
An R2R3-Type Transcription Factor OsMYBAS1 Regulates Seed Germination under Artificial Accelerated Aging in Transgenic Rice (Oryza sativa L.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Treatment Methods
2.2. Artificial Accelerated Aging Test
2.3. Standard Germination Test
2.4. Determination of Enzyme Activity and the Malondialdehyde and Proline Content
2.5. RNA Extraction, cDNA Library Preparation, and Transcriptome Sequencing
2.6. Differential Expression Analysis
2.7. GO and KEGG Enrichment Analysis
2.8. Gene Set Enrichment Analysis (GSEA)
2.9. qRT-PCR Detection
2.10. Statistical Analysis
3. Results
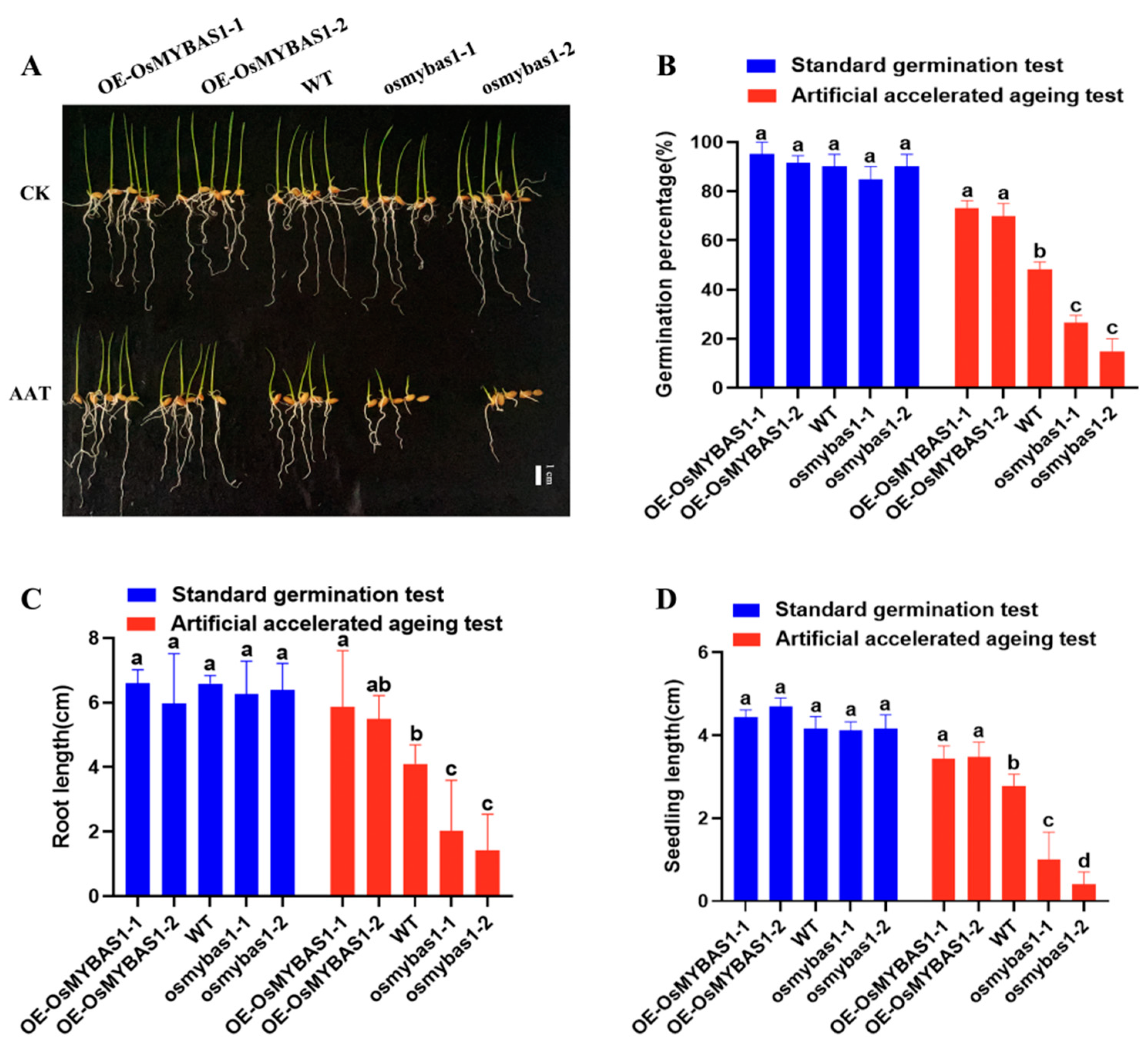
3.1. Effects of OsMYBAS1 on Rice Seed Germination
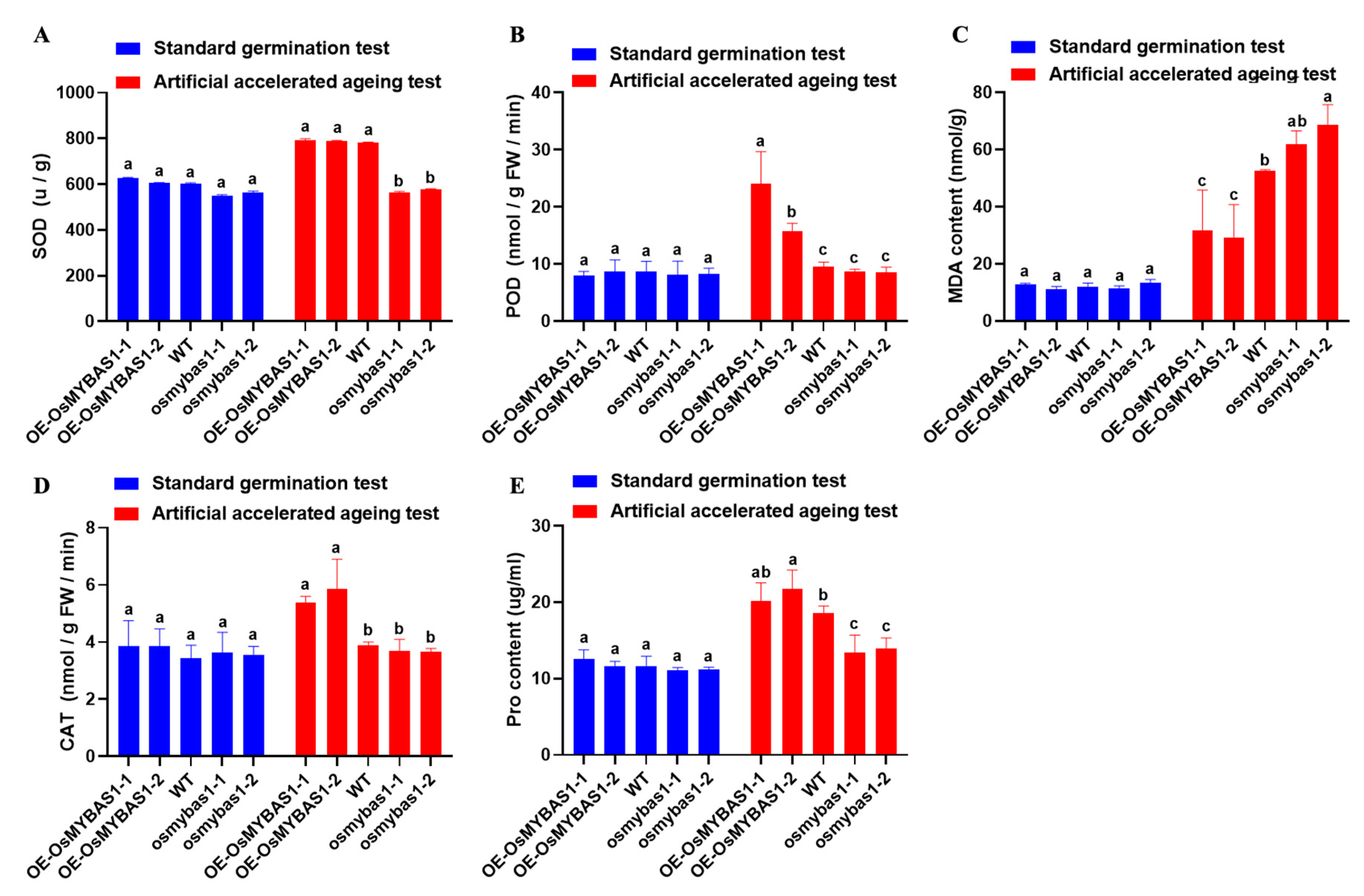
3.2. Effects of OsMYBAS1 on the Antioxidant Capacity of Rice Seed
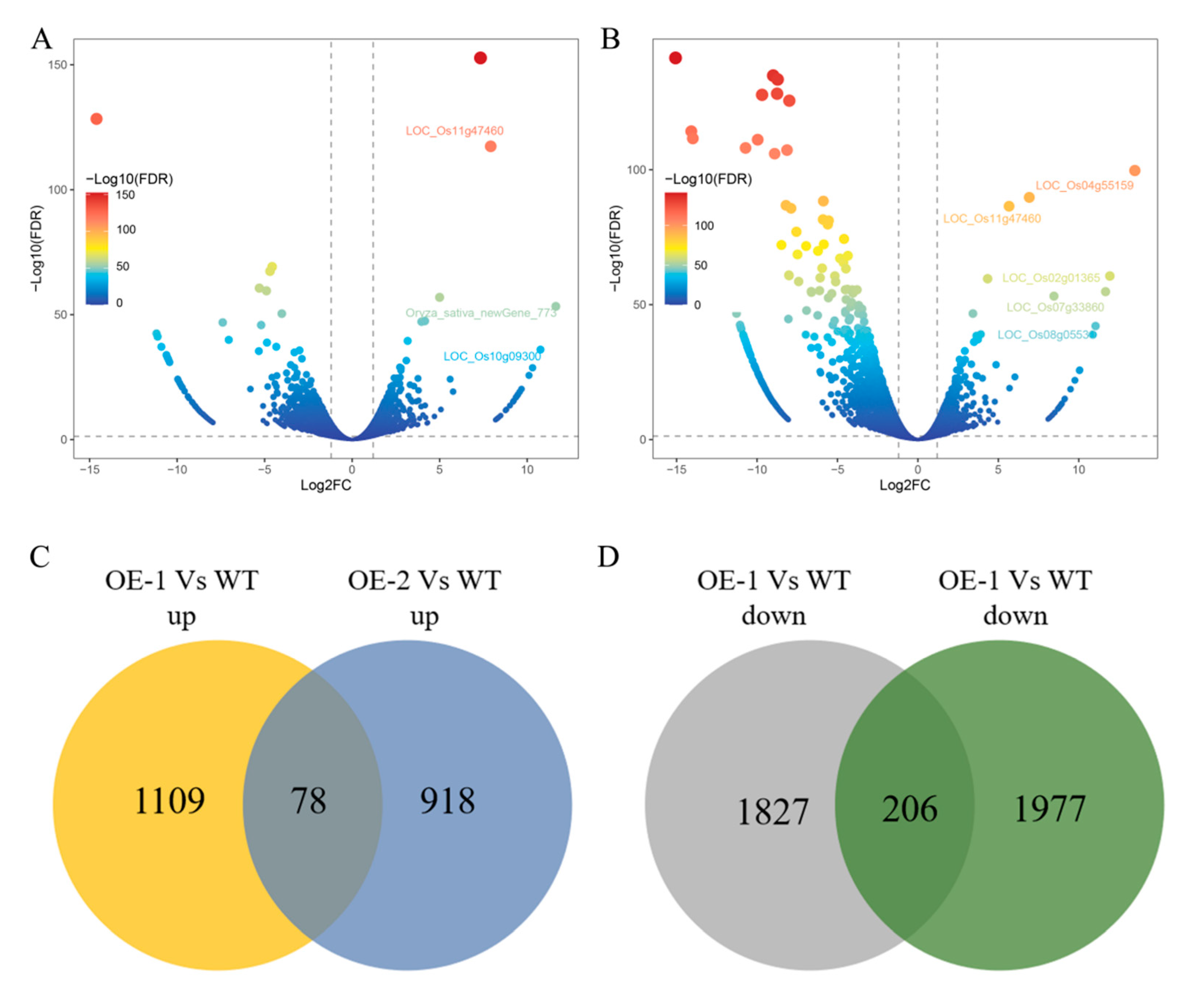
3.3. Artificial Accelerated Aging Induced Transcriptome Changes in Rice
3.4. GO and KEGG Enrichment Analysis of DEGs
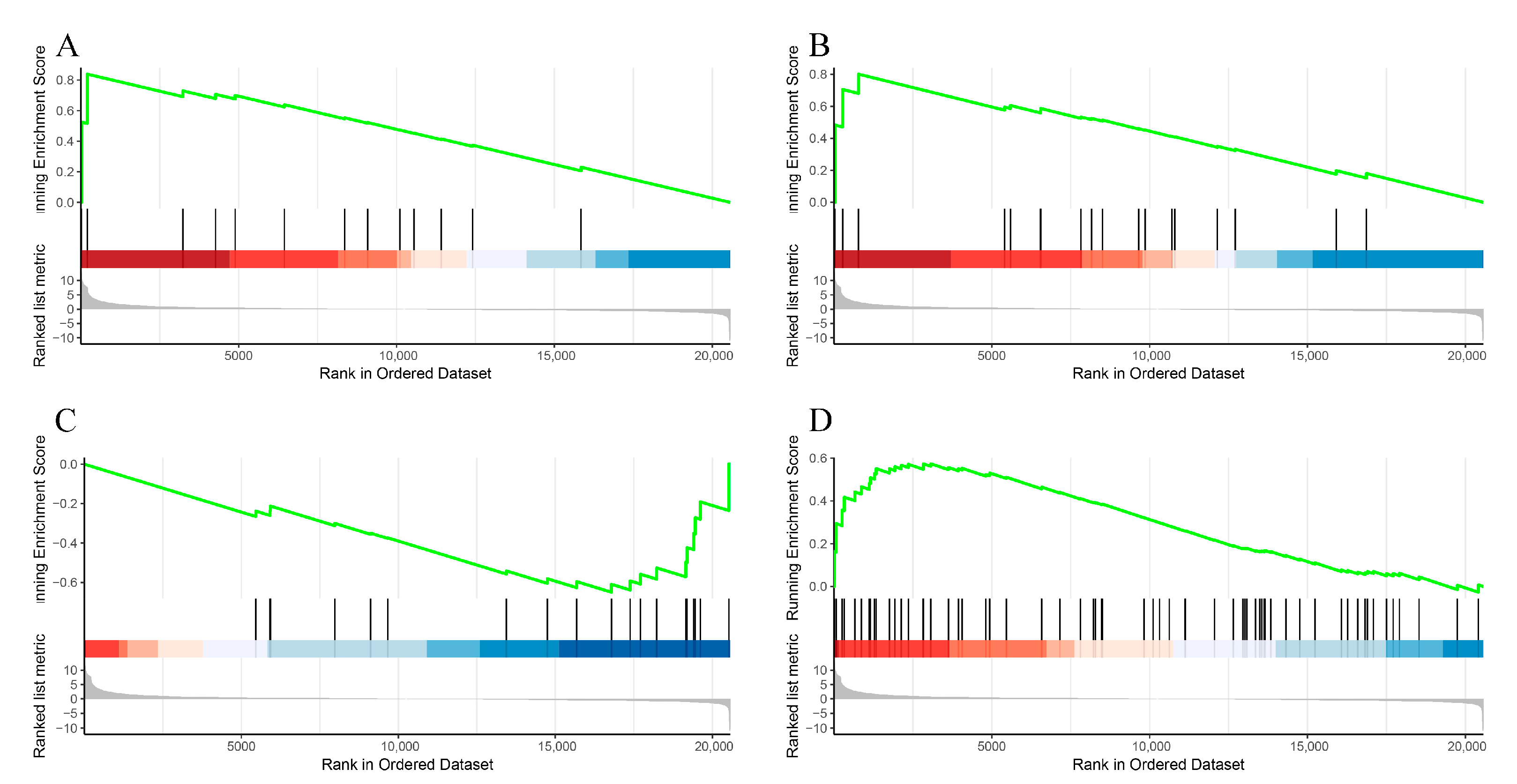
3.5. GSEA Analysis
3.6. Identification and Validation of Antioxidation-Related Genes and Downstream Transcription Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El-Maarouf-Bouteau, H.; Mazuy, C.; Corbineau, F.; Bailly, C. DNA alteration and programmed cell death during ageing of sunflower seed. J. Exp. Bot. 2011, 62, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.K.; Xin, X.; Song, C.; Chen, X.L.; Zhang, J.M.; Wu, S.H.; Li, R.F.; Liu, X.; Lu, X.X. Activity levels and expression of antioxidant enzymes in the ascorbate–Glutathione cycle in artificially aged rice seed. Plant Physiol. Biochem. 