
Monitoring the Bud Mite Pest in a Hazelnut Orchard of Central Italy: Do Plant Height and Irrigation Influence the Infestation Level?


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Monitoring
2.2.1. Infestation over the Years and Plant Irrigation
2.2.2. Infestation over Plant Height Bands
2.2.3. Calculation of the Overall Infestation Threshold over the Years and Irrigation
2.3. Statistical Analysis
2.3.1. Differences in Infestation over the Years and Plant Irrigation
2.3.2. Differences in Infestation over Plant Height Bands
2.3.3. Software for Data Analysis
3. Results
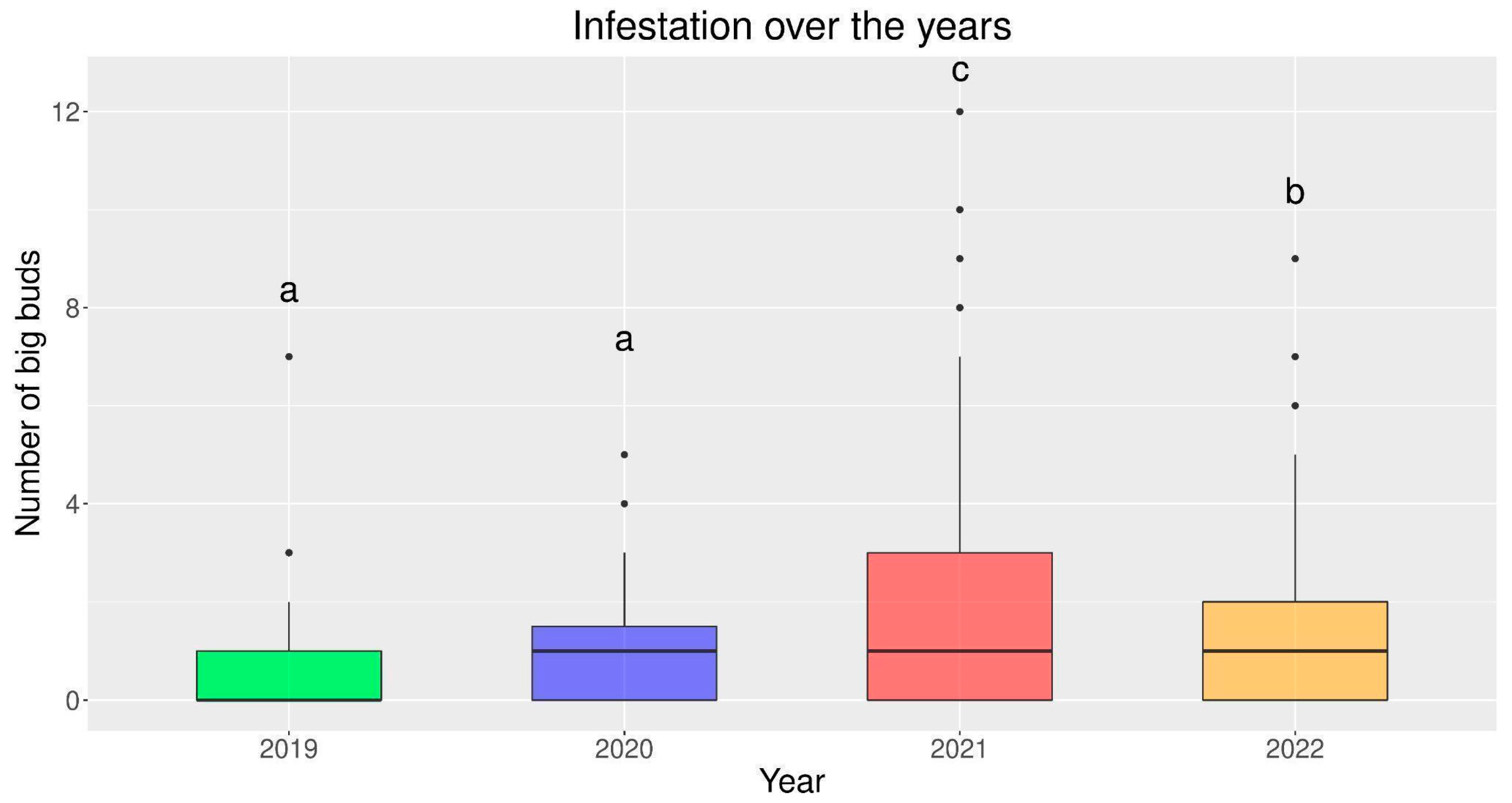
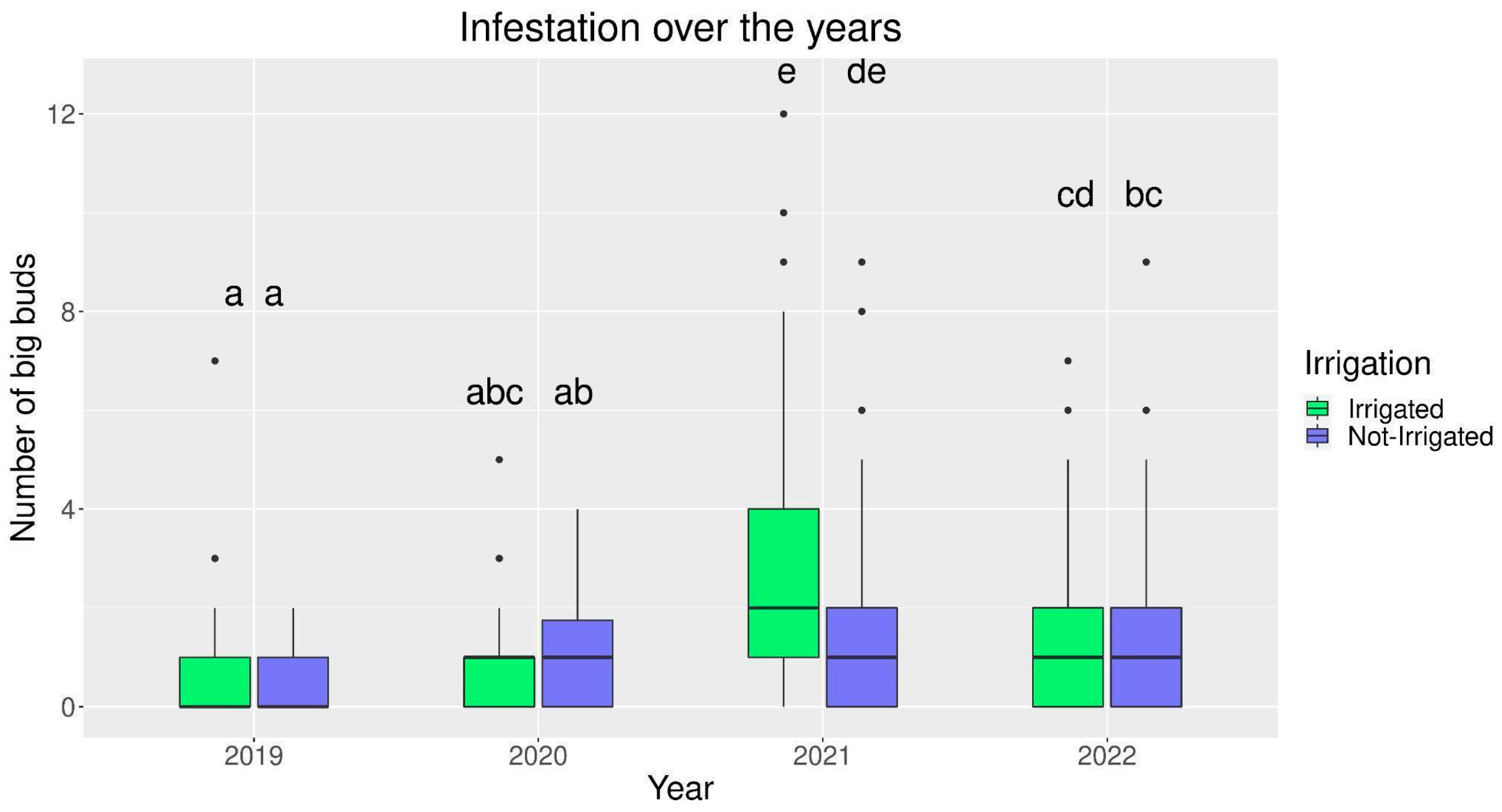
3.1. Infestation over the Years and among Different Plant Irrigation Systems
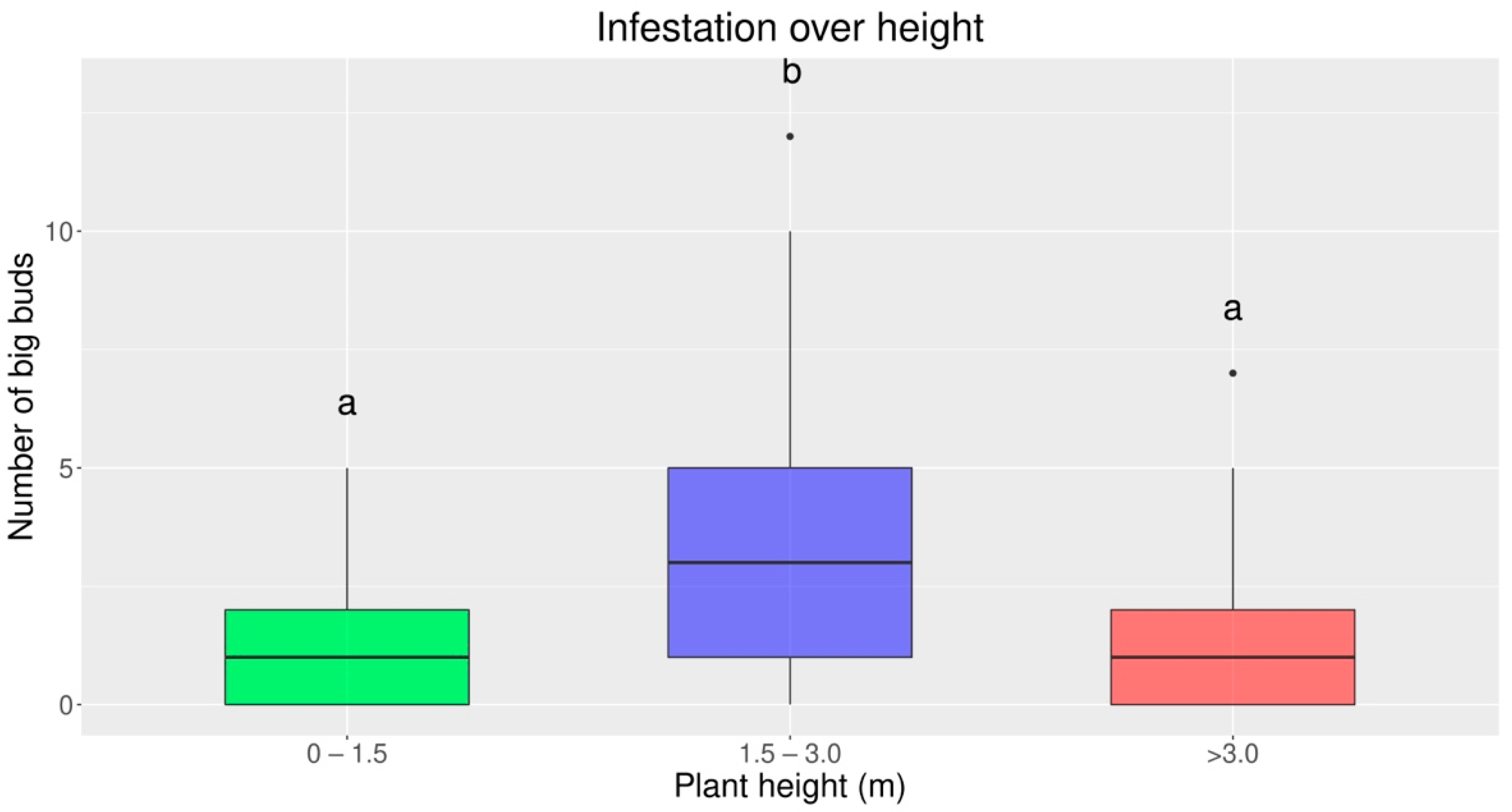
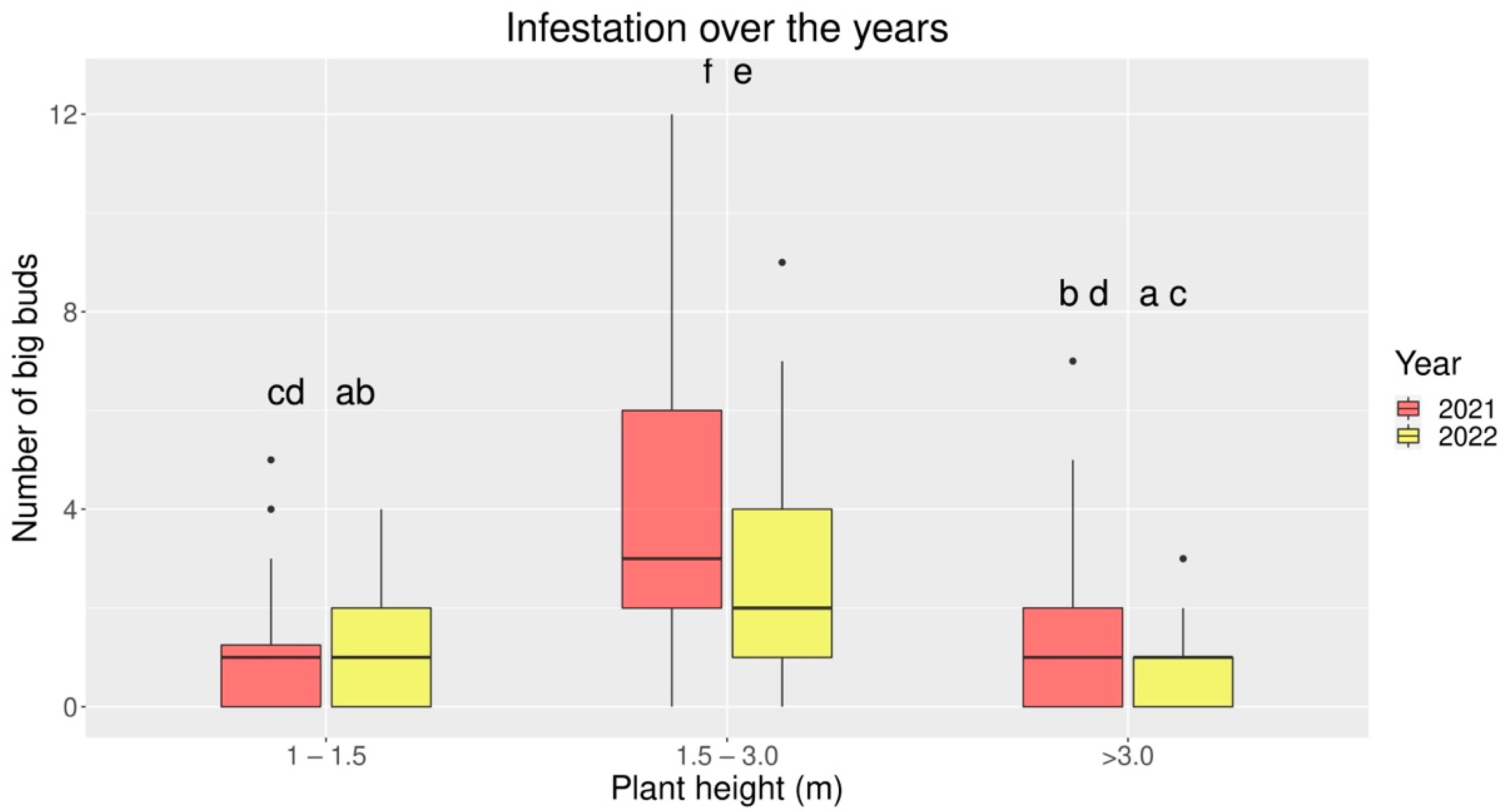
3.2. Infestation over Plant Height Bands
3.3. Infestation Ratio (Big Buds on Non-Infested Buds) over the Years and Plant Irrigation
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ISTAT. Available online: http://agri.istat.it/jsp/dawinci.jsp?q=plC190000010000011000&an=2017&ig=1&ct=270&id=15A%7C21A%7C30A (accessed on 17 July 2022).
