traits in quinoa seeds. Plants 2021, 10, 2128. [Google Scholar] [CrossRef]
- Thiam, E.; Allaoui, A.; Benlhabib, O. Quinoa productivity and stability evaluation through varietal and environmental interaction. Plants 2021, 10, 714. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Ohri, D. Chenopodium quinoa—An Indian perspective. Ind. Crops Prod. 2006, 23, 73–87. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Stanschewski, C.S.; Rey, E.; Gabriele, F.; Craine, E.B.; Wellman, G.; Melino, V.J.; Patiranage, D.S.R.; Johansen, K.; Schmöckel, S.M.; Bertero, D.; et al. Quinoa phenotyping methodologies: An international consensus. Plants 2021, 10, 1759. [Google Scholar] [CrossRef] [PubMed]
- García-Parra, M.A.; Stechauner-Rohringer, R.; Garcia-Molano, J.F.; Ortiz-Gonzalez, D. Analysis of the growth and morpho-physiological performance of three cultivars of colombian quinoa grown under a greenhouse. Rev. Cienc. Agrovet. 2020, 19, 73–83. [Google Scholar] [CrossRef]
- Manjarres-Hernández, E.; Morillo-Coronado, A.; Ojeda-perez, Z.; Cárdenas-Chaparro, A.; Arias-Moreno, D. Characterizacion of the yield components and selection of materials for breeding programs of quinoa (Chenopodium quinoa Willd.). Euphytica 2021, 217, 101. [Google Scholar] [CrossRef]
- Curti, R.N.; De la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, D. Multi-environmental evaluation for grain yield and its physiological determinants of quinoa genotypes across Northwest Argentina. Field Crop. Res. 2014, 166, 46–57. [Google Scholar] [CrossRef]
- Curti, R.N.; De la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, D. Adaptive responses of quinoa to diverse agro-ecological environments along an altitudinal gradient in North West Argentina. Field Crop. Res. 2016, 189, 10–18. [Google Scholar] [CrossRef]
- IGAC. Suelos de Boyacá no son tan Productivos Como se Piensa: IGAC. Pagina Oficial Instituto Geográfico Agustín Codazzi, 2016. Available online: https://noticias.igac.gov.co/en/contenido/suelos-de-boyaca-no-son-tan-productivos-como-se-piensa-igac (accessed on 6 June 2019).
- Sosa-Zuniga, V.; Brito, V.; Fuentes, F.; Steinfort, U. Phenological growth stages of quinoa (Chenopodium quinoa) based on the BBCH scale. Ann. Appl. Biol. 2017, 171, 117–124. [Google Scholar] [CrossRef]
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agric. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef]
- Alvar-Beltrán, J.; Dao, A.; Dalla Marta, A.; Saturnin, C.; Casini, P.; Sanou, J.; Orlandini, S. Effect of drought, nitrogen fertilization, temperature and photoperiodicity on quinoa plant growth and development in the Sahel. Agronomy 2019, 9, 607. [Google Scholar] [CrossRef] [Green Version]
- Dumschott, K.; Wuyts, N.; Alfaro, C.; Castillo, D.; Fiorani, F.; Zurita-Silva, A. Morphological and physiological traits associated with yield under reduced irrigation in chilean coastal lowland Quinoa. Plants 2022, 11, 323. [Google Scholar] [CrossRef] [PubMed]