2014, 80, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jeevan, K.; Rajendra, P.S.; Rintu, B.; Chakradhar, T. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Ewelina, R.; Malecka, A.; Ciereszko, I.; Staszak, A.M. Mitochondria Are Important Determinants of the Aging of Seeds. Int. J. Mol. Sci. 2019, 20, 1568. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Fan, K.; Xia, L.F.; Ding, X.L.; Tian, L.; Zhang, W.Q.; He, H.Z.; Yu, S.B. Genetic Dissection of Seed Storability and Validation of Candidate Gene Associated with Antioxidant Capability in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 4442. [Google Scholar] [CrossRef]
- Huang, J.X.; Cai, M.H.; Long, Q.Z.; Liu, L.L.; Lin, Q.Y.; Jiang, L.; Chen, S.H.; Wan, J.M. OsLOX2, a rice type I lipoxygenase, confers opposite effects on seed germination and longevity. Transgenic Res. 2014, 23, 643–655. [Google Scholar] [CrossRef]
- Xu, H.B.; Wei, Y.D.; Zhu, Y.S.; Lian, L.; Xie, H.G.; Cai, Q.H.; Chen, Q.S.; Lin, Z.P.; Wang, Z.H.; Xie, H.A.; et al. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotechnol. J. 2015, 13, 526–539. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Shin, J.H.; Kim, S.R.; An, G. Rice aldehyde dehydrogenase 7 is needed for seed maturation and viability. Plant Physiol. 2009, 149, 905–915. [Google Scholar] [CrossRef]
- Ribeiroa, C.W.; Korbesb, A.P.; Garighanb, J.A.; Jardim-Messederb, D.; Carvalhoc, F.E.L.; Sousac, R.H.V.; Caverzanb, A.; Teixeirab, F.K.; Silveirac, J.A.G.; Margis-Pinheiroa, M. Rice peroxisomal ascorbate peroxidase knockdown affects ROS signaling and triggers early leaf senescence. Plant Sci. 2017, 263, 55–65. [Google Scholar] [CrossRef]