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data (accessed on 17 July 2022).
- AliNiazee, M.T. Ecology and management of hazelnut pests. Annu. Rev. Entomol. 1998, 43, 395–419. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- Turan, A.; Erdoğan, V. Spread and damage of citrus longhorned beetle [Anoplophora chinensis (Forster, 1771) (Coleoptera: Cerambycidae)] to hazelnut orchards in Turkey. Turk. J. Agric.-Food Sci. Technol. 2022, 10, 531–535. [Google Scholar] [CrossRef]
- Mazzaglia, A.; Drais, M.I.; Turco, S.; Silvestri, C.; Cristofori, V.; Aymami, A.; Casadó, V.; Rovira, M. First report of Erysiphe corylacearum causing powdery mildew on Corylus avellana in Spain. New Dis. Rep. 2021, 44, 2–4. [Google Scholar] [CrossRef]
- Mezzalama, M.; Guarnaccia, V.; Martano, G.; Spadaro, D. Presence of powdery mildew caused by Erysiphe corylacearum on hazelnut (Corylus avellana) in Italy. Plant Dis. 2021, 105, 1565. [Google Scholar] [CrossRef]
- Sezer, A.; Dolar, F.S.; Lucas, S.J.; Köse, Ç.; Gümüş, E. First report of the recently introduced, destructive powdery mildew Erysiphe corylacearum on hazelnut in Turkey. Phytoparasitica 2017, 45, 577–581. [Google Scholar] [CrossRef]
- Turco, S.; Zuppante, L.; Drais, M.I.; Mazzaglia, A. Dressing like a pathogen: Comparative analysis of different Pseudomonas genomospecies wearing different features to infect Corylus avellana. J. Phytopathol. 2022, 170, 504–516. [Google Scholar] [CrossRef]
- Memoli, A.; Albanese, D.; Esti, M.; Lombardelli, C.; Crescitelli, A.; di Matteo, M.; Benucci, I. Effect of bug damage and mold contamination on fatty acids and sterols of hazelnut oil. Eur. Food Res. Technol. 2017, 243, 651–658. [Google Scholar] [CrossRef]
- Rossini, L.; Contarini, M.; Speranza, S. A novel version of the Von Foerster equation to describe poikilothermic organisms including physiological age and reproduction rate. Ric. Mat. 2021, 70, 489–503. [Google Scholar] [CrossRef]
- Rossini, L.; Contarini, M.; Giarruzzo, F.; Assennato, M.; Speranza, S. Modelling Drosophila suzukii adult male populations: A physiologically based approach with validation. Insects 2020, 11, 751. [Google Scholar] [CrossRef]
- Özman, S.K.; Toros, S. Life cycles of Phytoptus avellanae Nal. and Cecidiopyopsis vermiformis Nal. (Eriphyoidea: Acarina). Acta Hortic. 1997, 445, 493–502. [Google Scholar] [CrossRef]
- Ozman-Sullivan, S.K. Do the contradictory life cycles of the hazelnut big bud mite, Phytoptus avellanae (Nal.) (Acari: Phytoptidae) imply two species? In Proceedings of the Abstract book, XIV International Congress of Acarology, Kyoto, Japan, 13–18 July 2014; p. 62. [Google Scholar]
- Cvrković, T.; Chetverikov, P.; Vidović, B.; Petanović, R. Cryptic speciation within Phytoptus avellanae s.l. (Eriophyoidea: Phytoptidae) revealed by molecular data and observations on molting tegonotus-like nymphs. Exp. Appl. Acarol. 2016, 68, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Vacante, V. The Handbook of Mites of Economic Plants: Identification, Bio-Ecology and Control; CABI: Wallingford, UK, 2015. [Google Scholar]
- Ozman-Sullivan, S.K.; Akça, I. Efficiency of pesticides against big bud mites [Phytoptus avellanae Nal. and Cecidophyopsis vermiformis Nal. (Acarina: Eriophyoidea)] on hazelnut. Acta Hortic. 2005, 686, 393–400. [Google Scholar] [CrossRef]