- García-Parra, M.; García-Molano, J. Deaquiz-Oyola, Y. Physiological performance of quinoa (Chenopodium quinoa Willd.) under agricultural climatic conditions in Boyaca, Colombia. Agron. Colomb. 2019, 37, 160–168. [Google Scholar] [CrossRef]
- Lesjak, J.; Calderini, D. Increased night temperature negatively affects grain yield, biomass and grain number in chilean quinoa. Front. Plant Sci. 2017, 8, 352. [Google Scholar] [CrossRef] [PubMed]
- Akter, N.; Islam, M.R. Heat stress effects and management in wheat. A. review. Agron. Sustain. Dev. 2017, 37, 37. [Google Scholar] [CrossRef]
- Martínez, D.E.; Costa, M.L.; Gomez, F.M.; Otegui, M.S.; Guiamet, J. Senescence-associated vacuoles’ are involved in the degradation of chloroplast proteins in tobacco leaves. Plant J. 2008, 56, 196–206. [Google Scholar] [CrossRef]
- Bertero, H.D.; Ruiz, R.A. Determination of seed number in sea level quinoa (Chenopodium quinoa Willd.) cultivars. Eur. J. Agron. 2008, 28, 186–194. [Google Scholar] [CrossRef]
- Bascuñán-Godoy, L.; Sanhueza, C.; Hernández, C.E.; Cifuentes, L.; Pinto, K.; Álvarez, R.; González-Teuber, M.; Bravo, L.A. Nitrogen supply affects photosynthesis and photoprotective attributes during drought-induced senescence in quinoa. Front. Plant Sci. 2018, 9, 994. [Google Scholar] [CrossRef]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Phenotyping of wheat cultivars for heat tolerance using chlorophyll a fluorescence. Funct. Plant Biol. 2012, 39, 936–947. [Google Scholar] [CrossRef]
- Eustis, A.; Murphy, K.M.; Barrios-Masias, F.H. Leaf gas exchange performance of ten quinoa genotypes under a simulated heat wave. Plants 2020, 9, 81. [Google Scholar] [CrossRef]
- Jiménez-Suancha, S.; Alvarado, O.; Balaguera-López, H. Fluorescencia como indicador de estrés en Helianthus annuus L. Una revisión. Rev. Colomb. Cienc. Hortícolas 2015, 9, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Parra, M.; Stechauner-Rohringer, R.; Roa-Acosta, D.; Ortiz-González, D.; Ramirez-Correa, J.; Plazas-Leguizamón, N.; Colmenares-Cruz, A. Chlorophyll fluorescence and its relationship with physiological stress in Chenopodium quinoa Willd. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1742–1755. [Google Scholar] [CrossRef]
- Panda, D.; Sarkar, R.K. Natural leaf senescence: Probed by chlorophyll fluorescence, CO2 photosynthetic rate and antioxidant enzyme activities during grain filling in different rice cultivars. Physiol. Mol. Biol. Plants 2013, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Issa-Ali, O.; Fghire, R.; Anaya, F.; Benlhabib, O.; Wahbi, S. Physiological and morphological responses of two quinoa cultivars (Chenopodium quinoa Willd.) to Drought Stress. Gesunde Pflanz. 2019, 71, 123–133. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Z.; Li, S.; Lü, X.; Wang, X.; Han, X. Changes in specific leaf area of dominant plants in temperate grasslands along a 2500-km transect in northern China. Sci. Rep. 2017, 7, 10780. [Google Scholar] [CrossRef]