- Su, C.F.; Wang, Y.C.; Hsieh, T.H.; Lu, C.A.; Tseng, T.H.; Yu, S.M. A novel MYBS3-dependent pathway confers cold tolerance in rice. Plant Physiol. 2010, 153, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Fávero Peixoto-Junior, R.; Mara de Andrade, L.; dos Santos Brito, M.; Macedo Nobile, P.; Palma Boer Martins, A.; Domingues Carlin, S.; Vasconcelos Ribeiro, R.; Helena de Souza Goldman, M.; Felipe Nebó Carlos de Oliveira, J.; Vargas de Oliveira Figueira, A.; et al. Overexpression of ScMYBAS1 alternative splicing transcripts differentially impacts biomass accumulation and drought tolerance in rice transgenic plants. PLoS ONE 2018, 13, e0207534. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Dai, X.Y.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- El-Kereamy, A.; Bi, Y.M.; Ranathunge, K.; Beatty, P.H.; Good, A.G.; Rothstein, S.J. The rice R2R3-MYB transcription factor OsMYB55 is involved in the tolerance to high temperature and modulates amino acid metabolism. PLoS ONE 2012, 7, e52030. [Google Scholar] [CrossRef]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.L.; Sederoff, R.R.; Zhao, X. MYB-mediated regulation of anthocyanin biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Yang, X.; Ju, Q.; Li, W.; Lü, S.; Tran, L.P.; Xu, J. The R2R3-MYB transcription factor AtMYB49 modulates salt tolerance in Arabidopsis by modulating the cuticle formation and antioxidant defence. Plant Cell Environ. 2020, 43, 1925–1943. [Google Scholar] [CrossRef]