- Stamenkovic, S.; Milenkovic, S.; Pesic, M.; Mitrovic, M. Population dynamics, harmfulness and control of Phytoptus avellanae (Nalepa) in Western Serbia. Acta Hortic. 1997, 445, 521–526. [Google Scholar] [CrossRef]
- Westphal, E.; Manson, D.C.M. 1.4.6 Feeding effects on host plants: Gall formation and other distortions. In World Crop Pests; Elsevier: Amsterdam, The Netherlands, 1996; pp. 231–242. [Google Scholar]
- Viggiani, G.; Bianco, M. Osservazioni ed esperienze per una lotta chimica razionale contro Phytoptus avellanae Nalepa (Acarina: Eriophyidea). Boll. Lab. Entomol. Agrar. 1974, 31, 3–26. [Google Scholar]
- Rossini, L.; Bono Rosselló, N.; Speranza, S.; Garone, E. A general ODE-based model to describe the physiological age structure of ectotherms: Description and application to Drosophila suzukii. Ecol. Model. 2021, 456, 109673. [Google Scholar] [CrossRef]
- Lippi, M.; Bonucci, N.; Carpio, R.F.; Contarini, M.; Speranza, S.; Gasparri, A. A YOLO-based pest detection system for precision agriculture. In Proceedings of the 2021 29th Mediterranean Conference on Control and Automation (MED), Bari, Italy, 22–25 June 2021; pp. 342–347. [Google Scholar]
- Lippi, M.; Carpio, R.F.; Contarini, M.; Speranza, S.; Gasparri, A. A data-driven monitoring system for the early pest detection in the precision agriculture of hazelnut orchards. In Proceedings of the 7th IFAC Conference on Sensing, Control and Automation Technologies for Agriculture, Munich, Germany, 14–16 September 2022. [Google Scholar]
- Di Sora, N.; Rossini, L.; Contarini, M.; Chiarot, E.; Speranza, S. Endotherapic treatment to control Toumeyella parvicornis Cockerell infestations on Pinus pinea L. Pest Manag. Sci. 2022, 78, 2443–2448. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Abramishvili, T.; Gaganidze, D.; Ozman-Sullivan, S.K.; Abashidze, E. Phylogenetic analysis of hazelnut big bud mite—Phytoptus avellanae Nal. in the black sea region of Georgia. Bull. Georgian Natl. Acad. Sci. 2018, 12, 113–118. [Google Scholar]
- Kereselidze, M.; Goderdzishvili, G.; Khidesheli, Z. Monitoring damage caused by harmful pests in hazelnut orchards in Samegrelo Region, Georgia. Acta Hortic. 2018, 1226, 373–378. [Google Scholar] [CrossRef]
- Gantner, M. Susceptibility of large-fruited hazel cultivars grown in Poland to major pest and their crop productivity. Acta Hortic. 2005, 686, 377–384. [Google Scholar] [CrossRef]
- Mehlenbacher, S.A. Genetic improvement of hazelnut. Acta Hortic. 1994, 351, 23–38. [Google Scholar] [CrossRef]
- Tuncer, C. Arthropod management in organic hazelnut growing. Acta Hortic. 2009, 845, 571–578. [Google Scholar] [CrossRef]
- Vergni, L.; Todisco, F. Spatio-temporal variability of precipitation, temperature and agricultural drought indices in Central Italy. Agric. For. Meteorol. 2011, 151, 301–313. [Google Scholar] [CrossRef]
- González-Zamora, J.E.; Alonso-López, M.T.; Gómez-Regife, Y.; Ruiz-Muñoz, S. Decreased water use in a super-intensive olive orchard mediates arthropod populations and pest damage. Agronomy 2021, 11, 1337. [Google Scholar] [CrossRef]
- Flaherty, D.L.; Huffaker, C.B. Biological control of Pacific mites and Willamette mites in San Joaquin Valley vineyards: I. Role of Metaseiulus occidentalis. Hilgardia 1970, 40, 267–308. [Google Scholar] [CrossRef] [Green Version]