- Stuerz, S.; Asch, F. Responses of rice growth to day and night temperature and relative air humidity-dry matter, leaf area, and partitioning. Plants 2019, 8, 521. [Google Scholar] [CrossRef]
- Präger, A.; Munz, S.; Nkebiwe, P.; Mast, B.; Graeff-Hönninger, S. Yield and quality characteristics of different quinoa (Chenopodium quinoa Willd.) cultivars grown under field conditions in southwestern Germany. Agronomy 2018, 8, 197. [Google Scholar] [CrossRef]
- Yan, H.; Shah, S.S.; Zhao, W.; Liu, F. Variations in water relations, stomatal characteristics, and plant growth between quinoa and pea under salt-stress conditions. Pak. J. Bot. 2020, 52, 1–7. [Google Scholar] [CrossRef]
- Bunce, J.A. Variation in yield responses to elevated CO2 and a brief high temperature treatment in quinoa. Plants 2017, 6, 26. [Google Scholar] [CrossRef]
- Choukr-Allah, R.; Rao, N.K.; Hirich, A.; Shahid, M.; Alshankiti, A.; Toderich, K.; Gill, S.; Butt, K.U.R. Quinoa for marginal environments: Toward future food and nutritional security in MENA and central Asia regions. Front. Plant Sci. 2016, 7, 346. [Google Scholar] [CrossRef]
- Gonzalez, J.A.; Konishi, Y.; Bruno, M.; Valoy, M.; Prado, F.E. Interrelationships among seed yield, total protein and amino acid composition of ten quinoa (Chenopodium quinoa) cultivars from two different agroecological regions. J. Sci. Food Agric. 2012, 92, 1222–1229. [Google Scholar] [CrossRef]
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. Front. Plant Sci. 2018, 9, 1889. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, S.; Deng, X.; Zhang, Z.; Yin, L. Comprehensive evaluation of physiological traits under nitrogen stress and participation of linolenic acid in nitrogen-deficiency response in wheat seedlings. BMC Plant Biol. 2020, 20, 501. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, J.; Chae, M.; Kong, M.; Lee, C.; Kang, S.; Kim, Y. Growth-inhibition patterns and transfer-factor profiles in arsenic-stressed rice (Oryza sativa L.). Environ. Monit. Assess. 2017, 189, 2017. [Google Scholar] [CrossRef] [PubMed]
- Tahjib-Ul-Arif, T.; Siddiqui, N.; Sohag, A.; Sakil, A.; Rahman, M.; Polash, M.; Mostafa, M.; Tran, L. Salicylic acid-mediated enhancement of photosynthesis attributes and antioxidant capacity contributes to yield improvement of maize plants under salt stress. J. Plant Growth Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- Sanabria, K.M.; Lazo, H.O. Aclimatación a la alta temperatura y tolerancia al calor (TL 50) en 6 variedades de Chenopodium quinoa. Rev. Peru. Biol. 2018, 25, 147–152. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Geographical Position | Climate (Caldas Lang) | Altitude (m) | Soil Type * | T (°C) | Rainfall (mm) | Planting Day | |