- Beathard, C.; Mooney, S.; Saharin, R.A.; Goyer, A.; Hellmann, H. Characterization of Arabidopsis thaliana R2R3 S23 MYB Transcription Factors as Novel Targets of the Ubiquitin Proteasome-Pathway and Regulators of Salt Stress and Abscisic Acid Response. Front. Plant Sci. 2021, 12, 629208. [Google Scholar] [CrossRef]
- Chen, Y.S.; Ho, T.D.; Liu, L.H.; Lee, D.H.; Lee, C.H.; Chen, Y.R.; Lin, S.Y.; Lu, C.A.; Yu, S.M. Sugar starvation-regulated MYBS2 and 14-3-3 protein interactions enhance plant growth, stress tolerance, and grain weight in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef]
- Mehrotra, S.; Verma, S.; Kumar, S.; Kumari, S.; Mishra, B.N. Transcriptional regulation and signalling of cold stress response in plants: An overview of current understanding. Environ. Exp. Bot. 2020, 180, 104243. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.F.; Liu, X.Y.; Du, H.; Dai, M.Q.; Zhou, D.X.; Yang, W.J.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef]
- Wang, X.M.; Wu, R.; Shen, T.S.; Li, Z.N.; Li, C.Y.; Wu, B.K.; Jiang, H.Y.; Zhao, G.W. An R2R3-MYB Transcription Factor OsMYBAS1 Promotes Seed Germination under Different Sowing Depths in Transgenic Rice. Plants 2022, 11, 139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, J.; Qin, G.C.; Cui, H.W.; Wang, Q.T. Salicylic acid analogues with biological activity may induce chilling tolerance of maize (Zea mays) seeds. Botany 2012, 90, 845–855. [Google Scholar] [CrossRef]