- Kinn, D.N.; Joos, J.L.; Doutt, R.L. Influence of overhead sprinkler systems on spider mite populations in north coast vineyards of California. Environ. Entomol. 1972, 1, 795–796. [Google Scholar] [CrossRef]
- Opit, G.P.; Fitch, G.K.; Margolies, D.C.; Nechols, J.R.; Williams, K.A. Overhead and drip-tube irrigation affect twospotted spider mites and their biological control by a predatory mite on impatiens. HortScience 2006, 41, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Chandler, L.D.; Archer, T.L.; Ward, C.R.; Lyle, W.M. Influences of irrigation practices on spider mite. Densities on field corn. Environ. Entomol. 1979, 8, 196–201. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Arthropod vertical stratification in temperate deciduous forests: Implications for conservation-oriented management. For. Ecol. Manag. 2011, 261, 1479–1489. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Bruin, J. 1.5.3. Evolutionary ecology: Life history patterns, food plant choice and dispersal. In World Crop Pests; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar]
- Valenzano, D.; Bari, G.; Valeria, M.; de Lillo, E. Off-host survival of Eriophyoidea and remarks on their dispersal modes. Exp. Appl. Acarol. 2019, 79, 21–33. [Google Scholar] [CrossRef]
- Duffner, K.; Schruft, G.; Guggenheim, R. Passive dispersal of the grape rust mite Calepitrimerus vitis Nalepa 1905 (Acari, Eriophyoidea) in vineyards. Anz. Fur Schent Glyphadlingskunde 2001, 74, 1–6. [Google Scholar] [CrossRef]
- Galvão, A.S.; Melo, J.W.S.; Monteiro, V.B.; Lima, D.B.; de Moraes, G.J.; Gondim, M.G.C. Dispersal strategies of Aceria guerreronis (Acari: Eriophyidae), a coconut pest. Exp. Appl. Acarol. 2012, 57, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rossini, L.; Bono Rosselló, N.; Contarini, M.; Speranza, S.; Garone, E. Modelling ectotherms’ populations considering physiological age structure and spatial motion: A novel approach. Ecol. Inform. 2022, 70, 101703. [Google Scholar] [CrossRef]
- Calvet, É.C.; Lima, D.B.; Melo, J.W.S.; Gondim, M.G.C., Jr. Host plant discrimination through mobility parameters by eriophyoid mites. Syst. Appl. Acarol. 2020, 25, 1541–1551. [Google Scholar] [CrossRef]
- Rossello, N.B.; Garone, E. Carrier-vehicle system for delivery in city environments. IFAC-PapersOnLine 2020, 53, 15253–15258. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year of Survey | Infestation Index (R) on Irrigated Plants | Infestation Index on Not-Irrigated Plants |
|---|---|---|
| 2019 | | |
| 2020 | | |
| 2021 | | |
| 2022 | | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contarini, M.; Rossini, L.; Di Sora, N.; de Lillo, E.; Speranza, S. Monitoring the Bud Mite Pest in a Hazelnut Orchard of Central Italy: Do Plant Height and Irrigation Influence the Infestation Level? Agronomy 2022, 12, 1982. https://doi.org/10.3390/agronomy12081982
Contarini M, Rossini L, Di Sora N, de Lillo E, Speranza S. Monitoring the Bud Mite Pest in a Hazelnut Orchard of Central Italy: Do Plant Height and Irrigation Influence the Infestation Level? Agronomy. 2022; 12(8):1982. https://doi.org/10.3390/agronomy12081982
Chicago/Turabian StyleContarini, Mario, Luca Rossini, Nicolò Di Sora, Enrico de Lillo, and Stefano Speranza. 2022. "Monitoring the Bud Mite Pest in a Hazelnut Orchard of Central Italy: Do Plant Height and Irrigation Influence the Infestation Level?" Agronomy 12, no. 8: 1982. https://doi.org/10.3390/agronomy12081982