|---|---|---|---|---|---|---|---|---|
| Latitude | Longitude | |||||||
| Oicatá | 5°36′ N | 73°10′ W | Cold | 2648 | Association Vertic Haplustalf—Andic Dystrustepts | 12.1 | 741 | 8 April 2020 |
| Moniquirá | 5°52′ N | 73°33′ W | Temperate | 1707 | Complex Chromic Hapluderts—Typic Dystrudepts | 18.1 | 1305.9 | 14 April 2020 |
| Pauna | 5°39′ N | 74°04′ W | Warm | 698 | Typic Eustrudepts—Typic Udorthents—Humic Dystrudepts | 23.1 | 1984 | 18 April 2020 |
| Altitudinal Gradient | Cultivar | First Pair of Leaves Visible (11) | One Side Shoot Visible (21) | Inflorescence Present (50) | Beginning of Anthesis (60) | Milky Grain (81) | Thick Grain (85) | Plant Dead and Dry (97) |
|---|---|---|---|---|---|---|---|---|
| Cold | Blanca Real | 13.66 ± 1.52 abcde | 21.66 ± 1.15 f | 50 ± 2 abcd | 87 ± 2 ab | 97.66 ± 2.51 abc | 111.33 ± 2.88 abc | 124.33 ± 4.04 bcd |
| Pasankalla | 10.66 ± 0.57 bcdef | 32 ± 2 ab | 48.66 ± 1.15 abcd | 74.66 ± 1.52 fgh | 101.66 ± 3.21 ab | 110.66 ± 7.50 abc | 122.33 ± 2.08 bcde | |
| Nariño | 14.66 ± 1.15 abc | 23.66 ± 0.57 ef | 42.33 ± 2.51 fg | 75.33 ± 2.51 fgh | 86.33 ± 3.05 abc | 99 ± 7.81 bc | 111.33 ± 3.51 ef | |
| Soracá | 13.33 ± 1.52 abcde | 24.66 ± 2.51 cdef | 51.33 ± 1.52 abcd | 91.33 ± 1.52 a | 108.33 ± 5.50 a | 124.33 ± 12.89 a | 136.66 ± 1.52 a | |
| Titicaca | 14.33 ± 0.57 abcd | 33.66 ± 1.52 a | 51.66 ± 1.52 abc | 76 ± 1 efg | 87.33 ± 3.78 c | 100 ± 5 bc | 114 ± 5.29 def | |
| Nueva | 11 ± 1 bcdef | 23.33 ± 2.08 ef | 51 ± 1.15 abcd | 76.33 ± 1.52 efg | 86.66 ± 4.16 abc | 113.66 ± 5.13 ab | 111.33 ± 2.08 bcde | |
| Puno | 9.66 ± 1.52 cdef | 22.33 ± 1.52 ef | 47 ± 1.73 cdef | 77.66 ± 1.52 def | 89 ± 5.29 bc | 106 ± 5.19 bc | 116.33 ± 4.72 def | |
| Temperate | Blanca real | 15.66 ± 2.08 ab | 24 ± 1 ef | 50 ± 2 abcd | 85 ± 1 bc | 96 ± 5.29 abc | 107.66 ± 2.51 abc | 118.33 ± 1.52 cdef |
| Pasankalla | 16.33 ± 1.52 a | 31.33 ± 1.15 ab | 49.66 ± 1.52 abcd | 73 ± 1 fgh | 96.33 ± 5.50 abc | 109 ± 3.60 abc | 129.66 ± 1.53 abc | |
| Nariño | 13.33 ± 30.5 abcde | 24.33 ± 1.52 def | 43.33 ± 1.52 efg | 74 ± 1 fgh | 86.33 ± 5.50 c | 100.66 ± 6.65 bc | 113 ± 4.35 def | |
| Soracá | 14.33 ± 1.15 abcde | 28.66 ± 1.52 bcd | 51.33 ± 1.52 abcd | 83 ± 2.64 bc | 95 ± 5 abc | 112.66 ± 6.42 abc | 132 ± 2 ab | |
| Titicaca | 10.33 ± 1.52 cdef | 35.33 ± 1.52 a | 53 ± 1 ab | 75 ± 1 fgh | 87 ± 4.35 c | 101.33 ± 5.50 bc | 113.66 ± 5.05 def | |
| Nueva | 10 ± 1 cdef | 25 ± 1 cdef | 53.33 ± 1.52 a | 72.66 ± 1.52 fgh | 90 ± 2.64 bc | 104 ± 5.29 bc | 119.66 ± 1.52 cdef | |
| Puno | 9 ± 1 ef | 23.33 ± 1.15 ef | 48 ± 1 bcde | 76.33 ± 1.52 efg | 88 ± 2.64 bc | 102 ± 5.29 bc | 117.66 ± 2.51 def | |
| Warm | Blanca real | 13.66 ± 2.08 abcde | 21.66 ± 1.15 f | 48.33 ± 1.52 abcde | 82.66 ± 1.52 bcd | 97 ± 3.60 abc | 107.33 ± 4.50 abc | 118.33 ± 1.52 cdef |
| Pasankalla | 13.33 ± 1.52 abcde | 29 ± 1.73 bc | 50.33 ± 3.5 abcd | 70.66 ± 0.57 h | 93 ± 7 bc | 111 ± 3.60 abc | 118.66 ± 3.21 cdef | |