- Pinhero, R.G.; Rao, M.V.; Paliyath, G.; Murr, D.P.; Fletcher, R.A. Changes in Activities of Antioxidant Enzymes and Their Relationship to Genetic and Paclobutrazol-Induced Chilling Tolerance of Maize Seedlings. Plant Physiol. 1997, 114, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, J.G.; Gao, Y.; Wang, C.; Guo, G.Y.; Luo, Y.; Huang, Y.T.; Hu, W.M.; Sheteiwy, M.S.; Guan, Y.J.; et al. The Synergistic Priming Effect of Exogenous Salicylic Acid and H2O2 on Chilling Tolerance Enhancement during Maize (Zea mays L.) Seed Germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef]
- Zhang, X.H.; Shen, L.; Li, F.J.; Zhang, Y.X.; Meng, D.; Sheng, J. Up-regulating arginase contributes to amelioration of chilling stress and the antioxidant system in cherry tomato fruits. J. Sci. Food Agric. 2010, 90, 2195–2202. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.F.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Yu, G.C.; Wang, L.G.; Han, Y.Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Pommerrenig, B.; Ludewig, F.; Cvetkovic, J.; Trentmann, O.; Klemens, P.A.W.; Neuhaus, H.E. In Concert: Orchestrated Changes in Carbohydrate Homeostasis Are Critical for Plant Abiotic Stress Tolerance. Plant Cell physiol. 2018, 59, 1290–1299. [Google Scholar] [CrossRef]
- Gupta, A.K.; Kaur, N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 2005, 30, 761–776. [Google Scholar] [CrossRef]
- Kelly, A.A.; Feussner, I. Oil is on the agenda: Lipid turnover in higher plants. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1253–1268. [Google Scholar] [CrossRef] [PubMed]
- Alché, J.D.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef] [PubMed]
- Kiran, U.; Abdin, M.Z. Computational predictions of common transcription factor binding sites on the genes of proline metabolism in plants. Bioinformation 2012, 8, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Xing, G.L.; Li, J.Y.; Li, W.L.; Lam, S.M.; Yuan, H.L.; Shui, G.H.; Yang, J.S. AP2/ERF and R2R3-MYB family transcription factors: Potential associations between temperature stress and lipid metabolism in Auxenochlorella protothecoides. Biotechnol. Biofuels 2021, 14, 22. [Google Scholar] [CrossRef]
- Zheng, Y.P.; Deng, X.X.; Qu, A.L.; Zhang, M.M.; Tao, Y.; Yang, L.Y.; Liu, Y.D.; Xu, J.; Zhang, S.Q. Regulation of pollen lipid body biogenesis by MAP kinases and downstream WRKY transcription factors in Arabidopsis. PLoS Genet. 2018, 14, e1007880. [Google Scholar] [CrossRef]
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar]
- Ma, D.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Dai, X.Y.; Wang, Y.Y.; Yang, A.; Zhang, W.H. OsMYB2P-1, an R2R3 MYB transcription factor, is involved in the regulation of phosphate-starvation responses and root architecture in rice. Plant Physiol. 2012, 159, 169–183. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Hsu, P.K.; Dubeaux, G.; Takahashi, Y.; Schroeder, J.I. Signaling mechanisms in abscisic acid-mediated stomatal closure. Plant J. 2021, 105, 307–321. [Google Scholar] [CrossRef]