| Nariño | 11.66 ± 2.51 abcdef | 22.66 ± 1.15 ef | 41 ± 1 g | 71.66 ± 1.52 gh | 85 ± 4.35 c | 95.66 ± 4.93 c | 111.66 ± 7.63 ef | |
| Soracá | 12 ± 1 abcdef | 26.66 ± 1.52 cde | 49.33 ± 1.52 abcd | 81 ± 2.64 cde | 91 ± 4.58 bc | 109 ± 4.58 abc | 121 ± 3.60 bcdef | |
| Titicaca | 9.66 ± 2.08 cdef | 34 ± 1 a | 51.33 ± 1.15 abcd | 72.66 ± 1.52 fgh | 86.66 ± 4.16 c | 101 ± 2.64 bc | 111 ± 3.60 ef | |
| Nueva | 9.33 ± 1.52 def | 23 ± 1 ef | 51 ± 1 abcd | 71.33 ± 1.52 gh | 90.33 ± 4.35 bc | 102 ± 5.29 bc | 113 ± 4.35 def | |
| Puno | 8 ± 2 f | 21.33 ± 1.52 f | 46.33 ± 1.52 def | 73.33 ± 1.53 fgh | 85.33 ± 4.72 c | 96 ± 5.29 bc | 110 ± 4.58 f | |
| Altitudinal | ** | *** | * | *** | *** | ** | *** | |
| Cultivar | *** | *** | *** | *** | *** | *** | *** | |
| A/C | ** | NS | NS | * | NS | NS | * |
| Altitudinal Gradient | Cultivar | First Pair of Leaves Visible (11) | First Pair of Leaves Visible (21) | First Pair of Leaves Visible (60) | Fruit Set (70) |
|---|---|---|---|---|---|
| Cold | Blanca Real | 28 ± 2.64 bcd | 40 ± 2 bcde | 46.66 ± 1.57 cdef | 40 ± 2 ef |
| Pasankalla | 28.33 ± 3.78 bcd | 42 ± 1.73 bcd | 57.66 ± 2.51 a | 57.66 ± 2.51 a | |
| Nariño | 25 ± 3 cde | 37 ± 1.73 de | 50 ± 2 abcde | 32 ± 2 g | |
| Soracá | 20.66 ± 1.15 e | 34.66 ± 0.57 e | 56.66 ± 3.51 ab | 54.66 ± 5.03 ab | |
| Titicaca | 22.33 ± 2.08 de | 39.66 ± 1.52 bcde | 42.33 ± 2.51 ef | 31.66 ± 2.88 g | |
| Nueva | 22.66 ± 2.59 de | 34.66 ± 2.51 e | 48 ± 3 cdef | 40.33 ± 1.52 ef | |
| Puno | 21 ± 1 e | 40.33 ± 0.57 bcde | 48.33 ± 2.08 cdef | 36 ± 1.73 fg | |
| Temperate | Blanca Real | 35 ± 1 a | 44.66 ± 1.52 abc | 45 ± 2 ef | 37.66 ± 2.51 fg |
| Pasankalla | 25 ± 2.64 cde | 46 ± 5.29 ab | 54.33 ± 4.04 abc | 52.33 ± 2.51 abc | |
| Nariño | 26 ± 1.73 bcde | 39.66 ± 2.08 bcde | 47.66 ± 2.51 cdef | 31 ± 1 g | |
| Soracá | 26.66 ± 1.52 bcde | 39.33 ± 1.52 bcde | 53 ± 3.60 abcd | 50.33 ± 1.52 bcd | |
| Titicaca | 28 ± 2 bcd | 41.33 ± 3.51 bcde | 42.33 ± 2.51 ef | 31 ± 1 g | |
| Nueva | 26 ± 1.73 bcde | 39.66 ± 2.08 bcde | 46.33 ± 1.52 def | 37 ± 2 fg | |
| Puno | 30 ± 2 abc | 41.33 ± 1.52 bcde | 48.33 ± 2.51 cdef | 34 ± 2 fg | |
| Warm | Blanca Real | 27 ± 2 bcde | 41.33 ± 1.57 bcde | 43.33 ± 2.08 ef | 32.66 ± 2.08 g |
| Pasankalla | 32.33 ± 2.51 ab | 51 ± 1 a | 53 ± 2 abcd | 45 ± 3 de | |
| Nariño | 27.66 ± 2.08 bcd | 41 ± 1.73 bcde | 46.66 ± 1.52 cdef | 31 ± 1 g | |
| Soracá | 28 ± 1 bcd | 40 ± 2 bcde | 49.66 ± 2.51 bcdef | 46.66 ± 2.08 cde | |
| Titicaca | 26.66 ± 1.52 bcde | 38 ± 2 cde | 42 ± 2 f | 30.66 ± 1.15 g | |
| Nueva | 28.33 ± 1.15 bcd | 38.66 ± 2.51 cde | 44.66 ± 2.51 ef | 32.33 ± 2.51 g | |
| Puno | 28.66 ± 1.15 abcd | 41.33 ± 2.08 bcde | 48 ± 2 cdef | 33.66 ± 3.21 fg | |
| Altitudinal | *** | *** | *** | *** | |
| Cultivar | *** | *** | *** | *** | |
| A/C | *** | * | NS | ** |