- Wang, T.J.; Huang, S.Z.; Zhang, A.; Guo, P.; Liu, Y.T.; Xu, C.M.; Cong, W.X.; Liu, B.; Zheng, Y.X. JMJ17-WRKY40 and HY5-ABI5 modules regulate the expression of ABA-responsive genes in Arabidopsis. New Phytol. 2021, 230, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Viana, V.E.; Maia, L.; Busanello, C.; Pegoraro, C.; Oliveira, A. When rice gets the chills: Comparative transcriptome profiling at germination shows WRKY transcription factor responses. Plant Biol. 2021, 23, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Han, M.H.; Kim, C.Y.; Lee, J.; Lee, S.K.; Jeon, J.S. OsWRKY42 represses OsMT1d and induces reactive oxygen species and leaf senescence in rice. Mol. Cells 2014, 37, 532–539. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, R.; Ding, Y.; Li, C.; Wu, B.; Huang, Z.; Li, Z.; Wang, X.; Zhao, G. An R2R3-Type Transcription Factor OsMYBAS1 Regulates Seed Germination under Artificial Accelerated Aging in Transgenic Rice (Oryza sativa L.). Agronomy 2022, 12, 1955. https://doi.org/10.3390/agronomy12081955
Wu R, Ding Y, Li C, Wu B, Huang Z, Li Z, Wang X, Zhao G. An R2R3-Type Transcription Factor OsMYBAS1 Regulates Seed Germination under Artificial Accelerated Aging in Transgenic Rice (Oryza sativa L.). Agronomy. 2022; 12(8):1955. https://doi.org/10.3390/agronomy12081955
Chicago/Turabian StyleWu, Rong, Yunqian Ding, Chenyong Li, Bangkui Wu, Zhongji Huang, Zhenan Li, Xiaomin Wang, and Guangwu Zhao. 2022. "An R2R3-Type Transcription Factor OsMYBAS1 Regulates Seed Germination under Artificial Accelerated Aging in Transgenic Rice (Oryza sativa L.)" Agronomy 12, no. 8: 1955. https://doi.org/10.3390/agronomy12081955