| Altitudinal Gradient | Cultivar | First Pair of Leaves Visible (11) | First Pair of Leaves Visible (21) | First Pair of Leaves Visible (60) | Fruit Set (70) |
|---|---|---|---|---|---|
| Cold | Blanca Real | 0.81 ±0.010 abcd | 0.77 ± 0.010 abcd | 0.76 ± 0.011 ab | 0.68 ± 0.015 ab |
| Pasankalla | 0.82 ± 0.020 abc | 0.77 ± 0.020 abc | 0.76 ± 0.015 ab | 0.69 ± 0.010 a | |
| Nariño | 0.83 ± 0.010 a | 0.78 ± 0.015 a | 0.77 ± 0.010 a | 0.66 ± 0.011 ab | |
| Soracá | 0.82 ± 0.010 abc | 0.76 ± 0.010 abcde | 0.75 ± 0.020 abcd | 0.68 ± 0.015 ab | |
| Titicaca | 0.80 ± 0.005 abcde | 0.77 ± 0.010 abcd | 0.76 ± 0.010 abc | 0.67 ± 0.015 ab | |
| Nueva | 0.81 ± 0.010 abcd | 0.75 ± 0.005 abcdef | 0.78 ± 0.010 a | 0.67 ± 0.015 ab | |
| Puno | 0.82 ± 0.020 ab | 0.78 ± 0.005 ab | 0.77 ± 0.020 a | 0.69 ± 0.010 a | |
| Temperate | Blanca Real | 0.79 ± 0.015 abcdef | 0.74 ± 0.011 abcdef | 0.72 ± 0.015 bcde | 0.65 ± 0.020 ab |
| Pasankalla | 0.79 ± 0.010 abcdef | 0.72 ± 0.015 def | 0.71 ± 0.010 e | 0.64 ± 0.005 b | |
| Nariño | 0.79 ± 0.005 abcdef | 0.74 ± 0.017 abcdef | 0.72 ± 0.010 cde | 0.66 ± 0.015 ab | |
| Soracá | 0.76 ± 0.010 f | 0.74 ± 0.010 abcdef | 0.72 ± 0.015 bcde | 0.66 ± 0.005 ab | |
| Titicaca | 0.77 ± 0.015 def | 0.73 ± 0.025 cdef | 0.7 ± 0.015 e | 0.64 ± 0.005 b | |
| Nueva | 0.76 ± 0.015 f | 0.73 ± 0.025 bcdef | 0.71 ± 0.010 e | 0.65 ± 0.015 ab | |
| Puno | 0.78 ± 0.005 bcdef | 0.72 ± 0.017 ef | 0.7 ± 0.011 e | 0.66 ± 0.011 ab | |
| Warm | Blanca Real | 0.76 ± 0.015 ef | 0.71 ± 0.015 ef | 0.7 ± 0.010 e | 0.65 ± 0.025 ab |
| Pasankalla | 0.76 ± 0.011 ef | 0.71 ± 0.015 ef | 0.71 ± 0.020 de | 0.65 ± 0.020 ab | |
| Nariño | 0.78 ± 0.005 bcdef | 0.72 ± 0.010 ef | 0.71 ± 0.010 e | 0.64 ± 0.005 b | |
| Soracá | 0.78 ± 0.010 cdef | 0.73 ± 0.017 cdef | 0.71 ± 0.010 bcde | 0.64 ± 0.005 b | |
| Titicaca | 0.78 ± 0.020 bcdef | 0.71 ± 0.017 f | 0.71 ± 0.015 e | 0.65 ± 0.011 ab | |
| Nueva | 0.79 ± 0.005 abcdef | 0.71 ± 0.015 ef | 0.69 ± 0.011 e | 0.66 ± 0.020 ab | |
| Puno | 0.78 ± 0.015 bcdef | 0.71 ± 0.020 ef | 0.7 ± 0.010 e | 0.67 ± 0.005 ab | |
| Altitudinal | *** | *** | *** | *** | |
| Cultivar | NS | NS | NS | NS | |
| A/C | * | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Parra, M.; Roa-Acosta, D.; Bravo-Gómez, J.E. Effect of the Altitude Gradient on the Physiological Performance of Quinoa in the Central Region of Colombia. Agronomy 2022, 12, 2112. https://doi.org/10.3390/agronomy12092112
García-Parra M, Roa-Acosta D, Bravo-Gómez JE. Effect of the Altitude Gradient on the Physiological Performance of Quinoa in the Central Region of Colombia. Agronomy. 2022; 12(9):2112. https://doi.org/10.3390/agronomy12092112
Chicago/Turabian StyleGarcía-Parra, Miguel, Diego Roa-Acosta, and Jesús Eduardo Bravo-Gómez. 2022. "Effect of the Altitude Gradient on the Physiological Performance of Quinoa in the Central Region of Colombia" Agronomy 12, no. 9: 2112. https://doi.org/10.3390/agronomy12092112